
Prediction of Suitable Distribution of a Critically Endangered Plant Glyptostrobus pensilis

 ,
,
Abstract
:1. Introduction
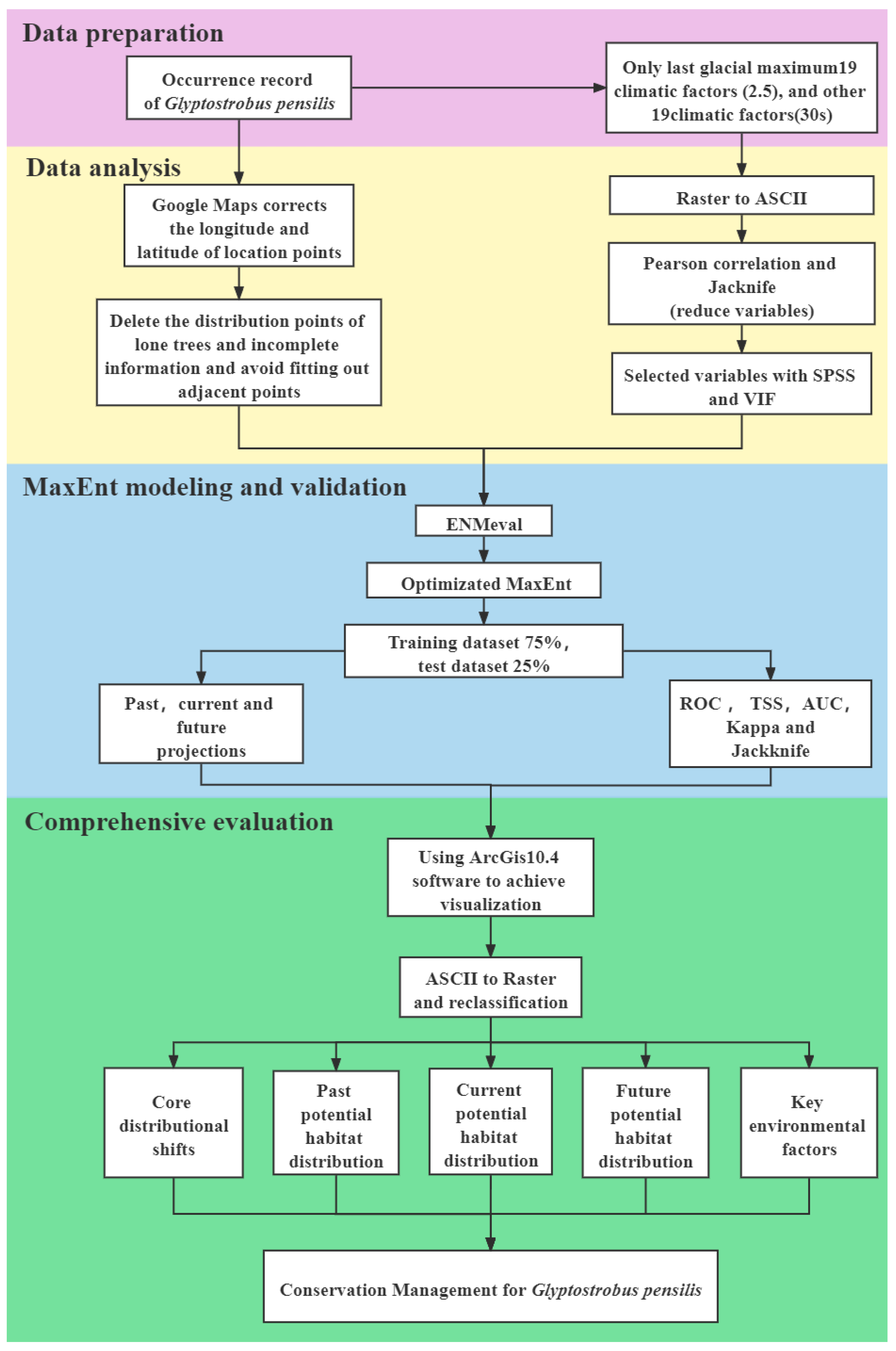
2. Materials and Methods
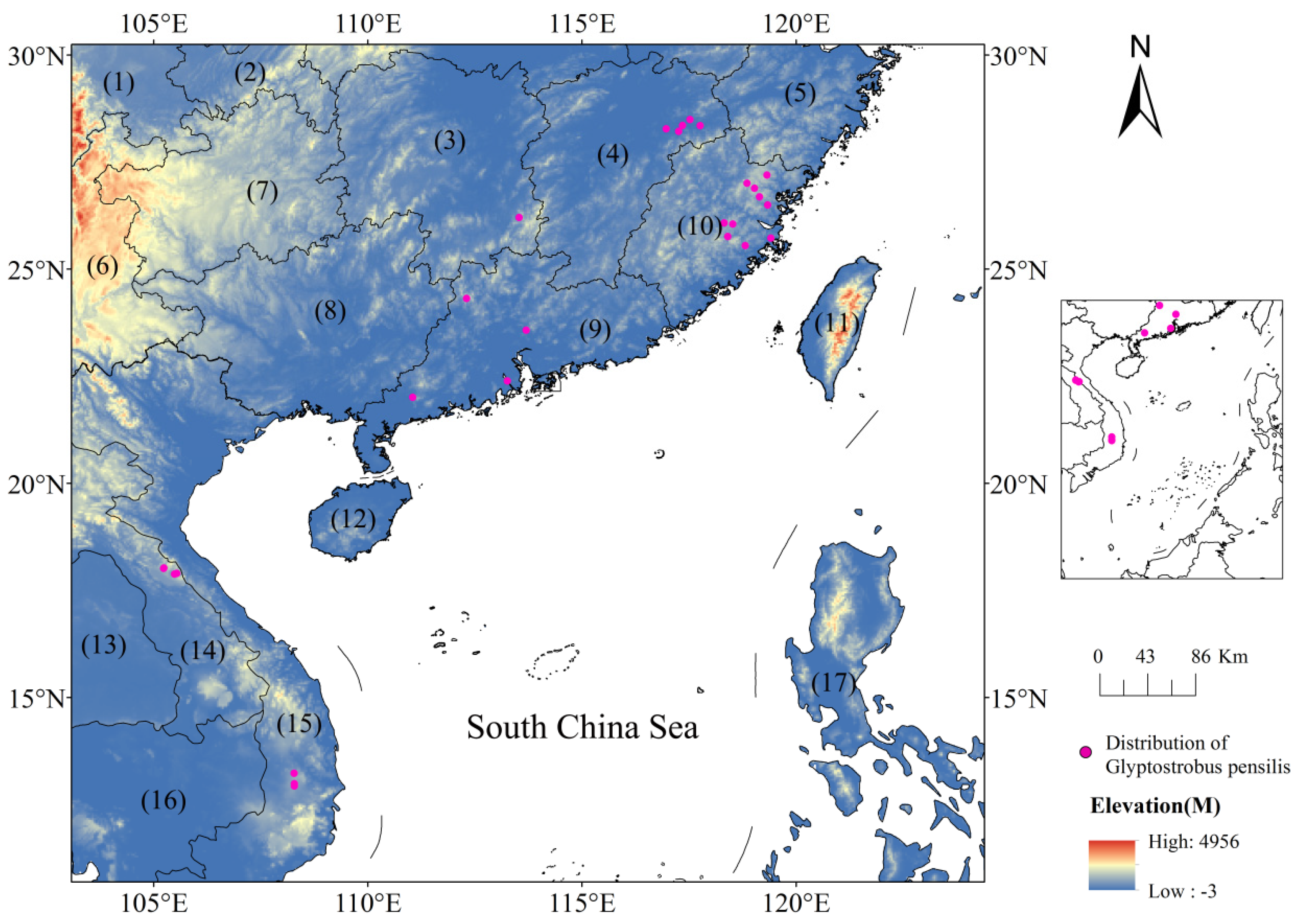
2.1. The Study Area
2.2. Collection and Screening of Sample Data
2.3. Environment Variable Filtering and Data Processing
2.4. Model Building, Optimization and Evaluation
2.5. Analysis of the Spatial Pattern Change and Core Point Migration in the Suitable Habitat Area of Species
