
Gemmamyces piceae Bud Blight Damage in Norway Spruce (Picea abies) and Colorado Blue Spruce (Picea pungens) Forest Stands

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
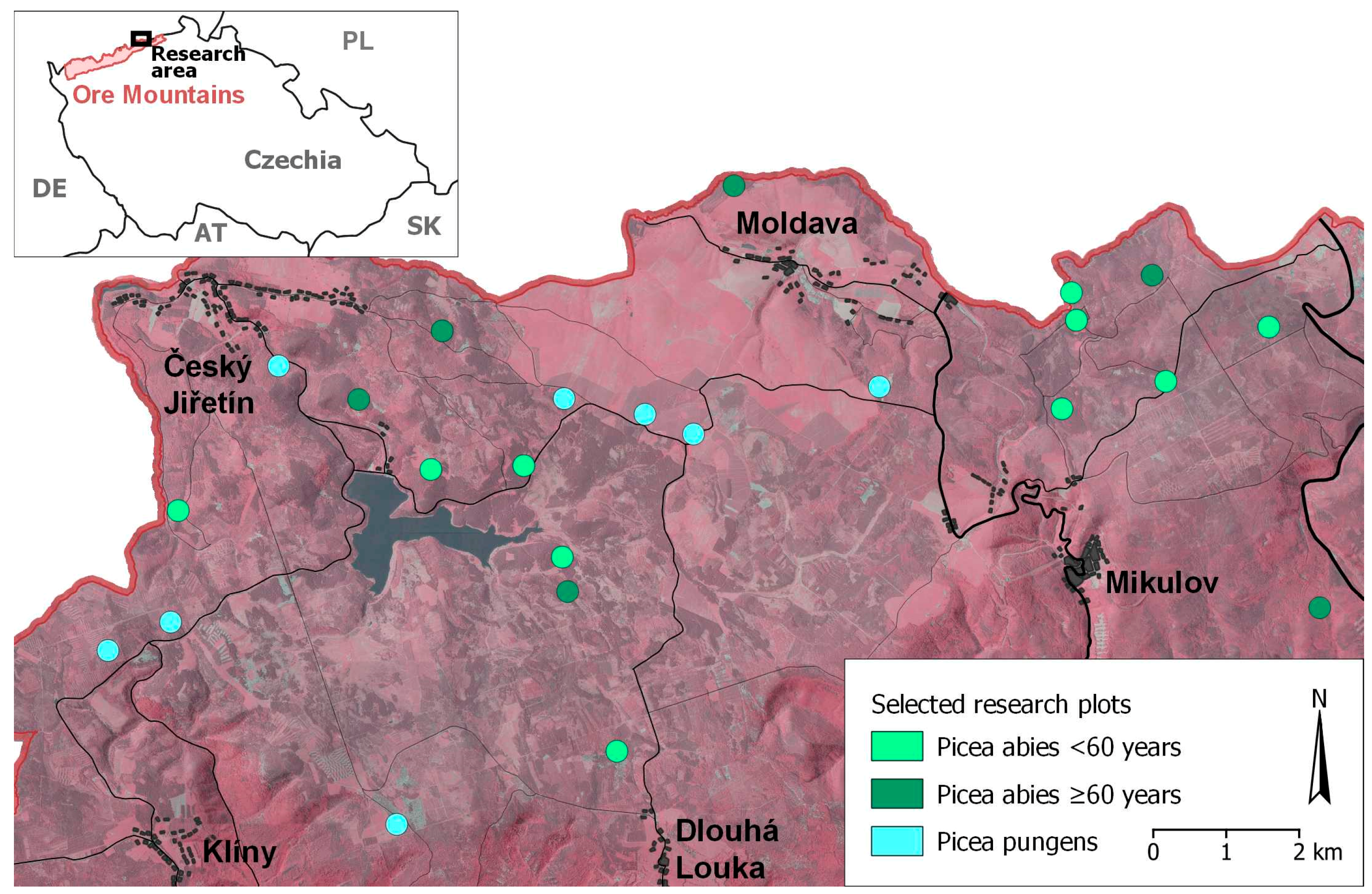
2. Materials and Methods
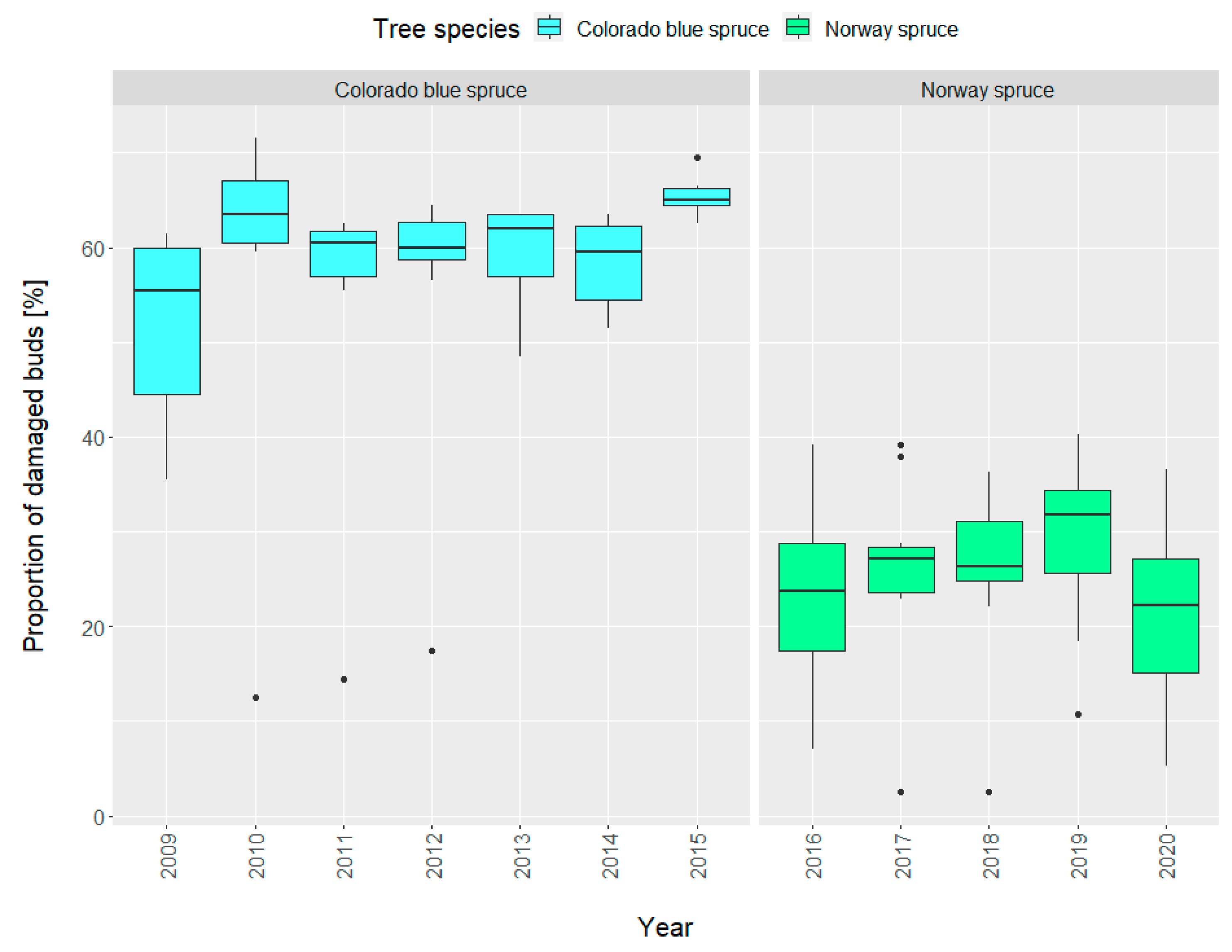
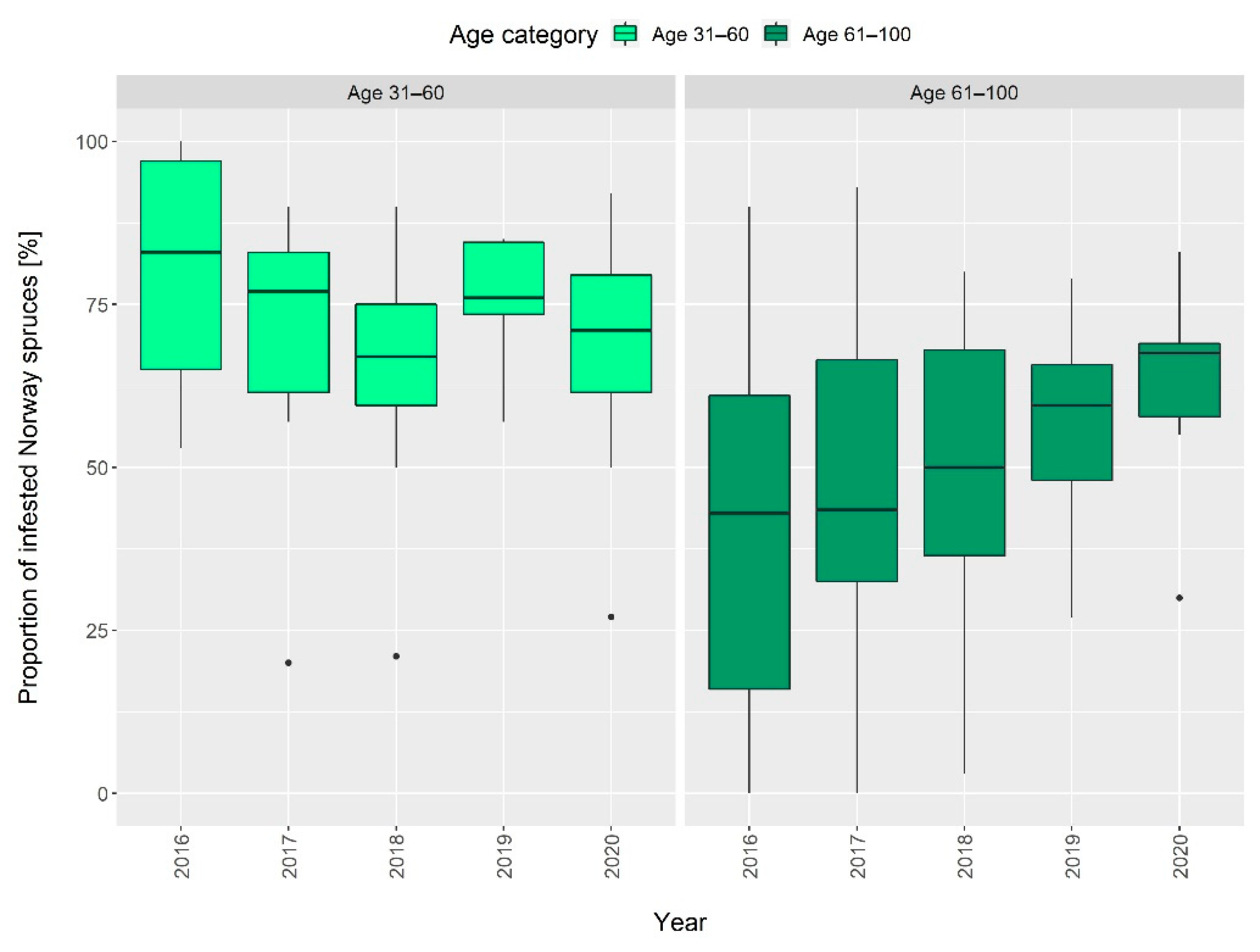
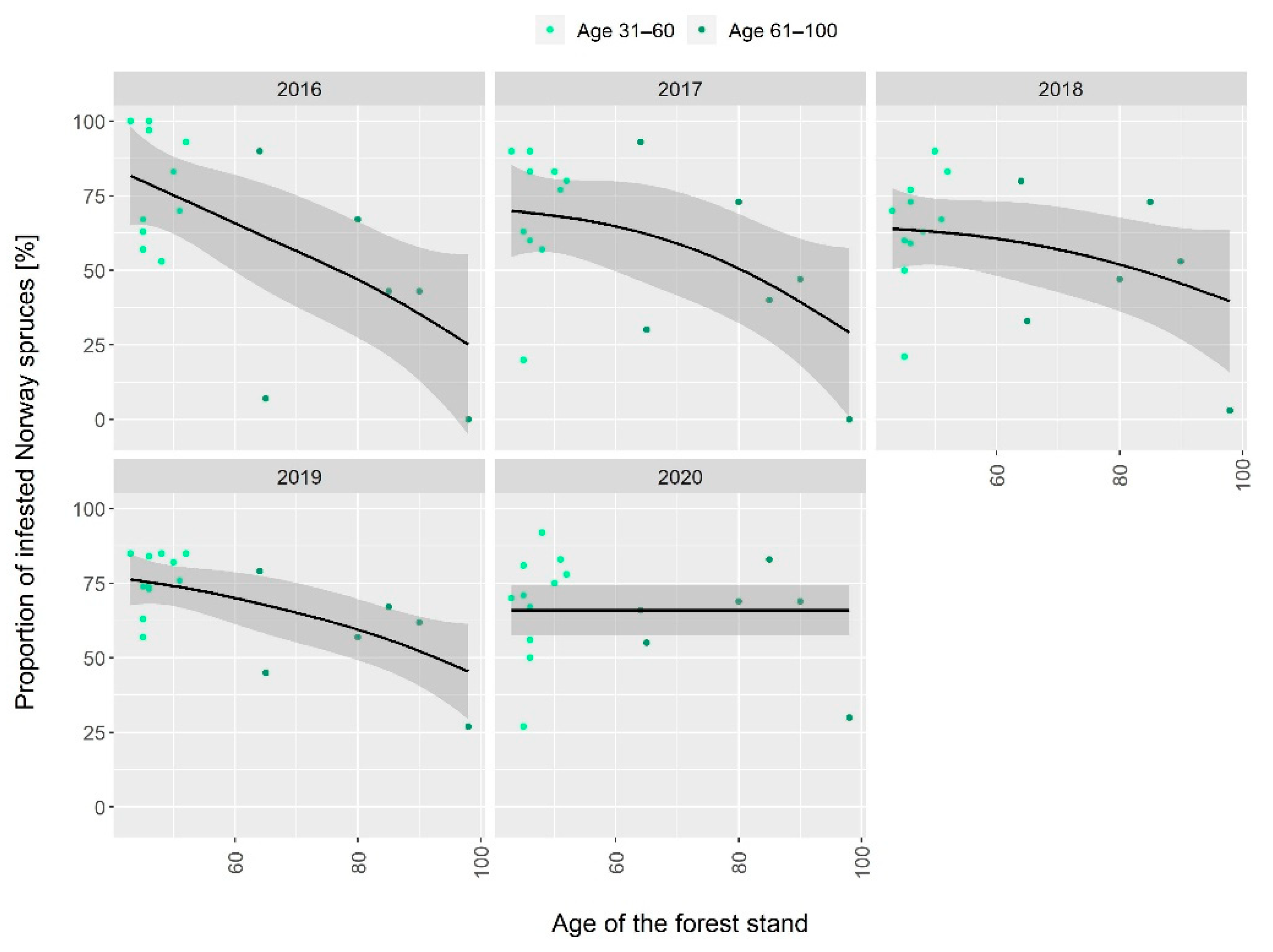
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kandler, O.; Innes, J.L. Air pollution and forest decline in Central Europe. Environ. Pollut. 1995, 90, 171–180. [Google Scholar] [CrossRef]
- Fuhrer, J. Formation of secondary air pollutants and their occurrence in Europe. Experientia 1985, 41, 286–301. [Google Scholar] [CrossRef]
- Ulrich, B. Effects of air pollution on forest ecosystems and waters. The principles demonstrated at a case study in central Europe. Atmos. Environ. 1984, 18, 621–628. [Google Scholar] [CrossRef]
- Lomský, B.; Materna, J.; Pfanz, H. Výzkumný ústav lesního hospodářství a myslivosti: Jíloviště-Strnady? In SO2—Pollution and Forests Decline in the Ore Mountains, 1st ed.; Forestry and Game Management Research Institute: Jíloviště, Czech Republic, 2002; 342p, ISBN 80-86461-24-6. [Google Scholar]
- Nožička, J. Kouřové škody v našich lesích a boj proti nim do roku 1918. Práce VÚL Čssr 1963, 26, 237–258. [Google Scholar]
- Stoklasa, J. Beschädingung der Vegetation durch Rauchgase und Fabrksexhalationen. Urban Und. Schwanzenberg Berl. 1923, 487. [Google Scholar]
