
The Effect of Stand Density, Biodiversity, and Spatial Structure on Stand Basal Area Increment in Natural Spruce-Fir-Broadleaf Mixed Forests

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
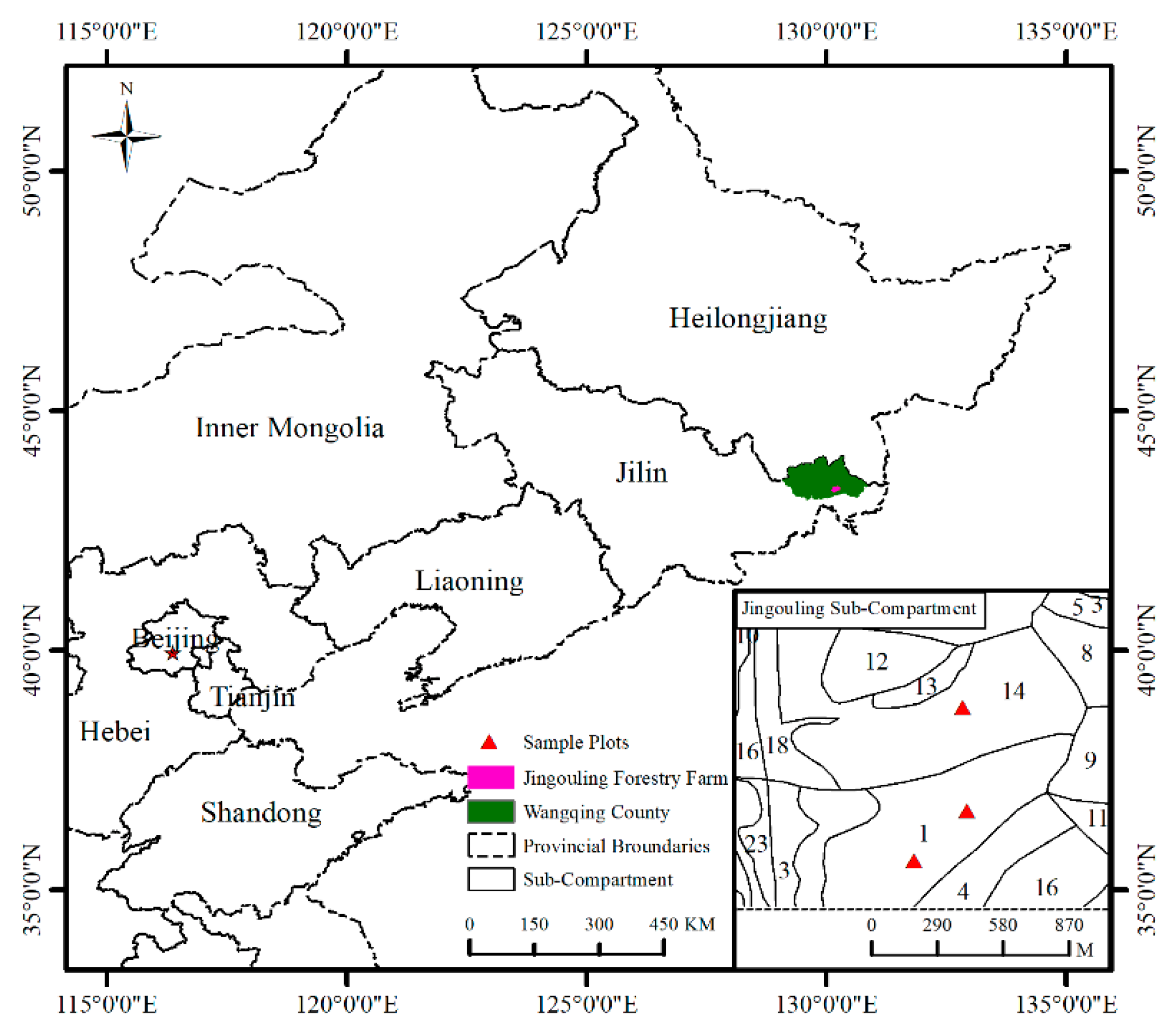
2.1. Study Area and Experimental Design
2.2. Predictor Variables
2.3. Random-Forest Algorithm for Predicting Stand Basal Area Increment (BAI)
3. Results
3.1. Random Forest Model Evaluation
3.2. The Relative Importance (%) of Predictors
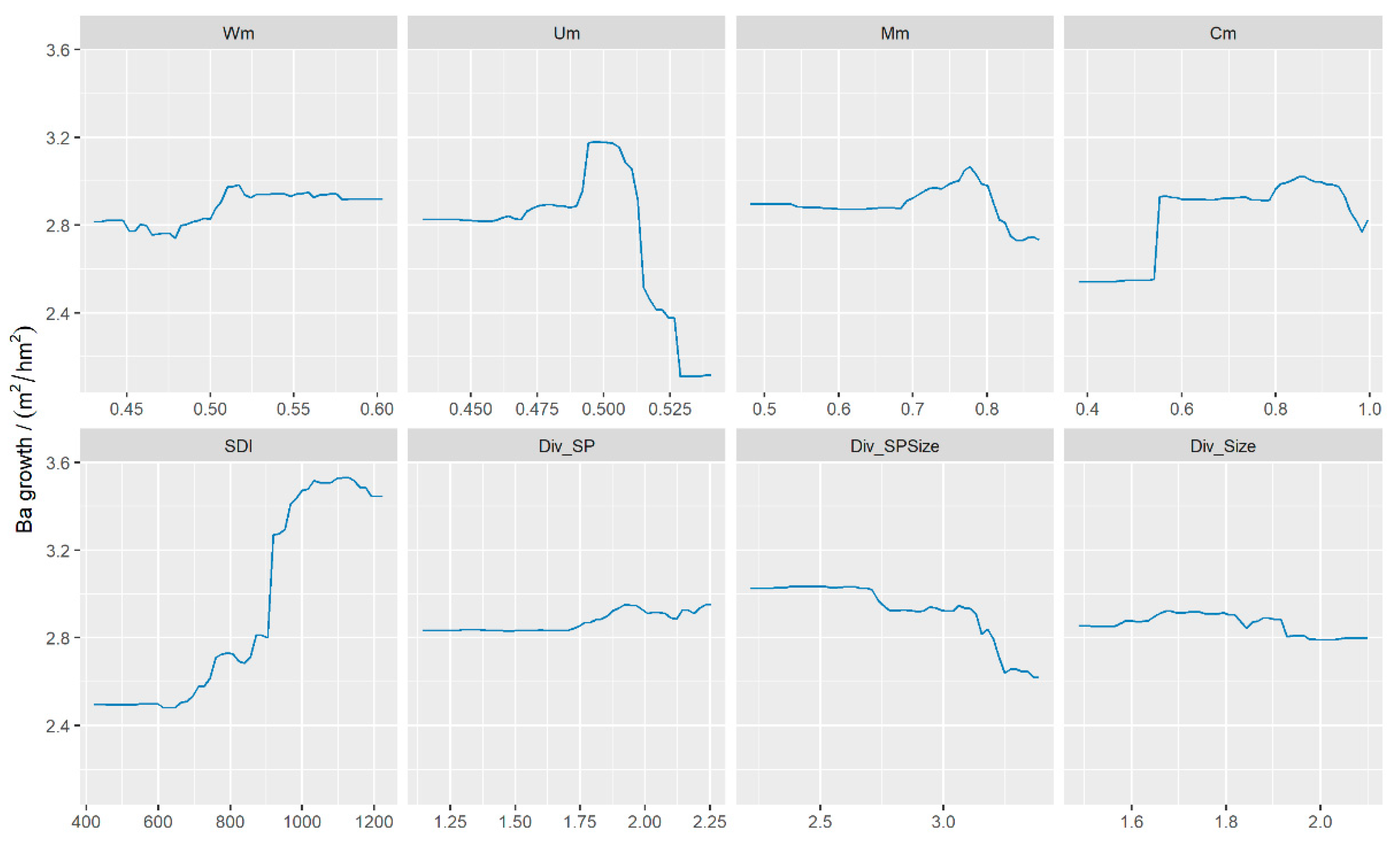
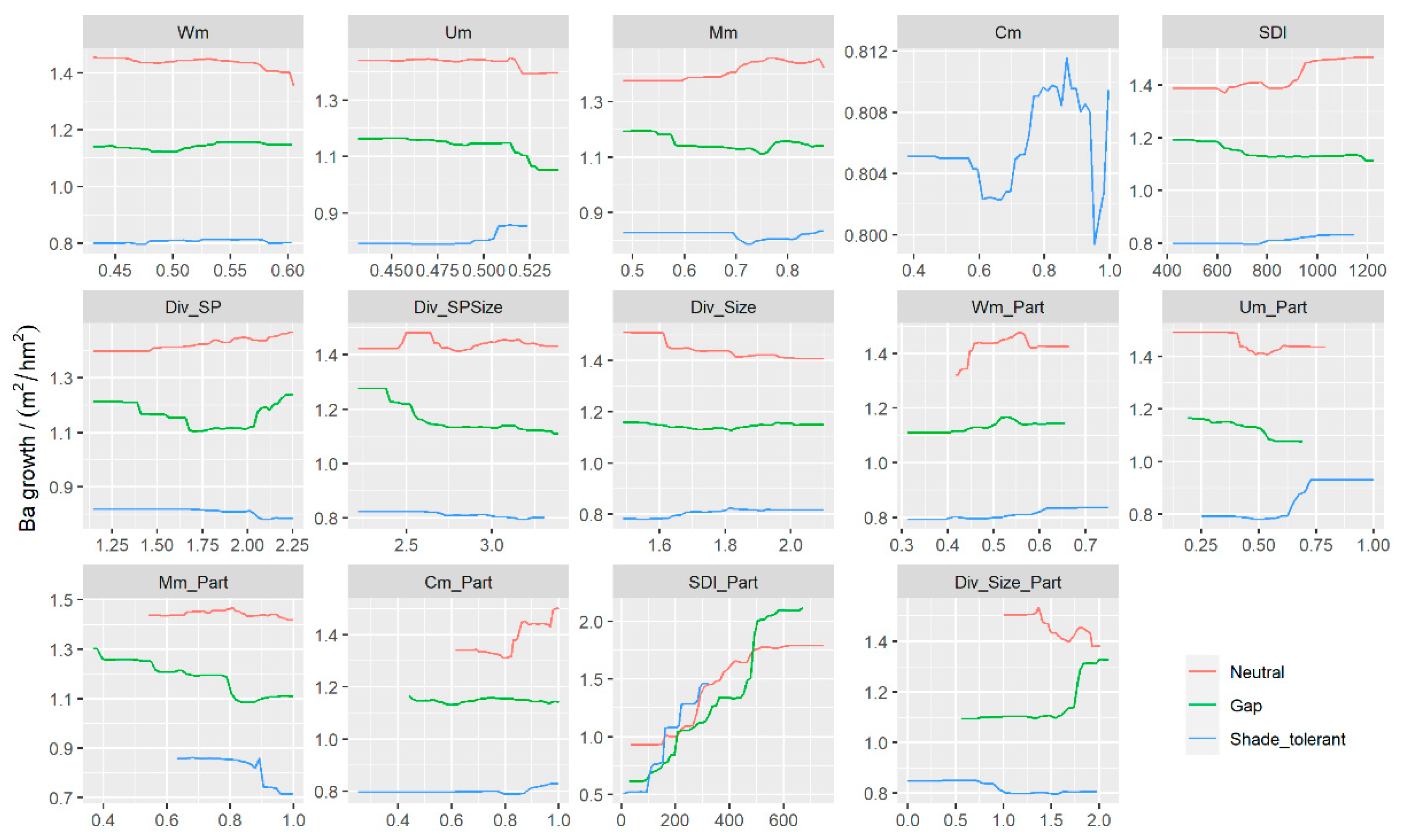
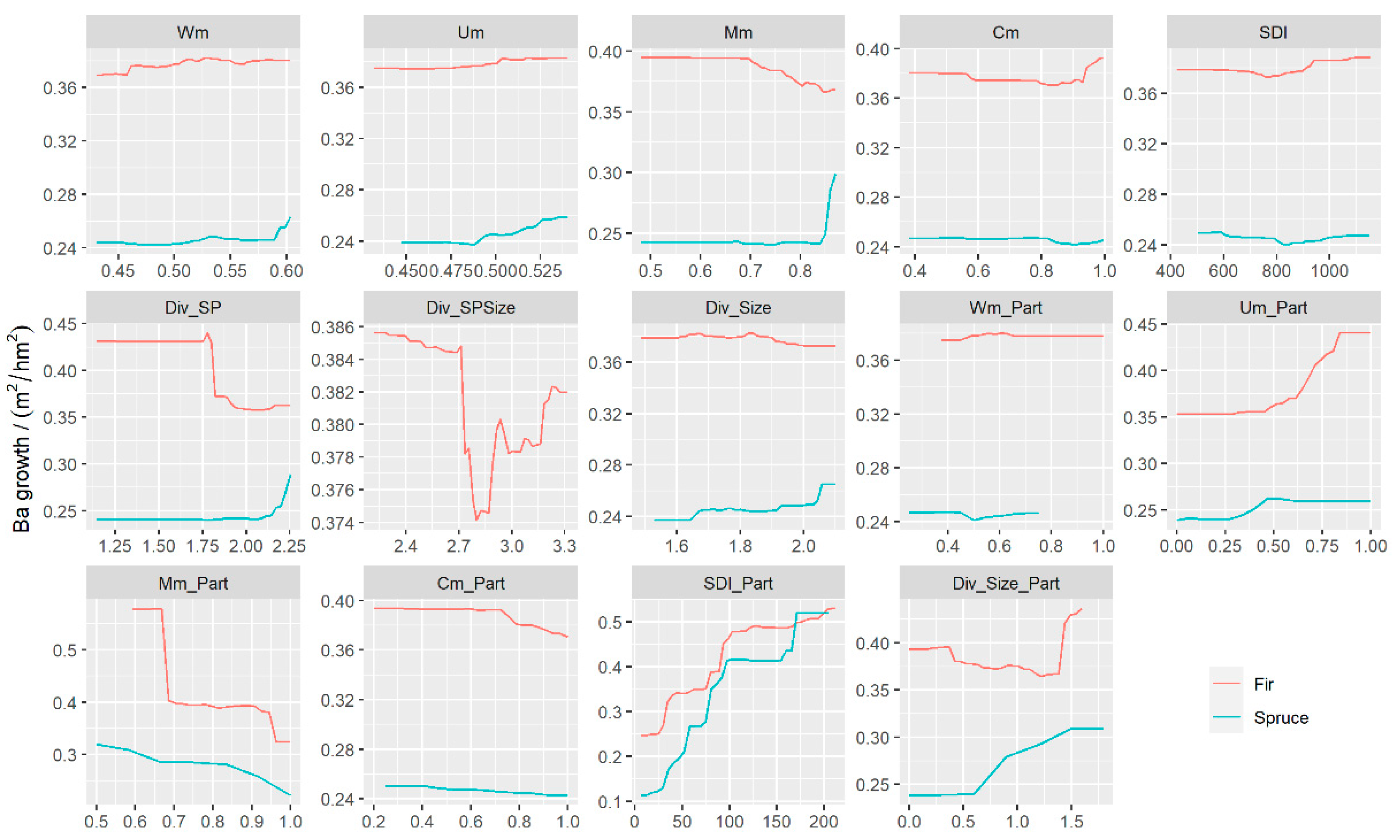
3.3. The Effects of Predictors on BAI
4. Discussion
4.1. Evaluation of Random-Forest Model
4.2. The Effects of STAND Density and Biodiversity on BAI
4.3. Simulated Effect of Forest Spatial Structure Variables on BAI Prediction
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Schlamadinger, B.; Bird, N.; Johns, T.; Brown, S.; Yamagata, Y. A synopsis of land use, land-use change and forestry (lulucf) under the kyoto protocol and marrakech accords. Environ. Sci. Policy 2007, 10, 271–282. [Google Scholar] [CrossRef]
- Jevšenak, J.; Skudnik, M. A random forest model for basal area increment predictions from national forest inventory data. For. Ecol. Manag. 2021, 479, 118601. [Google Scholar] [CrossRef]
- Pukkala, T. Predicting diameter growth in even-aged scots pine stands with a spatial and non-spatial model. Silva Fenn. 1989, 23, 101–116. [Google Scholar] [CrossRef]
- Gschwantner, T.; Alberdi, I.; Balázs, A.; Bauwens, S.; Bender, S.; Borota, D.; Bosela, M.; Bouriaud, O.; Cañellas, I.; Donis, J.; et al. Harmonisation of stem volume estimates in european national forest inventories. Ann. For. Sci. 2019, 76, 24. [Google Scholar] [CrossRef] [Green Version]
- Gleason, C.; Im, J. Forest biomass estimation from airborne lidar data using machine learning approaches. Remote Sens. Environ. 2012, 125, 80–91. [Google Scholar] [CrossRef]
- Joseph, M.; Asner, G.; Knapp, D.; Ty, K.; Martin, R.; Christopher, A.; Mark, H.; Dana, C.; Ben, B. A tale of two “forests”: Random forest machine learning aids tropical forest carbon mapping. PLoS ONE 2014, 9, e85993. [Google Scholar]
