
Impacts of Climate Change on Blue Carbon Stocks and Fluxes in Mangrove Forests


Abstract
:1. Introduction
2. Mangrove Blue Carbon: An Update
2.1. Carbon Stocks
2.2. Carbon Burial Rates
2.3. Carbon Mass Balance Dynamics
3. Climate Change Impacts
- Mangrove primary production and canopy respiration.
- Soil CORG accumulation and burial.
- Forest–coastal water exchange of dissolved carbon.
- Carbon processes within the soil horizon.
3.1. Sea-Level Rise (SLR)
3.2. Rising Temperatures, Increased Storms, Extreme Weather Events, and Precipitation Changes
3.3. Rising Atmospheric CO2
4. Predictions
Funding
Data Availability Statement
Conflicts of Interest
References
- Alongi, D.M. Mangroves. In Encyclopedia of Estuaries; Kennish, M., Ed.; Springer: Berlin, Germany, 2016; pp. 393–404. [Google Scholar]
- Feller, I.C.; Lovelock, C.E.; Berger, U.; McKee, K.L.; Joye, S.B.; Ball, M.C. Biocomplexity in mangrove ecosystems. Annu. Rev. Mar. Sci. 2010, 2, 395–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alongi, D.M. Blue Carbon: Carbon Sequestration for Climate Change Mitigation; Springer Nature: Cham, Switzerland, 2018. [Google Scholar]
- Friess, D.; Krauss, K.W.; Taillardat, P.; Adame, M.F.; Yando, E.S.; Cameron, C.; Sasmito, S.D.; Sillanpää, M. Mangrove blue carbon in the face of climate change, and restoration. Annu. Plant Rev. 2020, 3, 427–456. [Google Scholar]
- Jennerjahn, T.C.; Gilman, E.; Krauss, K.W.; Lacerda, L.D.; Nordhaus, I.; Wolanski, E. Mangrove ecosystems under climate change. In Mangrove Ecosystems: A Global Biogeographic Perspective; Rivera-Monroy, V.H., Lee, S.Y., Kristensen, E., Twilley, R.R., Eds.; Springer Nature: Cham, Switzerland, 2017; pp. 211–244. [Google Scholar]
- Alongi, D.M. The Energetics of Mangrove Forests; Springer: Dordrecht, The Netherlands, 2009. [Google Scholar]
- Alongi, D.M.; Mukhopadhyay, S.K. Contribution of mangroves to coastal carbon cycling in low latitude seas. Agric. For. Meteorol. 2015, 213, 266–272. [Google Scholar] [CrossRef]
- Alongi, D.M. Global significance of mangrove blue carbon in climate change mitigation. Science 2020, 2, 67. [Google Scholar] [CrossRef]
- Adame, M.F.; Santini, N.S.; Torres-Talamante, O.; Rogers, K. Mangrove sinkholes (cenotes) of the Yucatan Peninsula, a global hotspot of carbon sequestration. Biol. Lett. 2021, 17, 20210037. [Google Scholar] [CrossRef] [PubMed]
- Kida, M.; Watanabe, I.; Kinjo, K.; Kondo, M.; Yoshitake, S.; Tomotsune, M.; Iimura, Y.; Umnouysin, S.; Suchewaboripont, V.; Poungparn, S.; et al. Organic carbon stock and composition in 3.5-m core mangrove soils (Trat, Thailand). Sci. Tot. Environ. 2021, 801, 149682. [Google Scholar] [CrossRef] [PubMed]
- Trettin, C.C.; Dai, Z.; Tang, W.; Lagomasino, D.; Thomas, N.; Lee, S.K.; Simard, M.; Ebanega, M.O.; Stoval, A.; Fatoyinbo, T.E. Mangrove carbon stocks in Pongara National Park, Gabon. Estuar. Coast. Shelf Sci. 2021, 259, 107432. [Google Scholar] [CrossRef]
- Stringer, C.E.; Trettin, C.C.; Zarnoch, S.J.; Tang, W. Carbon stocks of mangroves within the Zambezi River Delta, Mozambique. For. Ecol. Manage. 2015, 354, 139–148. [Google Scholar] [CrossRef]
