1. Introduction
Ecological research focuses on how environmental conditions and population processes regulate the abundance and distribution of species [
1], which reach their highest abundance at the centre of their range and decrease towards the edges [
2]. In addition, peripheral populations often become more patchy, isolated and transient [
3,
4]. Although abiotic and biotic aspects are not always consistent with geographical components (latitude, longitude and altitude) [
5], the location is considered a key factor in species conservation because it greatly influences the capacity of any living system to persist in response to environmental changes [
6].
Moreover, the centre–periphery theory suggests that peripheral or marginal populations will diverge over time from central or core populations as a result of two important processes: genetic drift and natural selection [
7], because they are more fragmented and less likely to receive immigrants from other populations than central populations [
7,
8]. Thus, marginal areas are ecologically less favourable for species development [
1,
9,
10,
11,
12], and the probability of extinction is therefore relatively high in these areas [
13].
However, in some specific cases, the geographical and ecological areas are not congruent [
5,
7,
9,
10]. This incongruent pattern may be explained by one of the three main hypotheses concerning trends in genetic diversity across the central–peripheral clines, each of which has different spatial implications [
14]. The first hypothesis, developed by Carson [
15], argues that genetic diversity will increase from the periphery of the distribution range towards the centre. Carson suggested that the more continuous, denser and central populations undergo balancing selection and are therefore expected to display higher levels of within-population genetic diversity [
15,
16].
The second hypothesis, proposed by Fisher [
17], predicts that genetic diversity will decrease from the periphery towards the core of the range of distribution of a species. Accordingly, peripheral populations will sustain higher levels of genetic diversity due to fluctuating selection in spatially heterogeneous and unpredictable environments, while central populations will experience stabilising selection, which maintains genetic diversity [
17,
18,
19,
20].
The third hypothesis, proposed by Mayr [
16], considers homogeneous diversity from the periphery to the core and suggests that gene flow from the core may compensate for the effects of local selection and genetic drift at the periphery. In such cases, genetic diversity may be homogeneous throughout the species range [
16,
21,
22].
Numerous studies have tried to test this theory in animals and plants, comparing the genetic diversity in central and peripheral populations of species by using morphological markers, allozymes, RAPD (Random Amplified Polymorphic DNA), ISSR (Inter-simple Sequence Repeats), AFLPs (Amplified Fragment Length Polymorphisms) and microsatellites [
9,
14,
22,
23,
24,
25,
26,
27].
Contrasting results have been obtained. Some authors have concluded that genetic diversity is similar, and differentiation is greater in peripheral than in central populations [
28,
29]. However, in some cases, no evidence was found in peripheral populations for the greater genetic differentiation, lower genetic diversity and asymmetric genetic flow predicted by the central–peripheral hypothesis [
30,
31,
32,
33]. Therefore, results of empirical research remain ambiguous and limited [
8,
32,
34,
35].
One motivation for the conservation of species is to understand the patterns and the processes associated with the geographical variation in the genetic structure of the population, through the distribution ranges of the species of interest [
8,
36,
37,
38,
39,
40]. It is also stated that when a species is in danger of extinction, its geographic range will contract inwards, with central populations persisting until the final stages of decline [
41].
Consequently, the species adaptation will not decrease gradually from the centre towards the periphery but will decrease dramatically once an environmental tipping point is exceeded, as with abrupt changes in climate, land use and/or biotic interactions [
5,
42]. Accordingly, some authors suggest that peripheral populations are also gaining importance for the conservation of genes, as under climate change conditions, they may possess genotypes of future adaptive potential on which natural selection can act [
9,
37,
43].