- Zhao, Z.; Peng, C.; Qi, Y.; Meng, F.; Zhu, Q. Model prediction of biome-specific global soil respiration from 1960 to 2012: Biome-specific global rs. Earths Future 2017, 5, 715–729. [Google Scholar] [CrossRef]
- Henrique, N.; Bastos, G.; Raul, H. Artificial intelligence procedures for tree taper estimation within a complex vegetation mosaic in brazil. PLoS ONE 2016, 11, e0154738. [Google Scholar]
- Jevenak, J.; Deroski, S.; Zavadlav, S.; Levani, T. A machine learning approach to analyzing the relationship between temperatures and multi-proxy tree-ring records. Tree-Ring Res. 2018, 74, 210–224. [Google Scholar] [CrossRef]
- Jung, M.; Tautenhahn, S.; Wirth, C.; Kattge, J. Estimating basal area of spruce and fir in post-fire residual stands in central siberia using quickbird, feature selection, and random forests. Procedia Comput. Sci. 2013, 18, 2386–2395. [Google Scholar] [CrossRef] [Green Version]
- Fornara, D.; Tilman, D. Plant functional composition influences rates of soil carbon and nitrogen accumulation. J. Ecol. 2010, 96, 314–322. [Google Scholar] [CrossRef]
- Graz, F. Seasonal photosynthetic response of european beech to severe summer drought: Limitation, recovery and post-drought stimulation. Agric. For. Meteorol. 2016, 220, 83–89. [Google Scholar]
- Forrester, D.; Bauhus, J. A review of processes behind diversity—Productivity relationships in forests. Curr. For. Rep. 2016, 2, 45–61. [Google Scholar] [CrossRef] [Green Version]
- Lei, X.; Wang, W.; Peng, C. Relationships between stand growth and structural diversity in spruce-dominated forests in new brunswick, canada. Can. J. For. Res. 2009, 39, 1835–1847. [Google Scholar] [CrossRef]
- Simon, L. The problem of pattern and scale in ecology. Ecology 1992, 73, 1943–1967. [Google Scholar]
- Zeller, L.; Pretzsch, H. Effect of forest structure on stand productivity in central european forests depends on developmental stage and tree species diversity. For. Ecol. Manag. 2019, 434, 193–204. [Google Scholar] [CrossRef]