- Johnson, J.L.; Raw, J.L.; Adams, J.B. First report on carbon storage in a warm-temperate mangrove forest in South Africa. Estuar. Coast. Shelf Sci. 2020, 235, 106566. [Google Scholar] [CrossRef]
- Bai, J.; Meng, Y.; Gou, R.; Lyu, J.; Dai, Z.; Diao, X.; Zhang, H.; Luo, Y.; Zhu, X.; Lin, G. Mangrove diversity enhances plant biomass production and carbon storage in Hainan Island, China. Funct. Ecol. 2021, 35, 774–786. [Google Scholar] [CrossRef]
- Ragavan, P.; Kumar, S.; Kathiresan, k.; Mohan, P.M.; Jayaraj, R.S.C.; Ravichandaran, K.; Rana, T.S. Biomass and vegetation carbon stock in mangrove forests of the Andaman Islands, India. Hydrobiologia 2021, 848, 4673–4693. [Google Scholar] [CrossRef]
- Chatting, M.; LeVay, L.; Walton, M.; Skov, M.W.; Kennedy, H.; Wilson, S.; Al-Maslamani, I. Mangrove carbon stocks and biomass partitioning in an extreme environment. Estuar. Coast. Shelf Sci. 2020, 244, 106940. [Google Scholar] [CrossRef]
- Cooray, P.L.I.G.M.; Kodikara, K.A.S.; Kumara, M.P.; Jayasinghe, U.I.; Madarasinghe, S.K.; Dahdouh-Guebas, F.; Gorman, D.; Huxman, M.; Jayatissa, L.P. Climate and intertidal zonation drive variability in the carbon stocks of Sri Lankan mangrove forests. Geoderma 2021, 389, 114929. [Google Scholar]
- Banderjee, K.; Mitra, A.; Villasante, S. Carbon cycling in mangrove ecosystem of Western Bay of Bengal (India). Sustainability 2021, 13, 6740. [Google Scholar] [CrossRef]
- Merecí-Guamán, J.; Casanoves, F.; Delgado-Rodríguez, D.; Ochoa, P.; Cifuentes-Jara, M. Impact of shrimp ponds on mangrove blue carbon stocks in Ecuador. Forests 2021, 12, 816. [Google Scholar] [CrossRef]
- Barik, J.; Sanyal, P.; Ghosh, T.; Mukhopadhyay, S.K. Carbon stock and storage pattern in the Sundarbans mangrove forest, NE coast of India. Trop. Ecol. 2021, 62, 95–106. [Google Scholar] [CrossRef]
- Zakaria, R.M.; Chen, G.; Chew, L.L.; Sofawi, A.B.; Moh, H.H.; Chen, S.; Teoh, H.W.; Adibah, S.Y.S.N. Carbon stock of disturbed and undisturbed mangrove ecosystems in Klang Straits, Malaysia. J. Sea Res. 2021, 176, 1022113. [Google Scholar] [CrossRef]
- Pricillia, C.C.; Patria, M.P.; Herdiansyah, H. Environmental conditions to support blue carbon storage in mangrove forest: A case study in the mangrove forest, Nusa Lembongan, Bali, Indonesia. Biodiversitas 2021, 22, 3304–3314. [Google Scholar] [CrossRef]
- Wang, G.; Yu, C.; Singh, M.; Guan, D.; Xiong, Y.; Zheng, R.; Xiao, R. Community structure and ecosystem carbon stock dynamics along a chronosequence of mangrove plantations in China. Plant Soil 2021, 464, 605–620. [Google Scholar] [CrossRef]
- Hatje, V.; Masqué, P.; Patire, V.F.; Dórea, A.; Barros, F. Blue carbon stocks, accumulation rates, and associated spatial variability in Brazilian mangroves. Limnol. Oceanogr. 2020, 66, 321–334. [Google Scholar] [CrossRef]
- Kusumaningtyas, M.A.; Kepel, T.L.; Solihuddin, T.; Lubis, A.A.; Putra, A.D.P. Carbon sequestration potential in the rehabilitated mangroves in Indonesia. Ecol. Res. 2021, 37, 80–91. [Google Scholar] [CrossRef]