- Haselhoff, E.; Lindau, G. Die Beschädigung der Vegetation durch Rauch. Leipzig 1903, 412. [Google Scholar] [CrossRef] [Green Version]
- Slodičák, M. Pěstování porostů náhradních dřevin. In: Problematika zachování porostů náhradních dřevin v imisní oblasti Krušných hor. Sborník Ref. Z Celostátní Konf. Most 1999, 33–40. [Google Scholar]
- Slodičák, M.; Balcar, V.; Novák, J.; Lomský, B.; Šrámek, V.; Navrátil, P.; Kacálek, D.; Mauer, O.; Kula, E.; Podrázský, V.; et al. Lesnické hospodaření v Krušných horách. In LČR, VÚLHM: Hradec Králové, Strnady, Edice Grantové služby LČR Czech Republic; Česká zemědělská univerzita v Praze: Kamýcká, Czech Republic, 2008; p. 480. ISBN 978-80-86945-04-0. [Google Scholar]
- Šrámek, V.; Balcar, V.; Buriának, V.; Havránek, F.; Jurásek, A.; Liška, J.; Novák, M.; Slodičák, M. Aktualizace Studie. Lesnické Hospodaření v Krušných Horách, 1st ed.; Jíloviště, Výzkumný Ústav Lesního Hospodářství a Myslivosti, v.v.i.: Strnady, Czech Republic, 2015; p. 195. [Google Scholar]
- Vacek, S.; Hůnová, I.; Vacek, Z.; Hejcmanová, P.; Podrázský, V.; Král, J.; Moser, W.K. Effects of air pollution and climatic factors on Norway spruce forests in the Orlické hory Mts.(Czech Republic), 1979–2014. Eur. J. For. Res. 2015, 134, 1127–1142. [Google Scholar] [CrossRef]
- Gehring, C.A.; Mueller, R.C.; Haskins, K.E.; Rubow, T.K.; Whitham, T.G. Convergence in mycorrhizal fungal communities due to drought, plant competition, parasitism and susceptibility to herbivory: Consequences for fungi and host plants. Front. Microbiol. 2014, 5, 306. [Google Scholar] [CrossRef] [Green Version]