- Río, M.; Pretzsch, H.; Alberdi, I.; Bielak, K.; Bravo, F.; Brunner, A.; Condés, S.; Ducey, M.; Fonseca, T.; Lüpke, N.; et al. Characterization of the structure, dynamics, and productivity of mixed-species stands: Review and perspectives. Eur. J. For. Res. 2016, 135, 23–49. [Google Scholar]
- LH, R. Perfecting a stand-density index for evenaged forests. J. Agric. Res. 1933, 46, 627–638. [Google Scholar]
- Landsberg, J.; Waring, R. A generalised model of forest productivity using simplified concepts of radiation-use efficiency, carbon balance and partitioning. For. Ecol. Manag. 1997, 95, 209–228. [Google Scholar] [CrossRef]
- Burkhart, H.E. Comparison of maximum size–density relationships based on alternate stand attributes for predicting tree numbers and stand growth. For. Ecol. Manag. 2013, 289, 404–408. [Google Scholar] [CrossRef]
- Dahlhausen, J.; Uhl, E.; Heym, M.; Biber, P.; Ventura, M.; Panzacchi, P.; Tonon, G.; Horváth, T.; Pretzsch, H. Stand density sensitive biomass functions for young oak trees at four different European sites. Trees 2017, 31, 1811–1826. [Google Scholar] [CrossRef]
- Zhang, L.; Hui, G.; Hu, Y.; Zhao, Z. Spatial structural characteristics of forests dominated by pinus tabulaeformis carr. PLoS ONE 2018, 13, e0194710. [Google Scholar] [CrossRef] [Green Version]
- Chanthorn, W.; Hartig, F.; Brockelman, W. Structure and community composition in a tropical forest suggest a change of ecological processes during stand development. For. Ecol. Manag. 2017, 404, 100–107. [Google Scholar] [CrossRef]
- Zhang, G.; Hui, G.; Zhao, Z.; Hu, Y.; Wang, H. Composition of basal area in natural forests based on the uniform angle index. Ecol. Inform. 2018, 45, 1–8. [Google Scholar] [CrossRef]
- Brown, C.; Law, R.; Illian, J.; Burslem, D. Linking ecological processes with spatial and non-spatial patterns in plant communities. J. Ecol. 2011, 99, 1402–1414. [Google Scholar] [CrossRef]
- Franklin, J.; Spies, T.; Pelt, R.; Carey, A.; Thornburgh, D.; Berg, D.; Lindenmayer, D.; Hardo, M.; Keeton, W.; Shaw, D. Disturbances and structural development of natural forest ecosystems with silvicultural implications, using douglas-fir forests as an example. For. Ecol. Manag. 2002, 155, 399–423. [Google Scholar] [CrossRef]