- Yu, C.; Guan, D.; Gang, W.; Lou, D.; Wei, L.; Zhou, Y.; Feng, J. Development of ecosystem carbon stock with the progression of a natural mangrove forest in Yingluo Bay, China. Plant Soil 2021, 460, 391–401. [Google Scholar] [CrossRef]
- Rovai, A.S.; Coelho, C., Jr.; de Almeida, R.; Cunha-Lignon, M.; Menghini, R.P.; Twilley, R.R.; Cintrón-Molero, G.; Schaeffer-Novelli, Y. Ecosystem-level carbon stocks and sequestration rates in mangroves in the Cananéia-Iguape estuarine system, southeastern Brazil. For. Ecol Manage. 2021, 479, 118553. [Google Scholar] [CrossRef]
- Murdiyarso, D.; Sasmito, S.D.; Sillanpää, M.; MacKenzie, R.; Gaveau, D. Mangrove selective logging sustains biomass carbon recovery, soil carbon, and sediment. Sci. Rep. 2021, 11, 12325. [Google Scholar] [CrossRef]
- Hamilton, S.E.; Casey, D. Creation of high spatio-temporal resolution global database of continuous mangrove forest cover for the 21st century (CGMFC-21). Glob. Ecol. Biogeogr. 2016, 25, 729–738. [Google Scholar] [CrossRef]
- Giri, C.; Ochieng, E.; Tiezen, L.L.; Zhu, Z.; Singh, A.; Loveland, T.; Masek, J.; Duke, N.C. Status and distribution of mangrove forests of the world using earth observation satellite data. Glob. Ecol. Biogeogr. 2011, 20, 154–159. [Google Scholar] [CrossRef]
- Sanders, C.J.; Maher, D.T.; Tait, D.R.; Williams, D.; Holloway, C.; Sippo, J.Z.; Santos, I.R. Are global mangrove carbon stocks driven by rainfall? J. Geophys. Res. Biogeosci. 2016, 121, 2600–2609. [Google Scholar] [CrossRef]
- Collins, D.S.; Nguyen, V.L.; Ta, T.K.O.; Mao, L.; Ishii, Y.; Ktagawa, H.; Nakashima, R.; Vo, T.H.Q.; Tamura, T. Sedimentary evolution of a delta-margin mangrove in Can Gio, northeastern Mekong River delta, Vietnam. Mar. Geol. 2021, 433, 106417. [Google Scholar] [CrossRef]
- Hanggara, B.B.; Murdiyarso, D.; Ginting, Y.R.S.; Widha, Y.L.; Panjaitan, G.Y.; Lubis, A.A. Effects of diverse mangrove management practices on forest structure, carbon dynamics and sedimentation in North Sumatra, Indonesia. Estuar. Coast. Shelf Sci. 2021, 259, 107467. [Google Scholar] [CrossRef]
- Wigand, C.; Eagle, M.; Branoff, B.L.; Balogh, S.; Miller, K.M.; Martin, R.M.; Hanson, A.; Oczkowski, A.J.; Huertas, E.; Loffredo, J.; et al. Recent carbon storage and burial exceed historical rates in the San Juan Bay estuary peri-urban mangrove forests (Puerto Rico, United States). Front. For. Glob. Change 2021, 4, 676691. [Google Scholar] [CrossRef]
- Chen, L.; Lin, Q.; Krauss, K.W.; Zhang, Y.; Cormier, N.; Yang, Q. Forest thinning in the seaward fringe speeds up surface elevation increment and carbon accumulation in managed mangrove forests. J. Appl. Ecol. 2021, 58, 1899–1909. [Google Scholar] [CrossRef]
- Liu, T.; Liu, S.; Wu, B.; Xu, H.; Zhang, H. Increase of organic carbon burial response to mangrove expansion in the Nanliu River estuary, South China Sea. Prog. Earth Planet. Sci. 2020, 7, 71. [Google Scholar] [CrossRef]