- Šulc, M.; Tomášková, I.; Krejzková, A.; Samek, M.; Diuzheva, A.; Hradecký, J.; Pešková, V. Trehalose determination in Norway spruce (Picea abies) roots. Analytics matters. MethodsX 2021, 8, 101280. [Google Scholar] [CrossRef]
- Pešková, V.; Soukup, F. Prosychání a odumírání dřevin. Výskyt lesních škodlivých činitelů v roce 2008 a jejich očekávaný stav v roce 2009. In Zpravodaj Ochrany Lesa. Supplementum; Knížek, M., Ed.; Jíloviště,Výzkumný Ústav Lesního Hospodářství a Myslivosti, v.v.i: Strnady, Czech Republic, 2009; pp. 43–44. ISBN 978-80-7417-011-9. [Google Scholar]
- Pospíšil, F.; Pospíšil, J. Poškození smrku pichlavého kloubnatkou v Krušných horách. Zprav. Ochr. Lesa 2011, 15, 76–79. [Google Scholar]
- Zýka, V.; Černý, K.; Strnadová, V.; Zahradník, D.; Hrabětová, M.; Havrdová, L.; Romportl, D. Predikce Poškození Porostů Smrku Pichlavého Kloubnatkou Smrkovou v Krušných Horách, 1st ed.; Výzkumný Ústav Silva Taroucy Pro Krajinu a Okrasné Zahradnictví, v.v.i.: Průhonice, Czech Republic, 2018; p. 47. ISBN 978-80-87674-29-1. [Google Scholar]
- Borthwick, A.W. A new disease of Picea . In Notes from the Royal Botanic Garden, Edinburgh; Nabu Press: Edinburgh, UK, 1909; pp. 259–261. ISBN 978-1247782461. [Google Scholar]
- Shoemaker, R.A. Cucurbitaria piceae and associated Sphaeropsidales parasitic on spruce buds. Can. J. Bot. 1967, 45, 1243–1248. [Google Scholar] [CrossRef]