- Pommerening, A.; Stoyan, D. Reconstructing spatial tree point patterns from nearest neighbour summary statistics measured in small subwindows. Can. J. For. Res. 2008, 38, 1110–1122. [Google Scholar] [CrossRef] [Green Version]
- Hui, G.; Gadow, K. The neighbourhood pattern-a new structure parameter for describing distribution of foerst tree position. Sci. Silvae Sin. 1999, 35, 37–42. [Google Scholar]
- Hui, G.; Gadow, K.; Albert, M. A new parameter for stand spatial structure neighbourhood comparison. For. Res. 1999, 1, 4–9. [Google Scholar]
- Hui, G.; Hu, Y. Measuring species spatial isolation in mixed forests. For. Res. 2001, 14, 23–27. [Google Scholar]
- Hu, Y.; Hui, G. How to describe the crowding degree of trees based on the relationship of neighboring trees. J. Beijing For. Univ. 2015, 9, 1–8. [Google Scholar]
- Hui, G.; Zhang, G.; Zhao, Z.; Yang, A. Methods of forest structure research: A review. Curr. For. Rep. 2019, 5, 142–154. [Google Scholar] [CrossRef]
- Wan, P.; Zhang, G.; Wang, H.; Zhao, Z.; Hu, Y.; Zhang, G.; Hui, G.; Liu, W. Impacts of different forest management methods on the stand spatial structure of a natural Quercus aliena var. acuteserrata forest in xiaolongshan, China. Ecol. Inform. 2019, 50, 86–94. [Google Scholar] [CrossRef]
- Hubbell, S.; Ahumada, J.; Condit, R.; Foster, R. Local neighborhood effects on long-term survival of individual trees in a neotropical forest. Ecol. Res. 2001, 16, 859–875. [Google Scholar] [CrossRef]
- Stoll, P.; Newbery, D. Evidence of species-specific neighborhood effects in the dipterocarpaceae of a bornean rain forest. Ecology 2005, 86, 3048–3062. [Google Scholar] [CrossRef] [Green Version]
- Pommerening, A.; Stoyan, D. Edge-correction needs in estimating indices of spatial forest structure. Can. J. For. Res. 2006, 36, 1723–1739. [Google Scholar] [CrossRef] [Green Version]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Ou, Q.; Lei, X.; Shen, C. Individual tree diameter growth models of larch–spruce–fir mixed forests based on machine learning algorithms. Forests 2019, 10, 187. [Google Scholar] [CrossRef] [Green Version]