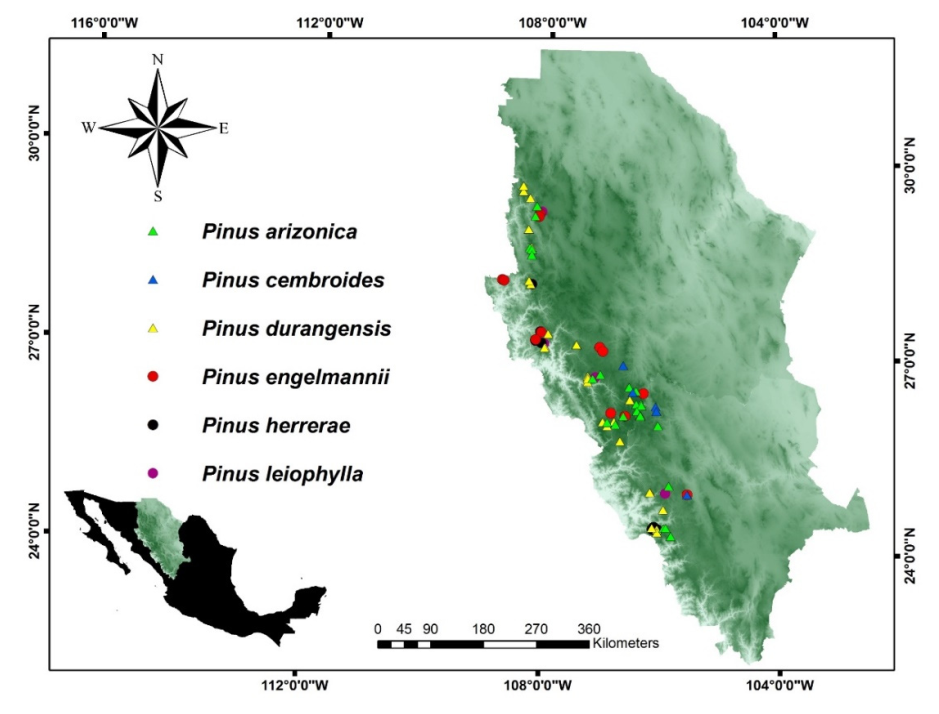
Summarising, populations at the edge of a species distribution range may differ substantially from central populations, in the context of current climate and geography. Therefore, the aim of the present study was to examine whether the centre–periphery theory helps to explain the genetic diversity and differentiation in six of the main pine species in the pine-oak region of the Sierra Madre Occidental (northern Mexico). In this region, the species considered were Pinus arizonica, P. cembroides, P. durangensis, Pinus engelmannii, P. herrerae and P. leiophylla, which are located at the margins and centre of the geographic distribution. Many communities depend on the services and products provided by the forests with these endemic pine species.
4. Discussion
In this study, we determined and evaluated the genetic diversity in 80 populations of six important Pinus spp. located in the centre and periphery of their natural distribution in the Sierra Madre Occidental, in the states of Durango and Chihuahua, Mexico. The main goal of the study was to determine whether the Centre–Periphery Theory could explain the distribution of some pine species, for the purposes of conservation and subsequent sustainable use.
According to the first partitioning criterion (the distance to the closest range edge), there were no significant differences among species´ populations, except for P. herrerae populations. The number of effective genetic variants, the average genetic distances and the elevation were important factors influencing the geographical location and helped to discriminate the central from the peripheral populations. On the other hand, the quartile stratification of elevation (second criterion) did not show significant differences between centre and peripheral populations, except for P. arizonica in which the elevation appeared to be a determining factor in diversity and differentiation measures. However, the hierarchical cluster analysis revealed that genetically, peripheral and central populations of P. arizonica are very similar. Therefore, genetic diversity of the six pine species evaluated is good, and it may be inferred that all the populations assessed were central.
In detail, analysis of 376 loci from AFLPs markers revealed non-significant differences between the genetic diversity of central and peripheral populations of four of the six species analysed (
P. arizonica,
P. cembroides,
P. durangensis and
P. engelmannii), according to proximity to the closest range border (criterion 1). These findings contrast with those of Schwartz et al. [
60], who stated that genetic diversity is expected to be greater in central than in peripheral populations, for two reasons. On the one hand, peripheral populations are usually smaller than central populations, and heterosygosity, allelic diversity and gene flow are therefore lower. On the other hand, genetic variation may be reduced in peripheral populations due to limited and poor connectivity with other populations. In addition, the actual pattern of genetic variation observed may be a result of historical and actual ecological forces [
5,
61,
62,
63], as well as the current population dynamics.
Population dynamics are related to the size of populations and the factors involved in their maintenance, decline or expand considering the limits of the distribution ranges of species related to emerging issues, such as biotic invasions and epidemic diseases under conditions of climate change [
64,
65,
66,
67]. Several studies have focused on native species borders and central–peripheral processes in natural settings [
2,
36,
68,
69]. The Sierra Madre Occidental is home to 46% of the Mexican native pine species [
70], including those evaluated in the present study. However, in four of the six species studied, i.e.,
P. arizonica, P. cembroides, P. durangensis and
P. engelmannii, genetic variables were not correlated with either the central or peripheral location (criterion 1). In the context of climate change and emerging issues, it may be necessary to carry out a further examination of the central–peripheral patterns and processes of invasive species, partly due to their highly dynamic populations and management implications [
64,
67,
68,
71,
72]. This issue may raise questions as to the dynamics of the six
Pinus spp. analysed in the present study and their probability of being invaded by foreign species.