- Jaklitsch, W.M.; Voglmayr, H. Three former taxa of Cucurbitaria and considerations on Petrakia in the Melanommataceae. Sydowia 2017, 69, 81. [Google Scholar] [CrossRef] [PubMed]
- Pešková, V.; Soukup, F. Současná situace v porostech smrku pichlavého v Krušných horách. Lesn. Práce 2013, 10, 46–47. [Google Scholar]
- Černý, K.; Pešková, V.; Soukup, F.; Havrdová, L.; Strnadová, V.; Zahradník, D.; Hrabětová, M. Gemmamyces bud blight of Picea pungens: A sudden disease outbreak in Central Europe. Plant Pathol. 2016, 65, 1267–1278. [Google Scholar] [CrossRef] [Green Version]
- Pešková, V.; Modlinger, R.; Tomášková, I.; Samek, M.; Baťa, D.; Lorenc, F.; Dušek, D.; Kacálek, D. Vliv faktorů prostředí na napadení smrku ztepilého kloubnatkou smrkovou a návrh praktických postupů omezujících její šíření. In Závěrečná Zpráva, Grantová Služba Lesy České Republiky, s.p.; Výzkumný Ústav Silva Taroucy Pro Krajinu a Okrasné Zahradnictví, v.v.i.: Jíloviště, Czech Republic, 2020; p. 94. [Google Scholar]
- Köck, G. Ein für Österreich neuer Schädling auf Picea pungens. Osterr. Gart. 1918, 13, 147–148. [Google Scholar]
- Ferdinandsen, C.; Jørgensen, C.A. Skovtræernes Sygdomme. Copenhagen. Den. Gyldendahl. 1938, 570. [Google Scholar]
- Muller, E. Eine neue Krankheit der Stechfichte. Schweiz. Beitr. Zur Dendrol. 1950, 2, 69–72. [Google Scholar]
- Thiringer, J.O. Eine fur Schweden neue Knospenkrankheit auf Koniferen. Goteb. Bot. Tradg. 1964, 27, 1–6. [Google Scholar]
- Schneider, R.; Daebeler, F. Eine fur Deutschland neue Knospenkranheit an Picea pungens var. glauca (Erreger: Cucurbitaria piceae Borthwick). Phytopathol. Z. 1968, 63, 388–392. [Google Scholar] [CrossRef]