- Setyadi, G.; Sugianto, D.N.; Wijayanti, D.P.; Pribadi, R.; Supardy, E. Sediment accretion and total organic carbon accumulation among different mangrove vegetation communities in the Kamora Estuary of Mimika Regency, Papua, Indonesia. J. Ecol. Eng. 2021, 22, 142–156. [Google Scholar] [CrossRef]
- Zhang, Y.; Meng, X.; Xia, P.; Zhang, J.; Liu, D.; Li, Z.; Wang, W. Spatiotemporal variations in the organic carbon accumulation rate in mangrove sediments from the Yingluo Bay, China, since 1900. Acta Oceanol. Sinica 2021, 40, 65–77. [Google Scholar] [CrossRef]
- Passos, T.; Penny, D.; Sanders, C.; De França, E.; Oliveira, T.; Santos, L.; Barcellos, R. Mangrove carbon and nutrient accumulation shifts driven by rapid development in a tropical estuarine system, northeast Brazil. Mar. Pollut. Bull 2021, 166, 112219. [Google Scholar] [CrossRef] [PubMed]
- Rani, V.; Nandan, S.B.; Schwing, P.T. Carbon source characterization and historical carbon burial in three mangrove ecosystems on the southwest coast of India. Catena 2021, 197, 104980. [Google Scholar] [CrossRef]
- MacKenzie, R.; Sharma, S.; Rovai, A.R. Environmental drivers of blue carbon burial and soil carbon stocks in mangrove forests. In Dynamic Sedimentary Environments of Mangrove Coasts; Sidik, F., Friess, D.A., Eds.; Elsevier: Amsterdam, The Netherlands, 2021; pp. 275–294. [Google Scholar]
- Nardin, W.; Vona, I.; Fagherazzi, S. Sediment deposition affects mangrove forests in the Mekong delta, Vietnam. Cont. Shelf Res. 2021, 213, 104319. [Google Scholar] [CrossRef]
- Alongi, D.M. Carbon cycling in the world’s mangrove ecosystems revisited: Significance of non-steady state diagenesis and subsurface linkages between the forest floor and the coastal ocean. Forests 2020, 11, 977. [Google Scholar] [CrossRef]
- Alongi, D.M. Carbon balance in salt marsh and mangrove ecosystems: A global synthesis. J. Mar. Sci. Eng. 2020, 8, 767. [Google Scholar] [CrossRef]
- Kristensen, E.; Connolly, R.M.; Otero, X.L.; Marchand, C.; Ferreira, T.O.; Rivera-Monroy, V.H. Biogeochemical cycles: Global approaches and perspectives. In Mangrove Ecosystems: A Global and Biogeographic Perspective; Rivera-Monroy, V.H., Lee, S.Y., Kristensen, E., Twilley, R.R., Eds.; Springer: Cham, Switzerland, 2017; pp. 163–209. [Google Scholar]
- Kristensen, E.; Bouillon, S.; Dittmar, T.; Marchand, C. Organic carbon dynamics in mangrove ecosystems: A review. Aquat. Bot. 2008, 89, 201–219. [Google Scholar] [CrossRef] [Green Version]
- Maher, D.T.; Santos, I.R.; Schulz, K.G.; Call, M.; Jacobsen, G.E.; Sanders, C.J. Blue carbon oxidation revealed by radiogenic and stable isotopes in a mangrove system. Geophys. Res. Lett. 2017, 44, 4889–4896. [Google Scholar] [CrossRef]
- Ward, R.D.; de Lacerda, L.D. Responses of mangrove ecosystems to sea level change. In Dynamic Sedimentary Environments of Mangrove Coasts; Sidik, F., Friess, D.A., Eds.; Elsevier: Amsterdam, The Netherlands, 2021; pp. 235–253. [Google Scholar]
- IPCC. Climate change 2021: The physical science basis. In Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, S.L., Connors, S.L., Péan, C., Berger, S., Caud, M., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge: Cambridge, UK, 2021; pp. 1–676. [Google Scholar]