- Liaw, A.; Wiener, M. Classification and regression by randomforest. R News 2001, 2, 18–23. [Google Scholar]
- Monserud, R.; Sterba, H. A basal area increment model for individual trees growing in even- and uneven-aged forest stands in austria. For. Ecol. Manag. 1996, 80, 57–80. [Google Scholar] [CrossRef]
- Forrester, D. Linking forest growth with stand structure: Tree size inequality, tree growth or resource partitioning and the asymmetry of competition. For. Ecol. Manag. 2019, 447, 139–157. [Google Scholar] [CrossRef]
- Ni, R.; Baiketuerhan, Y.; Zhang, C.; Zhao, X.; Gadow, K. Analysing structural diversity in two temperate forests in northeastern china. For. Ecol. Manag. 2014, 316, 139–147. [Google Scholar] [CrossRef]
- Boris, Z. Optimal stand density: A solution. Can. J. For. Res. 2004, 34, 846–854. [Google Scholar]
- Liang, J.; Buongiorno, J.; Monserud, R.A.; Kruger, E.L.; Zhou, M. Effects of diversity of tree species and size on forest basal area growth, recruitment, and mortality. For. Ecol. Manag. 2007, 243, 116–127. [Google Scholar] [CrossRef]
- Legendre, P.; Mi, X.; Ren, H.; Ma, K.; Yu, M.; Sun, I.F.; He, F. Partitioning beta diversity in a subtropical broad-leaved forest of china. Ecology 2009, 90, 663–674. [Google Scholar] [CrossRef] [Green Version]
- Porté, A.; Bartelink, H. Modelling mixed forest growth: A review of models for forest management. Ecol. Model. 2002, 150, 141–188. [Google Scholar] [CrossRef]
- Pastorella, F.; Paletto, A. Stand structure indices as tools to support forest management: An application in trentino forests (Italy). J. For. Sci. 2013, 59, 159–168. [Google Scholar] [CrossRef] [Green Version]
- Ruprecht, H.; Dhar, A.; Aigner, B.; Oitzinger, G.; Vacik, K. Structural diversity of english yew (Taxus baccata L.) populations. Eur. J. For. Res. 2010, 129, 189–198. [Google Scholar] [CrossRef]
- Bieng, M.; Perot, T.; Coligny, F.; Goreaud, F. Spatial pattern of trees influences species productivity in a mature oak-pine mixed forest. Eur. J. For. Res. 2013, 132, 841–850. [Google Scholar] [CrossRef] [Green Version]