- Muskett, A.; Malone, J. Catalogue of Irish Fungi—IV.Ascomycotina. Proc. R. Ir. Academy. Sect. B Biol. Geol. Chem. Sci. 1983, 83B, 151–213. [Google Scholar]
- Tuffen, M.G.; Grogan, H.M. Current, emerging and potential pest threats to Sitka spruce plantations and the role of pest risk analysis in preventing new pest introductions to Ireland. Forestry 2019, 92, 26–41. [Google Scholar] [CrossRef]
- Feige, G.B.; Ale-Agha, N.; Jensen, M.; Christiaans, B.; Kricke, R. New, rare or remarkable microfungi in the Italian Alps (Carnic Alps)—part I—ascomycotina. Commun. Agric. Appl. Biol. Sci. 2004, 69, 457–465. [Google Scholar] [PubMed]
- Tomiczek, C.; Cech, T.L.; Fürst, A.; Hoyer-Tomiczek, U.; Krehan, H.; Perny, B.; Steyrer, G. Waldschutzsituation 2010 in Osterreich. Forstsch. Aktuell 2011, 523. [Google Scholar]
- Ogris, N.; Jurc, D. Tree diseases determined by the reporting, prognostic and diagnostic service for forests in Slovenia 1982–2012. Acta Silvae Ligni 2013, 102, 31–42. [Google Scholar] [CrossRef] [Green Version]
- Tubeuf, C. Schilderungen und Bilder aus nordamerikanischen Wäldern Naturwissenschaftliche Zeitschrift für Forst- und Landwirtschaft. Z. Für Pflanzenkrankh. (Pflanzenpathol.) Und Pflanzenschutz 1919, 17, 1–44. [Google Scholar]
- Redfern, D.; Boswell, R.; Proudfoot, J. Forest condition 1996. Research information note 291. For. Comm. 1997, 6. [Google Scholar]