- Woodroffe, C.D.; Rogers, K.; McKee, K.L.; Lovelock, C.E.; Mendelssohn, I.A.; Saintilan, N. Mangrove sedimentation and response to relative sea-level rise. Annu. Rev. Mar Sci. 2016, 8, 243–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friess, D.A.; McKee, K.L. The history of surface-elevation paradigms in mangrove biogeomorphology. In Dynamic Sedimentary Environments of Mangrove Coasts; Sidik, F., Friess, D.A., Eds.; Elsevier: Amsterdam, The Netherlands, 2021; pp. 179–198. [Google Scholar]
- Alongi, D.M. Responses of mangrove ecosystems to climate change in the Anthropocene. In Mangroves: Ecology, Biodiversity, and Management; Rastogi, R.P., Phulwara, M., Gupta, D.K., Eds.; Springer Nature: Singapore, 2021; pp. 201–224. [Google Scholar]
- Saintilan, N.; Khan, N.S.; Ashe, E.; Kelleway, J.J.; Rogers, K.; Woodroffe, C.D.; Horton, B.P. Thresholds of mangrove survival under rapid sea-level rise. Science 2020, 368, 1118–1121. [Google Scholar] [PubMed]
- McKee, K.L.; Cahoon, D.R.; Feller, I.C. Caribbean mangroves adjust to rising sea level through biotic controls on change in soil elevation. Glob. Ecol. Biogeogr. 2007, 16, 545–556. [Google Scholar] [CrossRef]
- Kelleway, J.J.; Saintilan, N.; Macreadie, P.I.; Skilbeck, C.G.; Zawadzki, A.; Ralph, P.J. Seventy years of continuous encroachment substantially increase ‘blue carbon’ capacity as mangroves replace salt marshes. Glob. Change Biol. 2016, 22, 1097–1109. [Google Scholar]
- Rogers, K.; Kelleway, J.J.; Saintilan, N.; Megonigal, J.P.; Adams, J.B.; Holmquist, J.R.; Lu, M.; Schile-Beers, L.; Zawadzki, A.; Mazumder, D.; et al. Wetland carbon storage controlled by millennial-scale variation in relative sea-level rise. Nature 2019, 567, 91–95. [Google Scholar] [CrossRef]
- Buffington, K.J.; MacKenzie, R.A.; Carr, J.A.; Apwong, M.; Krauss, K.W.; Thorne, K.M. Mangrove species’ response to sea-level rise across Pohnpei, Federated States of Micronesia. In U.S. Geological Survey Open-File Report 2021–1002; U.S. Geological Survey: Reston, VA, USA, 2021; pp. 1–44. [Google Scholar] [CrossRef]
- Lovelock, C.E.; Reef, R. Variable impacts of climate change on blue carbon. One Earth 2020, 3, 195–211. [Google Scholar] [CrossRef]
- Zhang, Y.; Meng, X.; Xia, P.; Li, Z. Response of mangrove development to air temperature variation over the past 3000 years in Quizhou Bay, tropical China. Front. Earth Sci. 2021, 9, 678189. [Google Scholar] [CrossRef]
- Cavanaugh, K.C.; Parker, J.D.; Cook-Patton, S.C.; Feller, I.C.; Park Williams, A.; Kellner, J.R. Integrating physiological threshold experiments with climate modelling to project mangrove species’ range expansion. Glob. Change Biol. 2015, 21, 1928–1938. [Google Scholar] [CrossRef] [Green Version]
- Yando, E.S.; Osland, M.J.; Willis, J.M.; Day, R.H.; Krauss, K.W.; Hester, M.W. Salt marsh-mangrove ecotones: Using structural gradients to investigate the effects of woody plant encroachment on plant-soil interactions and ecosystem carbon pools. J. Ecol. 2016, 104, 1020–1031. [Google Scholar] [CrossRef] [Green Version]