Traditionally, most central–peripheral comparisons, including the present study, have been conducted on native species, especially for conservation purposes [
36,
73,
74,
75,
76,
77]. However, many patterns and processes in central and peripheral populations are often non-linear, as pointed out by Sexton and Dickman [
67]. In addition, marginal populations located in different directions are often structurally and dynamically very different [
64,
67,
78,
79]. Consequently, the underlying mechanisms may be key to explaining the patterns of distribution of at least three of the six
Pinus spp. considered in the present study. The population sizes may also be important factors, as populations of
P. arizonica (
Figure S1) and
P. durangensis (
Figure S3) have higher densities and number of individuals in comparison with the other four species tested, and according to Lázaro-Nogal et al. [
78], population size can at least partly override the effects of geographical periphery.
The highest values of genetic diversity of the six
Pinus species analysed, measured as the effective number of variants
[
52,
80,
81,
82], were (1.387) for
P. arizonica and (1.414) for
P. durangensis. The differences in average gene diversity between central and peripheral populations were not significant for any of the two species under the centre–periphery partitioning (criterion 1). This behaviour may be explained by Mayr’s hypothesis, whereby the gene flow from the core may compensate for the effects of local selection and genetic drift at the periphery. In such cases, genetic diversity may be homogeneous throughout the species range [
16]. Nonetheless, in other cases, such as those mentioned by Pironon et al. [
5], geographical and environmental gradients do not strictly overlap and therefore the abundance should not systematically follow a geographical or ecological central–peripheral pattern [
5,
66].
The rarity index (DW) showed that
P. cembroides was the most strongly differentiated species of the six evaluated, but only five populations of this species were sampled; however, given the distribution range of this pine, these observations were probably insufficient to analyse the dynamics and genetic diversity that contribute to the distribution patterns. Moreover, ecological, and genetic factors are closely related to each other, and their interactions regulate population dynamics over space and time [
83,
84]. For example, for many species, climate conditions can limit species distribution, but whether the species can expand its range (birth > death) may be affected by the genetic pool and evolutionary potential of its component populations, especially peripheral populations [
85,
86].
However, small population size and spatial isolation can promote inbreeding in natural populations, driving excess of homosygote frequencies in subsequent generations [
87,
88], and the possibility that recessive deleterious or lethal alleles will be expressed [
89]. According to criterion 1, this may be the case in species with fewer and smaller populations, where the DW value may be higher and populations occur at higher elevation, as the possible case in
P. leiophylla and
P. cembroides, with four and five populations, respectively. However, the highest DW value of 0.426 in
P. cembroides, together with the high and stable number of mean genetic variants of 1.380 and the 86.6% of polymorphic loci make this species the most differentiated and well-adapted to its growth conditions, across its range. This finding is similar to that of Sexton et al. [
83], who reported that adaptation of individuals in marginal populations may be high if these individuals reside in favourable habitats, even though such habitats may be scarce toward the range edge; therefore, it is important to conserve front-edge populations that may contribute to species adaptation [
68].
These findings contradict the hypothesis that species with small ranges generally exhibit lower genetic variation than more widespread species [
87]. The distribution of
P. cembroides is more consistent with Fisher´s hypothesis, which predicts that peripheral populations sustain higher levels of genetic diversity due to fluctuating selection in spatially heterogeneous and unpredictable environments [
17,
18,
19,
67,
68,
83,
84].
As conifers are self-compatible plants, homosygosity may arise from self-fertilisation and biparental inbreeding, affecting the mating system. A study of reproductive traits and mating systems in
Pinus strobus revealed significantly lower reproductive adaptation in small, peripheral stands than in large, central populations [
88]. Conifers have one of the highest known numbers of lethal equivalents [
89,
90], and an increase in homosygosity is likely to have strong negative effects on offspring. Salzer and Gugerli [
90] found that around 76% of the fully developed seeds collected in the peripheral stands of
Pinus cembra were empty, while seeds from large central populations only showed an average embryo abortion of 30%, which may lead to better survival of individuals in central populations. This scenario provides a possible explanation for the trend observed in the
P. leiophylla species evaluated here using criterion 1.
In the present study, based on criterion 1, the lack of significant differences in genetic diversity between central and peripheral populations in 66% of the species under study indicates the possibility that longevity and large enough population sizes could reduce the negative effects of genetic drift in peripheral populations [
79,
85]. In summary, the absence of statistically significant correlations between genetic variants and geographical distances (criterion 1) was evident in five of the six species under study. Therefore, this distribution pattern was not explained by the general centre–periphery hypothesis [
2,
7,
8]; the same finding has been demonstrated in other studies with conifers [
26,
31] and the endemic
Lilium pomponium [
91].
By contrast, the general centre–periphery hypothesis supported and explained the pattern observed in
P. herrerae [
2,
7,
8,
37], based on criterion 1. Statistically significant correlations between genetic and geographic variants and the different peripheral and central populations were found in this species, showing that the existing diversity and differentiation depended on elevation, as they were slightly more pronounced at higher elevations. Therefore, the establishment of greater number of effective allelic variants will appear in populations occurring at higher elevations, due to the extreme climatic conditions to which they have adapted [
92,
93,
94,
95].