- Winton, L.; Spruce bud blights. Spruce Bud Blights. Region 10—Forest & Grassland Health. US Forest Service. 2020. Available online: https://www.fs.usda.gov/detailfull/r10/forest-grasslandhealth/?cid=FSEPRD535386 (accessed on 10 July 2020).
- Alford, D.V. (Ed.) Pest and Disease Management Handbook; British Crop Protection Enterprises: Oxford, UK, 2000; p. 624. [Google Scholar] [CrossRef]
- Pešková, V.; Modlinger, R. Stav napadení smrku pichlavého a ztepilého kloubnatkou smrkovou. Lesn. Práce 2015, 3, 48–49. [Google Scholar]
- Soukup, F.; Pešková, V. Gemmamyces piceae (Borthw.) Casagr. kloubnatka smrková. Lesn. Práce 2009, 88, 1–4. [Google Scholar]
- Šefl, J.; Mottlová, V.; Schořálková, I. Bud blight (Gemmamyces piceae) in the eastern part of the Krušné hory Mountains. J. For. Sci. 2020, 66, 309–317. [Google Scholar] [CrossRef]
- Pešková, V.; Modlinger, R.; Soukup, F.; Ručková, J. Nárůst napadení smrku ztepilého kloubnatkou smrkovou v Krušných horách. Lesn. Práce 2016, 95, 46–47. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Elphick, C.S. A protocol for data exploration to avoid common statistical problems. Methods Ecol. Evol. 2010, 1, 3–14. [Google Scholar] [CrossRef]
- Brooks, M.E.; Kristensen, K.; Benthem, K.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Mächler, M.; Bolker, B.M. glmmTMB Balances Speed and Flexibility among Packages for Zero-inflated Generalized Linear Mixed Modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. URL. 2020. Available online: https://www.R-project.org/ (accessed on 12 September 2021).