- Raw, J.L.; Julie, C.L.; Adams, J.B. A comparison of soil carbon pools across a mangrove -salt marsh ecotone at the southern African warm-temperate range limit. S. Afr. J. Bot. 2019, 127, 301–307. [Google Scholar] [CrossRef]
- Simpson, L.T.; Stein, C.M.; Osborne, T.Z.; Feller, I.C. Mangroves dramatically increase carbon storage after 3 years of encroachment. Hydrobiologia 2019, 834, 13–26. [Google Scholar] [CrossRef]
- Lewis, D.B.; Brown, J.A.; Jimenez, K.L. Effects of flooding and warming on soil organic matter mineralization in Avicennia germinans mangrove forests and Juncus roemerianus salt marshes. Estuar. Coast. Shelf Sci. 2014, 139, 11–19. [Google Scholar] [CrossRef]
- Hansen, J.; Sato, M.; Ruedy, R. Perception of climate change. Proc. Nat. Acad. Sci. USA 2012, 109, E2415–E2423. [Google Scholar] [CrossRef] [Green Version]
- Lovelock, C.E.; Feller, I.C.; Reef, R.; Hickey, S.; Ball, M.C. Mangrove dieback during fluctuating sea levels. Sci. Rep. 2017, 7, 1680. [Google Scholar] [CrossRef]
- Abhik, S.; Hope, P.; Hendon, H.H.; Hutley, L.B.; Johnson, S.; Drosdowsky, W.; Brown, J.R.; Duke, N.C. Influence of the 2015–2016 El Niño on the record-breaking mangrove dieback along the northern Australia coast. Sci. Rep. 2021, 11, 20411. [Google Scholar] [CrossRef]
- Sippo, J.Z.; Sanders, C.J.; Santos, I.R.; Jeffrey, L.C.; Call, M.; Harada, Y.; Maguire, K.; Brown, D.; Conrad, S.R.; Maher, D.T. Coastal carbon cycle changes following mangrove loss. Limnol. Oceanogr. 2020, 65, 2642–2656. [Google Scholar] [CrossRef]
- Gomes, L.E.; Sanders, C.J.; Nobrega, G.N.; Vescovi, L.C.; Queiroz, H.M.; Kauffman, J.B.; Ferreira, T.O.; Bernardino, A.F. Ecosystem carbon losses following a climate-induced mangrove mortality in Brazil. J. Environ. Manage. 2021, 297, 113381. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Sun, C.; Qin, Z. Drought-induced salinity enhancement weakens mangrove greenhouse gas cycling. J. Geophys. Res. Biogeosci. 2021, 126, e2021JG006416. [Google Scholar] [CrossRef]
- Krauss, K.W.; Osland, M.J. Tropical cyclones and the organization of mangrove forests: A review. Ann. Bot. 2020, 125, 213–234. [Google Scholar] [CrossRef]
- Jones, M.C.; Wingard, G.L.; Stackhouse, B.; Keller, K.; Willard, D.; Marot, M.; Landacre, B.; Bernhardt, C.E. Rapid inundation of southern Florida coastline despite low relative sea-level rise rates during the late Holocene. Nat. Commun. 2019, 10, 3231. [Google Scholar] [CrossRef] [PubMed]
- Simard, M.; Fatoyinbo, L.; Smetanka, C.; Rivera-Monroy, V.H.; Castañeda-Moya, E.; Thomas, N.; Van der Stocken, T. Mangrove canopy height globally related to precipitation, temperature, and cyclone frequency. Nat. Geosci. 2019, 12, 40–45. [Google Scholar] [CrossRef]