- Davies, O.; Pommerening, A. The contribution of structural indices to the modelling of sitka spruce (Picea sitchensis) and birch (Betula spp.) crowns. For. Ecol. Manag. 2008, 256, 68–77. [Google Scholar] [CrossRef]
- Li, Y.; Zhao, Z.; Hui, G.; Hu, Y.; Ye, S. Spatial structural characteristics of three hardwood species in Korean pine broad-leaved forest—Validating the bivariate distribution of structural parameters from the point of tree population. For. Ecol. Manag. 2014, 314, 17–25. [Google Scholar] [CrossRef]
- Dan, B. A hypothesis about the interaction of tree dominance and stand production through stand development. For. Ecol. Manag. 2004, 190, 265–271. [Google Scholar]
- Li, Y.; Ye, S.; Hui, G.; Hu, Y.; Zhao, Z. Spatial structure of timber harvested according to structure-based forest management. For. Ecol. Manag. 2014, 322, 106–116. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot Code | Stem Number /ha | Mean DBH/ cm | Basal Area/ (m2/ha) | Stock Volume/ (m3/ha) | Canopy Density | Ba Growth in 5 Years/ (m2/ha) |
|---|---|---|---|---|---|---|
| 1 | 996 | 17.63 | 24.30 | 199.97 | 0.85 | 1.85 |
| 2 | 1024 | 18.23 | 26.72 | 216.00 | 0.86 | 2.12 |
| 3 | 1018 | 17.38 | 24.15 | 182.75 | 0.63 | 2.55 |
| Variable Name | Formula | Description | |
|---|---|---|---|
| Biodiversity | Tree species diversity | Ni is the number of trees in the i-th tree species; N is the total number of trees. | |
| Tree size diversity | Nj is the number of trees in the j-th diameter class; N is the same as above. When Nj and N are the number of trees in j-th diameter class and the total of a tree species group or a tree species, respectively, the index is going to be Div_Size_Part. | ||
| Integrated diversity index of tree species and size | Nij is the number of trees of the j-th diameter class in the i-th tree species; N is same as above. | ||
| Stand density | Reineke’s stand density index | N is same as above; D is the actual average diameter of the stand; D0 is the standard average diameter of 20 cm; b is the natural thinning slope coefficient of 1.605. When D and N are the actual average diameter and the number of trees of a tree species group or a tree species, respectively, the index is going to be SDI_Part. | |