- Fiala, P.; Reininger, D.; Samek, T. Chemismus půdního prostředí a jehlic smrku ztepilého (Picea abies/L./Karst.) ve vápněných a kontrolních porostech Krušných hor. Zprávy Lesn. Výzkumu 2017, 62, 23–32. [Google Scholar]
- Hadaš, P. Stav imisí a depozicí v přírodní lesní oblasti Krušných hor. Lesn. Práce 2002, 81, 11. [Google Scholar]
- Czech Hydrometeorological Institute. Air Pollution in the Czech Republic in 2019; CHMI: Prague, Czech Republic, 2020; p. 204. ISBN 978-80-7653-013-3. [Google Scholar]
- Baciak, M.; Warmiński, K.; Bęś, A. The effect of selected gaseous air pollutants on woody plants. Leśne Pr. Badaw. 2015, 76, 401–409. [Google Scholar] [CrossRef] [Green Version]
- Paoletti, E.; Schaub, M.; Matyssek, R.; Wieser, G.; Augustaitis, A.; Bastrup-Birk, A.M.; Bytnerowicz, A.; Günthardt-Goerg, M.S.; Müller-Starck, G.; Serengil, Y. Advances of air pollution science: From forest decline to multiple-stress effects on forest ecosystem services. Environ. Pollut. 2010, 158, 1986–1989. [Google Scholar] [CrossRef]
- Kozlowski, T.T. Susceptibility of young tree seedlings to environmental stress—Amer. Nurseryman 1976, 144, 55–59. [Google Scholar]
- Bytnerowicz, A. Physiological aspects of air pollution stress in forests. Phyton 1996, 36, 15–22. [Google Scholar]
- Šefl, J. Kloubnatka smrková (Gemmamyces piceae) ve vybraných porostech v oblasti Flájské přehrady v letech 2010–2012. Studia Oecologica 2013, 1, 59–71. [Google Scholar]
- Kozlowski, T.T.; Kramer, P.J.; Pallardy, S.G. The Physiological Ecology of Woody Plants; Academic Press: Cambridge, MA, USA, 1990; p. 678. ISBN 978-0-12-424160-2. [Google Scholar]
- Peralta, S.M.G.; Adams, G.C.; Winton, L.; Cerny, K.; Everhart, S.E. An outbreak of bud blight disease of spruce (Picea spp.) in Alaska: An epidemiological study of Gemmamyces piceae. Plant Health. 2020. Available online: https://apsnet.confex.com/apsnet/2020/meetingapp.cgi/Paper/16863 (accessed on 17 September 2021).
- Pettersson, J.M. Diseases on Christmas Trees in Southern Sweden and Western North Carolina with Emphasis on Phytophthora Root Rot and Neonectria Canker. Ph.D. Thesis, Southern Swedish Forest Research Centre, Swedish University of Agricultural Sciences, Uppsala, Sweden, 2018; p. 78, ISBN 978-91-7760-170-8. [Google Scholar]
- Fanta, J. Rehabilitating degraded forests in Central Europe into self–sustaining forest ecosystems. Ecol. Eng. 1997, 8, 289–297. [Google Scholar] [CrossRef]
- Klopcic, M.; Poljanec, A.; Gartner, A.; Boncina, A. Factors related to natural disturbances in mountain Norway spruce (Picea abies) forests in the Julian Alps. Ecoscience 2009, 16, 48–57. [Google Scholar] [CrossRef]
- Timmermann, V.; Nagy, N.E.; Hietala, A.M.; Børja, I.; Solheim, H. Progression of ash dieback in Norway related to tree age, disease history and regional aspects. Balt. For. 2017, 23, 150–158. [Google Scholar]
- Chandelier, A.; Helson, M.; Dvorak, M.; Gischer, F. Detection and quantification of airborne inoculum of Hymenoscyphus pseudoalbidus using real-time PCR assays. Plant Pathol. 2014, 63, 1296–1305. [Google Scholar] [CrossRef]
- Funk, A.; Sutton, B.C. A disease of conifer buds in western Canada associated with Dichomera gemmicola n. sp. Can. J. Bot. 1972, 50, 1513–1518. [Google Scholar] [CrossRef]
- Sherwood, P.; Villari, C.; Capretti, P.; Bonello, P. Mechanism of induced susceptibility to Diplodia tip blight in drought-stressed Austrian pine. Tree Physiol. 2015, 35, 549–562. [Google Scholar] [CrossRef] [Green Version]