- Chambers, L.G.; Steinmuller, H.E.; Breithaupt, J.L. Toward a mechanistic understanding of ‘peat collapse’ and its potential contribution to coastal wetland loss. Ecology 2019, 100, e02720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osland, M.J.; Feher, L.C.; Griffith, K.T.; Cavanaugh, K.C.; Enwright, N.M.; Day, R.H.; Stagg, C.L.; Krauss, K.W.; Howard, R.J.; Grace, J.B.; et al. Climatic controls on the global distribution, abundance, and species richness of mangrove forests. Ecol. Monogr. 2017, 87, 341–359. [Google Scholar] [CrossRef] [Green Version]
- Reef, R.; Slot, M.; Motro, U.; Motro, M.; Motro, Y.; Adame, M.F.; Garcia, M.; Aranda, J.; Lovelock, C.E.; Winter, K. The effects of CO2 and nutrient fertilization on the growth and temperature response of the mangrove Avicennia germinans. Photosyn. Res. 2016, 129, 159–170. [Google Scholar] [CrossRef] [Green Version]
- Tamimia, B.; Wan Juliana, W.A.; Nizam, M.S.; Zain, C.R.C.M. Elevated CO2 concentration and air temperature impacts on mangrove plants (Rhizophora apiculata) under controlled environment. Iraqi J. Sci. 2019, 60, 1658–1666. [Google Scholar] [CrossRef]
- Jacotot, A.; Marchand, C.; Gensous, S.; Allenbach, M. Effects of elevated atmospheric CO2 and increased tidal flooding on leaf gas-exchange parameters of two common mangrove species: Avicennia marina and Rhizophora stylosa. Photosyn. Res. 2018, 138, 249–260. [Google Scholar]
- Yin, P.; Yin, M.; Cai, Z.; Wu, G.; Lin, G.; Zhou, J. Structural instability of the rhizosphere microbiome in mangrove plant Kandelia obovata under elevated CO2. Mar. Environ. Res. 2018, 140, 422–432. [Google Scholar] [CrossRef]
- Manea, A.; Geedicke, I.; Leishman, M.R. Elevated carbon dioxide and reduced salinity enhance mangrove seedling establishment in an artificial salt marsh community. Oecologia 2020, 192, 273–280. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Region | AGBCORG | BGBCORG | SCORG | TECORG |
|---|---|---|---|---|
| Africa | 89.5 | 88.0 | 402.4 | 638.8 |
| Southeast Asia | 115.7 | 44.9 | 656.1 | 806.4 |
| South and East Asia | 94.2 | 35.9 | 445.4 | 482.8 |
| Central and North America and Caribbean | 92.5 | 41.4 | 577.4 | 730.1 |
| South America | 104.3 | 85.2 | 236.9 | 419.4 |
| Middle East | 30.6 | 41.2 | 150.9 | 224.9 |
| Australia and New Zealand | 55.1 | 79.7 | 465.3 | 563.4 |
| Pacific Islands | 213.7 | 172.9 | 599.7 | 987.4 |
| Inputs | Outputs | ||
|---|---|---|---|
| GPP (mangrove) | 35.3 | RCANOPY | 22.3 |
| GPP (microalgae) | 4.4 | RMICROALGAE | 2.3 |
| DIC production (soil to 1 m) | 18.2 | Air–soil CH4 release | 0.015 |
| Marine | ? | Soil CO2 + DIC release (air + water) | 6.13 |
| Upstream | ? | RTIDAL WATER | 3.35 |
| Groundwater | ? | Air–water CH4 release | 0.0116 |
| Net DOC export | 5.90 | ||
| Net DIC export | 14.0 | ||
| Net CH4 export | 0.0277 | ||
| Net POC export | 1.73 | ||
| Burial | 1.11 | ||
| Total | 57.9 | Total | 56.9 |
| Year Intervals | (A) CO2 Emissions Scenarios | ||||
|---|---|---|---|---|---|
| Very low | Low | Intermediate | High | Very High | |
| 2040–2060 | 4.2 (2.9–6.1) | 4.9 (3.6–6.9) | 5.9 (4.5–8.0) | 6.5 (5.1–8.7) | 7.3 (5.7–9.8) |
| 2080–2100 | 4.3 (2.5–6.6) | 5.3 (3.3–8.1) | 7.8 (5.3–11.5) | 10.4 (7.5–14.9) | 12 (8.8–17.7) |