| Spatial-structure indices | Uniform angle index | M is the number of target trees; i is the i-th target tree; j is the j-th adjacent tree of the i-th target tree; zij is the judgment result of angles with standard angle; αj is the j-th angle formed by the j-th adjacent tree and the previous one; α0 is the standard angle of 72°. When M is the number of target trees of a tree species group or a tree species, the index is going to be Wm_Part. | |
| Dominance | M, i, and j are same as above; kij represents the judgment result of diameters; DBHj is the DBH of the j-th adjacent tree; DBHi is the DBH of the i-th object tree. When M is the number of target trees of a tree species group or a tree species, the index is going to be Um_Part. | ||
| Mingling | M, i, and j are same as above; vij is the judgment result of tree species codes; spj is the tree species code of the j-th adjacent tree; spi is the tree species code of the i-th object tree. When M is the number of target trees of a tree species group or a tree species, the index is going to be Mm_Part. | ||
| Crowding | M, i, and j are same as above; yij is the judgment result of two crown radii with distance; cj is the crown radius of the j-th adjacent tree; ci is the crown radius of the i-th object tree; distij is the distance between the j-th adjacent tree and the i-th object tree. When M is the number of target trees of a tree species group or a tree species, the index is going to be Cm_Part. |
| Species Group | Tree Species |
|---|---|
| Gap | Asian white birch (Betula platyphylla), Korean pine (Pinus koraiensis), Changbai larch (Larix olgensis), Ussuri popular (Populus ussuriensis), elm (Ulmus japonica). |
| Neutral | Linden (Tilia amurensis), ribbed birch (Betula costata), maple (Acer mono), ash (Fraxinus mandschurica). |
| Shade_tolerant | Corktree (Phellodendron amurense), fir (Abies nephrolepis), spruce (Picea jezoensis). |
| Groups | mtry | R2 ± std | RMSE ± std (m2/ha) |
|---|---|---|---|
| Whole stand | 8 | 0.223 ± 0.468 | 0.534 ± 0.132 |
| Gap | 8 | 0.722 ± 0.207 | 0.236 ± 0.063 |
| Neutral | 10 | 0.622 ± 0.306 | 0.231 ± 0.066 |