- Soukup, F.; Pešková, V. Napadení poškozených smrků v Orlických horách houbou Ascocalyx abietina. Lesn. Práce 2000, 79, 472–473. [Google Scholar]
- Tomášková, I. Water use efficiency of Norway spruce with bud blight disease. Acta Hortic. 2018, 1222, 133–136. [Google Scholar] [CrossRef]
- Hansson, P. Gremmeniella abietina in Northern Sweden: Silvicultural Aaspects of Disease Development in the Introduced Pinus contorta and in Pinus sylvestris. Management of Pinus sylvestris Stands Infected by Gremmeniella Abietina. Ph.D. Thesis, Southern Swedish Forest Research Centre, Swedish University of Agricultural Sciences, Uppsala, Sweden, 1998; p. 40. [Google Scholar]
- Sierota, Z.; Grodzki, W.; Szczepkowski, A. Abiotic and biotic disturbances affecting forest health in Poland over the past 30 years: Impacts of climate and forest management. Forests 2019, 10, 75. [Google Scholar] [CrossRef] [Green Version]
- Hansson, P. Susceptibility of different provenances of Pinus sylvestris, Pinus contorta and Picea abies to Gremmeniella abietina. Eur. J. For. Pathol. 1998, 28, 21–32. [Google Scholar] [CrossRef]
- Senn, J. Tree mortality caused by Gremmeniella abietina in a subalpine afforestation in the central Alps and its relationship with duration of snow cover. Eur. J. For. Pathol. 1999, 29, 65–74. [Google Scholar] [CrossRef]
- Stefansson, E.; Sinko, M. Experiments with provenanees of Scots Pine, with special regard to high ground in N. Sweden. Studia For. Suec. 1967, 47, 108. [Google Scholar]
- Pulkkinen, P. Frost hardiness development and lignification of young Norway spruce seedlings of southern and northern Finnish origin. Silva Fenn. 1993, 27, 47–54. [Google Scholar] [CrossRef] [Green Version]
- Pešková, V.; Samek, M.; Modlinger, R.; Véle, A. Původní patogen smrku pichlavého se stává nebezpečím pro naše horské smrčiny. Krkonoše Jizerské Hory 2019, 8, 22–23. [Google Scholar]
- Skovsgaard, J.P.; Thomsen, I.M.; Skovgaard, I.M.; Martinussen, T. Associations among symptoms of dieback in even-aged stands of ash (Fraxinus excelsior L.). For. Pathol. 2010, 40, 7–18. [Google Scholar] [CrossRef]
- Hlásny, T.; Zimová, S.; Merganičová, K.; Štěpánek, P.; Modlinger, R.; Turčáni, M. Devastating outbreak of bark beetles in the Czech Republic: Drivers, impacts, and management implications. For. Ecol. Manag. 2021, 490, 13. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Samek, M.; Modlinger, R.; Bat’a, D.; Lorenc, F.; Vachová, J.; Tomášková, I.; Pešková, V. Gemmamyces piceae Bud Blight Damage in Norway Spruce (Picea abies) and Colorado Blue Spruce (Picea pungens) Forest Stands. Forests 2022, 13, 164. https://doi.org/10.3390/f13020164
Samek M, Modlinger R, Bat’a D, Lorenc F, Vachová J, Tomášková I, Pešková V. Gemmamyces piceae Bud Blight Damage in Norway Spruce (Picea abies) and Colorado Blue Spruce (Picea pungens) Forest Stands. Forests. 2022; 13(2):164. https://doi.org/10.3390/f13020164
Chicago/Turabian StyleSamek, Michal, Roman Modlinger, Daniel Bat’a, František Lorenc, Jana Vachová, Ivana Tomášková, and Vítězslava Pešková. 2022. "Gemmamyces piceae Bud Blight Damage in Norway Spruce (Picea abies) and Colorado Blue Spruce (Picea pungens) Forest Stands" Forests 13, no. 2: 164. https://doi.org/10.3390/f13020164
APA StyleSamek, M., Modlinger, R., Bat’a, D., Lorenc, F., Vachová, J., Tomášková, I., & Pešková, V. (2022). Gemmamyces piceae Bud Blight Damage in Norway Spruce (Picea abies) and Colorado Blue Spruce (Picea pungens) Forest Stands. Forests, 13(2), 164. https://doi.org/10.3390/f13020164