| (B) Global surface air temperature scenarios | |||||
| 1.5 °C | 2.0 °C | 3.0 °C | 4.0 °C | 5.0 °C | |
| 2040–2060 | 4.1 (3.0–5.8) | 5.1 (3.8–7.1) | 6.0 (4.7–8.2) | 6.5 (5.1–8.6) | 7.3 (5.8–9.8) |
| 2080–2100 | 4.3 (2.6–6.5) | 5.5 (3.5–8.3) | 7.9 (5.4–11.6) | 9.9 (7.2–14.2) | 11.8 (8.6–17.0) |
| Climate Change | Blue Carbon Stocks | Carbon Fluxes |
|---|---|---|
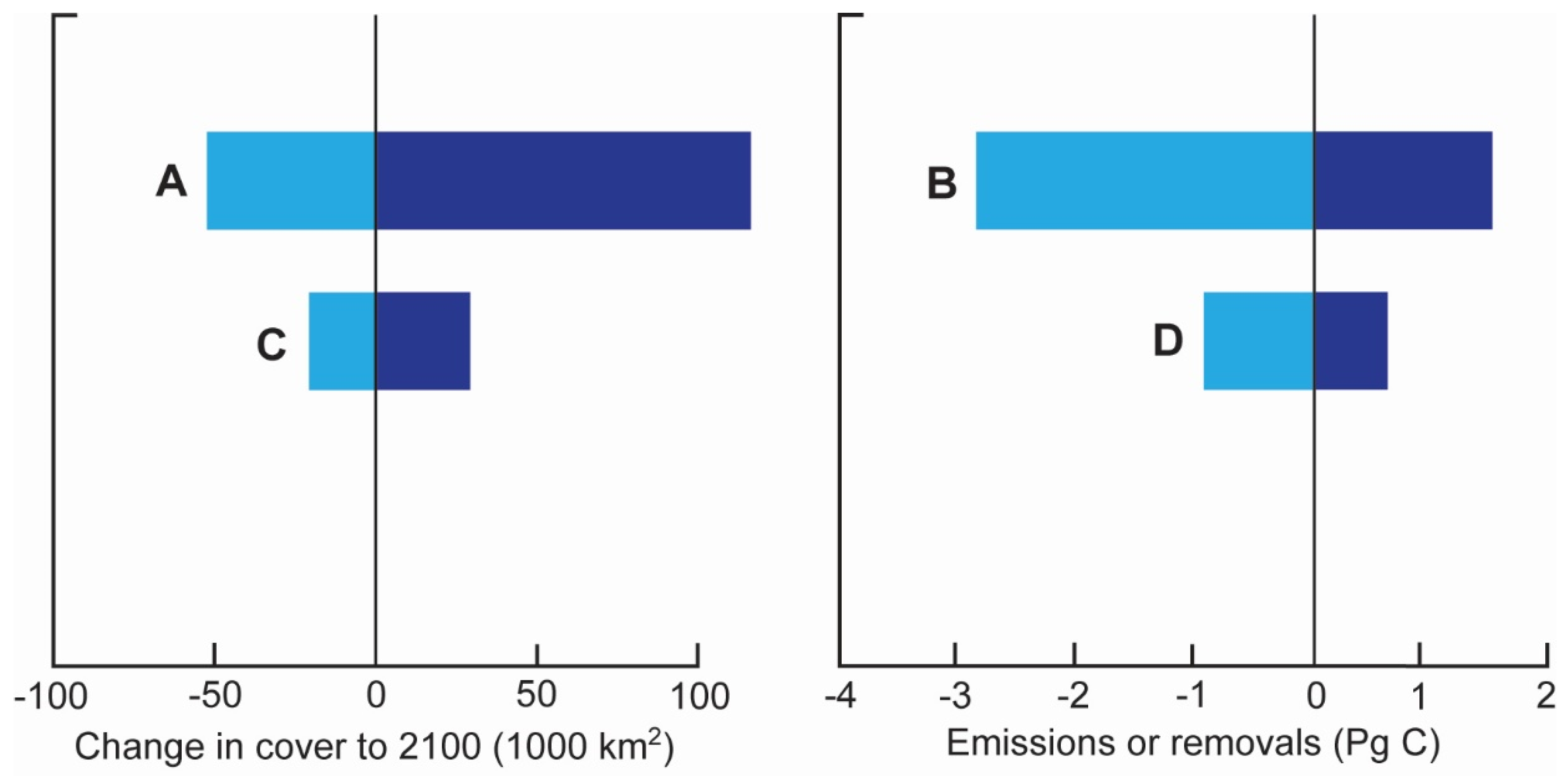
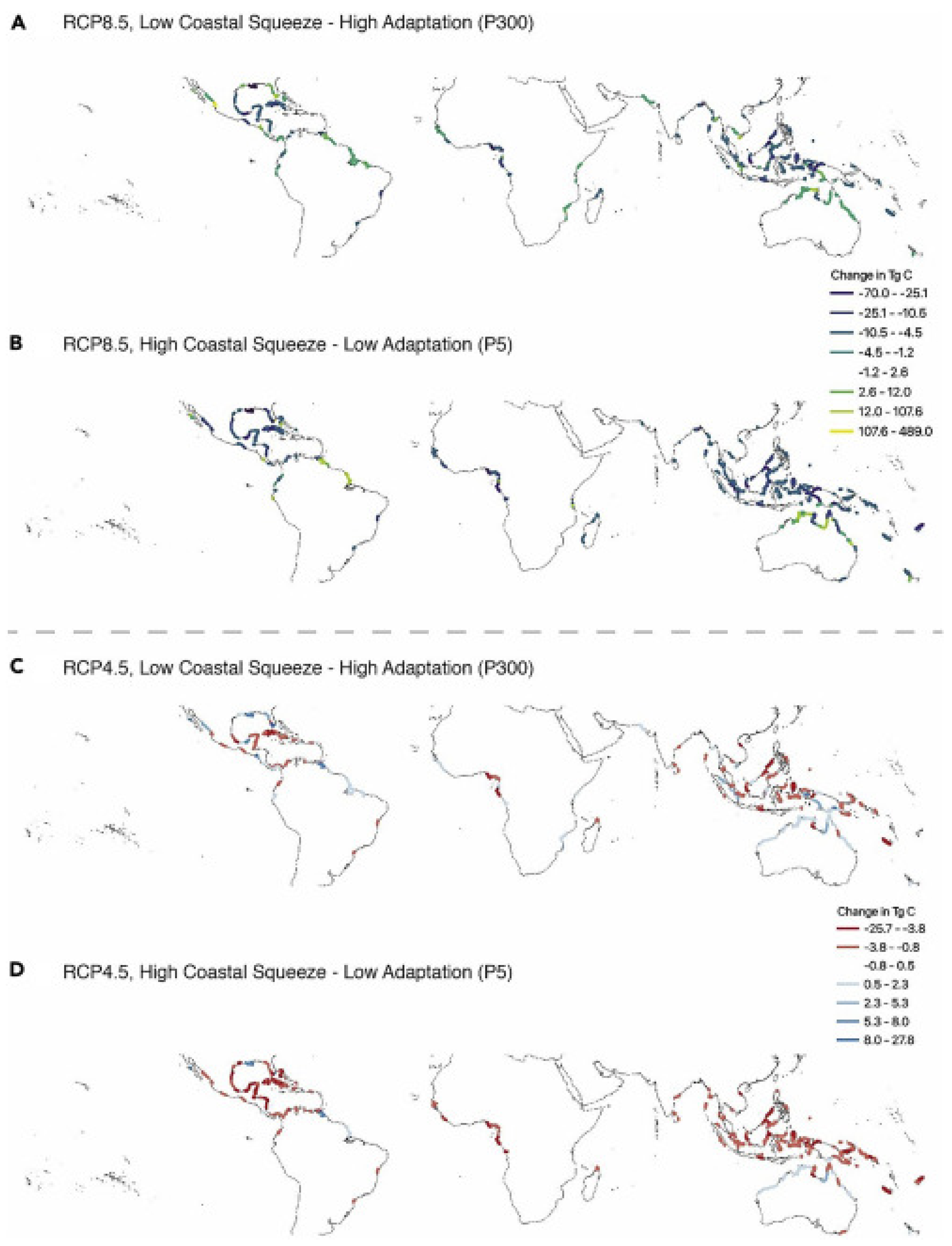
| SLR | Outcome dependent on scenario and species (flood tolerance): (1) critical SLR threshold ≈ 6 mm a−1. Survival only under very low-low CO2 emissions scenarios. (2) tidal migration or latitudinal expansion = + biomass and soil C (3) forest death = + soil C, − biomass C; + peat formation/C burial (4) net gain = +1.5 Pg C (low coastal squeeze) (5) net gains and losses = −0.3 to +0.5 Pg C (high coastal squeeze) | Outcome dependent on scenario and species (flood tolerance): (1) critical SLR threshold ≈ 6 mm a−1. Survival only under very low-low CO2 emissions scenarios. (2) tidal migration or latitudinal expansion = + CO2 and CH4 emissions (3) forest death = + in CO2 and CH4 emissions (4) under high coastal squeeze, − of 3.4 Pg C in CO2 emissions (5) under low coastal squeeze, −1.1 Pg C |
| Increasing °C | + in carbon stocks up to critical °C threshold − in carbon stocks in arid regions + in carbon stocks with continuing expansion | + in CO2 assimilation and − respiratory losses up to critical °C threshold − above °C threshold |
| Increasing Storms | − damage carbon biomass losses and soil carbon erosion + or − if storm-induced massive bedload sedimentation | − CO2 losses nutrients exhausted + or − if storm-induced massive bedload sedimentation |
| Extreme Weather Events | − massive dieback | − carbon export and CO2 and CH4 cycling |
| Precipitation Changes | + in regions of increased rainfall − in regions of decreased rainfall | + in regions of increased rainfall − in regions of decreased rainfall |
| Rising CO2 | Responses species-specific and complex due to interactive effects with °C, ‰, CO2 level, etc.: + most species, but − or 0 for some species | Responses species-specific and complex due to interactive effects with °C, ‰, CO2 level, etc.: + most species, but − or 0 for some species |
| Coastal Acidification | 0 | 0 |
| Increasing Hypoxia | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alongi, D.M. Impacts of Climate Change on Blue Carbon Stocks and Fluxes in Mangrove Forests. Forests 2022, 13, 149. https://doi.org/10.3390/f13020149
Alongi DM. Impacts of Climate Change on Blue Carbon Stocks and Fluxes in Mangrove Forests. Forests. 2022; 13(2):149. https://doi.org/10.3390/f13020149
Chicago/Turabian StyleAlongi, Daniel Michael. 2022. "Impacts of Climate Change on Blue Carbon Stocks and Fluxes in Mangrove Forests" Forests 13, no. 2: 149. https://doi.org/10.3390/f13020149
APA StyleAlongi, D. M. (2022). Impacts of Climate Change on Blue Carbon Stocks and Fluxes in Mangrove Forests. Forests, 13(2), 149. https://doi.org/10.3390/f13020149