Regarding standard diversity measures, such as FST, and the results of the AMOVA using the AFLP markers, the main source of variation was within populations for all the species evaluated [
58,
93]. In addition, considering the very low FST values, it is likely that two key genetic parameters: the effective number of migrants among populations (Nm) and the rate of gene flow (m), will be homogeneous, and genetic diversity will therefore be evenly distributed in the centre and at the margins of the range. As a result, geographically peripheral populations were not detected in this study; according to criterion 1, populations of the six species studied can be considered central populations because they all exhibit higher genetic diversity and extremely low genetic differentiation.
By contrast, according to results of the non-parametric Kruskal–Wallis test using genetic diversity indexes from Gregorius (1978) [
52] based on criterion 1, we verified that the peripheral populations of
P. herrerae are less diverse and more differentiated than the central population. This finding is consistent with those reported in other studies that also show greater genetic diversity in central populations [
35,
94], and larger genetic differentiation in peripheral populations [
37,
78].
The evolutionary theory suggests that, in virtue of their isolation, peripheral populations should adapt more closely to their environment than central populations. The degree to which a population can adapt to its environment not only depends on the rate of environmental change, but also on how much maladaptive gene flow a population receives [
95]. Therefore, from ecological and evolutionary perspectives, individuals in peripheral populations of
P. herrerae are worth conserving in face of climate change as new arguments for their conservation importance may be provided [
68].
Although we found a correlation between geographic distance (DGEO) and DW for
P. leiophylla, the relationship was not statistically significant because of the small number of observations. However, the observed trend indicates the probable differentiation that may be occurring in this species. The adaptive trajectory of the population types with respect to climate by means of population size, population connectivity and climatic environment may be responsible for this trend [
68,
78].
For criterion 2, by using elevation as a partitioning criterion for distinguishing between central and peripheral populations, a similar pattern was observed in all the populations of the six species studied. There is an ecological gradient from the sampling points, which was adequate for classifying central and peripheral populations derived from elevation.
The current pattern observed in
P. arizonica, in which central populations are more diverse (
Table 8) may be explained by pollination processes and seed dispersion traits. Moreover, populations are widely distributed and sufficiently dense to compensate for any disturbing phenomenon that may occur. In this regard, the altitude range in central populations seems to be optimal. The difference between central and peripheral populations may be due to a lack of homogeneous bidirectional gene flow.
In this study, three groups of plots were visible in the
P. arizonica distribution pattern: two plots of peripheral populations and one of central populations (
Figure 3). However, one peripheral population was more similar to the central group than to the other peripheral group, with distinctly different forest traits in the Sierra Madre Occidental. In addition, forest structural attributes, such as number of trees and average tree size, may be affected by elevation.
Tree species diversity was similar in central and peripheral populations of all the six species evaluated. This general result may indicate that elevation is a better partitioning criterion than the distance to the closest edge. However, in P. herrerae the pattern seems to correspond to both partitioning criteria. In addition, the data trend is related to the elevation, although this species shows a closer relation with the geographic distance (criterion 1).
Regarding diversity, the results are consistent with those of Corral–Rivas et al. [
96], who reported that the diversity profile of forest species may be characterised by a number of attributes, including the distribution of species and sizes of individuals.
The lowest peripheral population recorded belongs to
P. engelmannii, at an elevation of 1820 m; however, diversity was very similar in both central and peripheral populations. This may be a good indicator of adaptation, as studies by Bickford et al. [
97] have shown that increasing frequency and severity of drought predicted for the lowest regions of the Sierra Madre Occidental in the coming decades will reduce growth of
P. engelmannii, with a greater impact on low-elevation populations of
P. engelmannii.
The findings of the present study rule out differences in central and peripheral populations of
P. leiophylla as the diversity measures are very similar in both types of populations at different elevations. These findings are consistent with those reported by Castellanos-Acuña et al. [
98], who reported that growth decreases when populations shift to sites that are colder (at higher elevation) than the sites of origin. However, no differences were found at an elevational shift of 300 m for
P. leiophylla populations. This shift would not lead to severe maladaptation and may thus represent a viable management strategy for
P. leiophylla populations in scenarios of climate change [
98].
Finally, the importance of traits differs depending on ecosystem conditions. One of the basic requirements appears to be grouping species into cohorts with similar traits, such as
P. arizonica and
P. durangensis. This is an important factor to consider for developing more advanced methods of harvest control in the forests in the Sierra Madre Occidental [
99]. Therefore, it is important to know that these two species are genetically diverse and have ample distribution.


 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}