| Shade_tolerant | 14 | 0.609 ± 0.295 | 0.167 ± 0.074 |
| Spruce | 11 | 0.730 ± 0.214 | 0.061 ± 0.038 |
| Fir | 14 | 0.575 ± 0.282 | 0.157 ± 0.070 |
| Predictors | Whole Stand | Gap | Neutral | Shade_Tolerant | Spruce | Fir |
|---|---|---|---|---|---|---|
| Wm | 11.61 | 2.27 | 3.01 | 2.29 | 2.19 | 4.74 |
| Um | 21.18 | 2.91 | 3.33 | 4.61 | 2.21 | 2.90 |
| Mm | 8.76 | 3.96 | 4.17 | 3.10 | 3.80 | 8.36 |
| Cm | 10.30 | / | / | 1.62 | 1.12 | 3.44 |
| SDI | 21.83 | 2.45 | 5.74 | 2.44 | 1.53 | 8.43 |
| Div_SP | 6.66 | 7.51 | 3.65 | 2.33 | 3.08 | 6.21 |
| Div_SPSize | 10.63 | 4.76 | 5.84 | 1.95 | / | 3.27 |
| Div_Size | 9.02 | 2.17 | 3.60 | 2.92 | 2.86 | 1.84 |
| SDI_part | / | 50.69 | 38.28 | 53.27 | 52.58 | 24.98 |
| Div_Size_part | / | 7.97 | 7.83 | 3.07 | 11.50 | 6.34 |
| Wm_part | / | 3.02 | 5.80 | 2.85 | 0.94 | 1.34 |
| Um_part | / | 2.40 | 5.29 | 8.56 | 4.49 | 8.18 |
| Mm_part | / | 6.84 | 5.03 | 8.04 | 12.73 | 15.37 |
| Cm_part | / | 3.07 | 8.42 | 2.97 | 0.97 | 4.61 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, D.; Zhou, C.; He, X.; Zhang, X.; Feng, L.; Zhang, H. The Effect of Stand Density, Biodiversity, and Spatial Structure on Stand Basal Area Increment in Natural Spruce-Fir-Broadleaf Mixed Forests. Forests 2022, 13, 162. https://doi.org/10.3390/f13020162
Liu D, Zhou C, He X, Zhang X, Feng L, Zhang H. The Effect of Stand Density, Biodiversity, and Spatial Structure on Stand Basal Area Increment in Natural Spruce-Fir-Broadleaf Mixed Forests. Forests. 2022; 13(2):162. https://doi.org/10.3390/f13020162
Chicago/Turabian StyleLiu, Di, Chaofan Zhou, Xiao He, Xiaohong Zhang, Linyan Feng, and Huiru Zhang. 2022. "The Effect of Stand Density, Biodiversity, and Spatial Structure on Stand Basal Area Increment in Natural Spruce-Fir-Broadleaf Mixed Forests" Forests 13, no. 2: 162. https://doi.org/10.3390/f13020162
APA StyleLiu, D., Zhou, C., He, X., Zhang, X., Feng, L., & Zhang, H. (2022). The Effect of Stand Density, Biodiversity, and Spatial Structure on Stand Basal Area Increment in Natural Spruce-Fir-Broadleaf Mixed Forests. Forests, 13(2), 162. https://doi.org/10.3390/f13020162