3. Results
3.1. Model Optimization and Accuracy Evaluation
3.2. Importance of Environment Variables
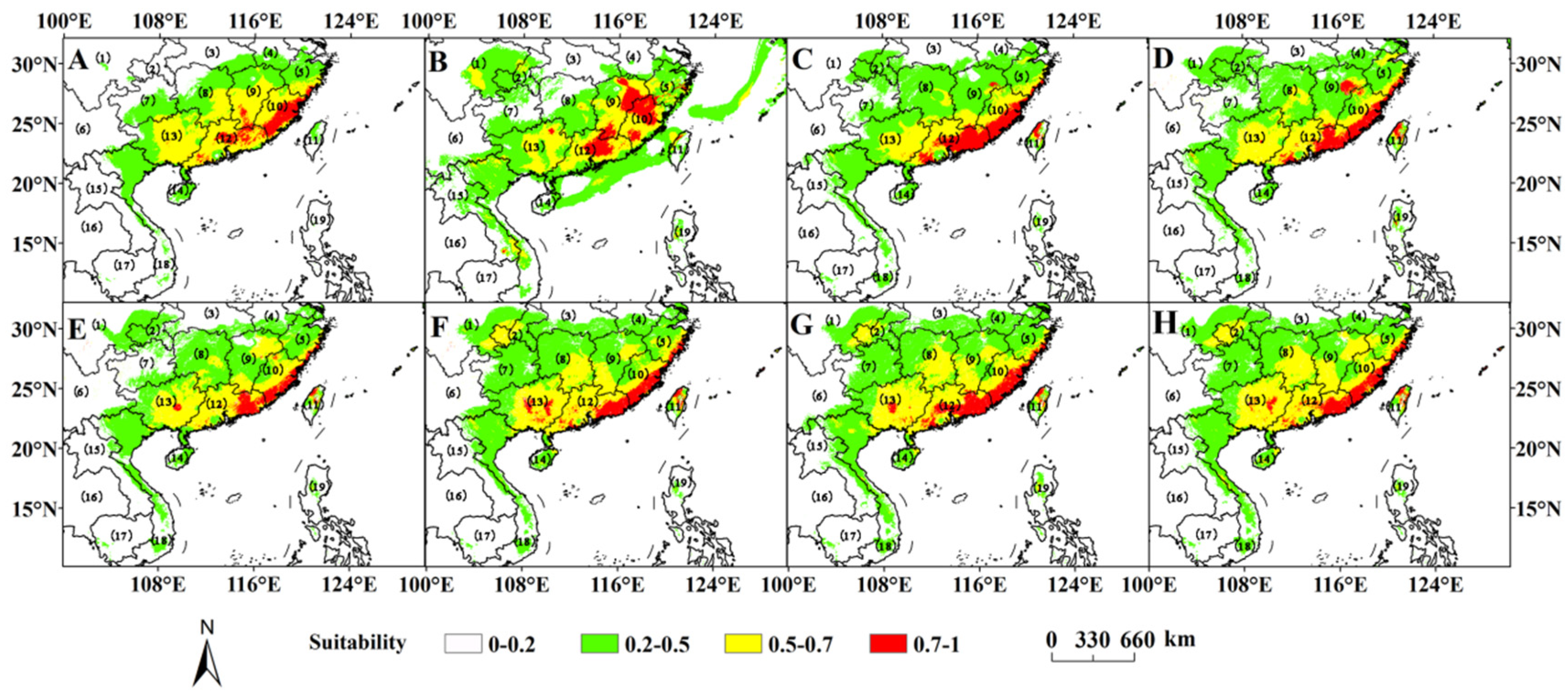
3.3. Current Potential Suitable Habitats
3.4. Potential Distribution in the Past and Future
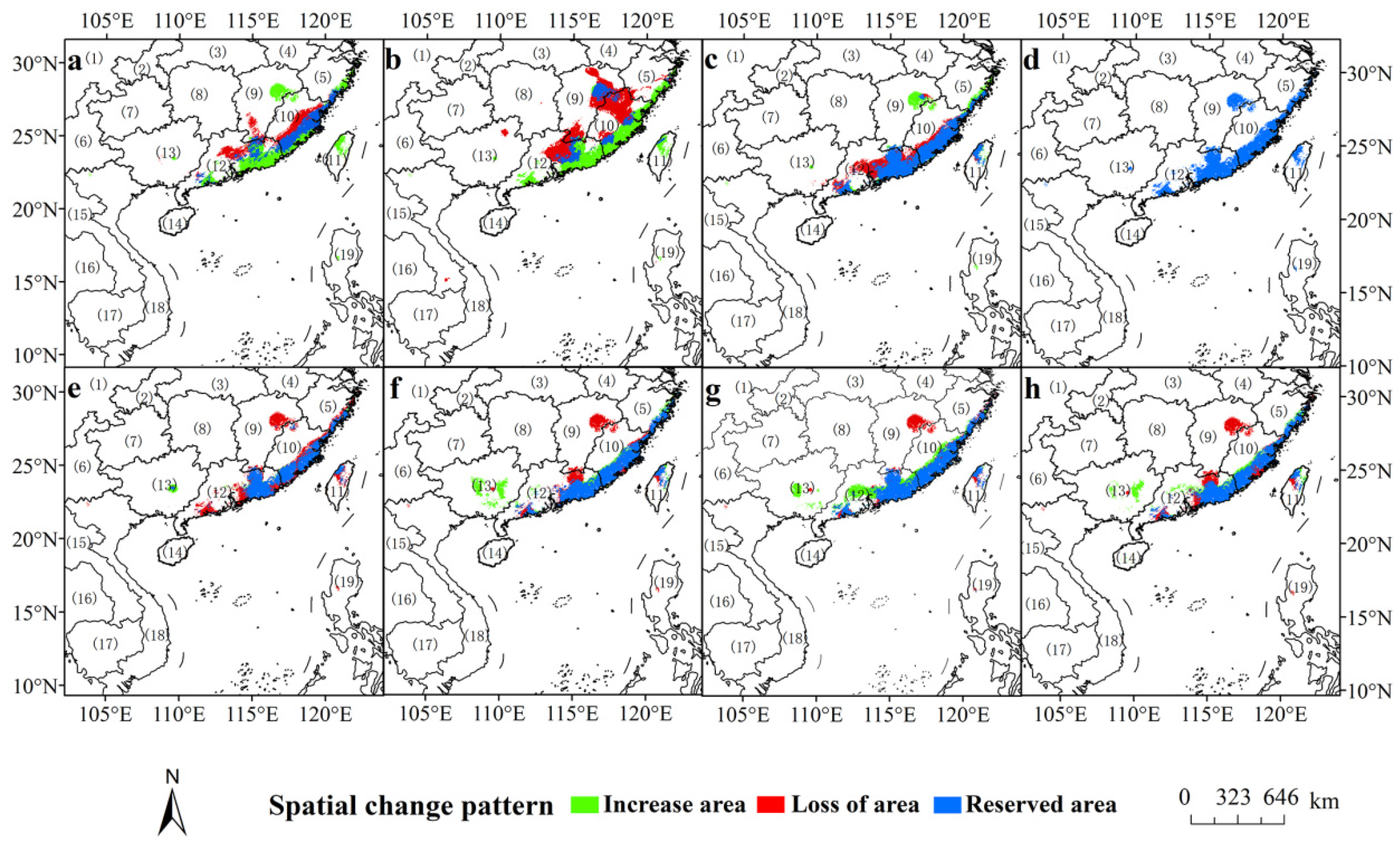
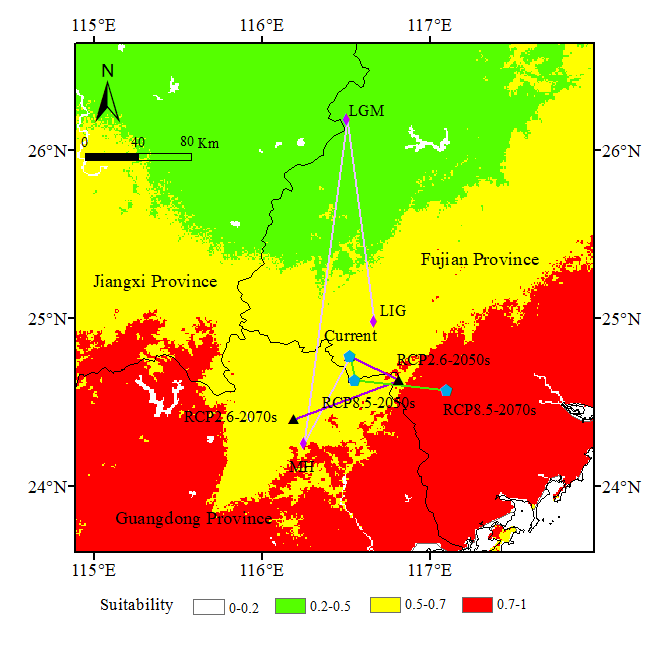
3.5. Shifts of the Core Suitable Habitat Distributions under Climate Change Scenarios
4. Discussion
4.1. The Change Pattern of Potential Distribution Area
4.2. Constraints of Climatic Factors on Geographical Distribution
4.3. Conservation Management
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Armenise, L.; Simeone, M.C.; Piredda, R.; Schirone, B. Validation of DNA barcoding as an efficient tool for taxon identification and detection of species diversity in Italian conifers. Eur. J. For. Res. 2012, 131, 1337–1353. [Google Scholar] [CrossRef]
- Crisp, M.D.; Cook, L.G. Cenozoic extinctions account for the low diversity of extant gymnosperms compared with angiosperms. New Phytol. 2011, 192, 997–1009. [Google Scholar] [CrossRef]
- Wu, F.; Dai, S.J.; Vlasak, J.; Spirin, V.; Dai, Y.C. Phylogeny and global diversity of Porodaedalea, a genus of gymnosperm pathogens in the Hymenochaetales. Mycologia 2019, 111, 40–53. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Liu, B.; Dennis, M.N. Red list assessment and conservation status of gymnosperms from China. Biodivers. Sci. 2017, 25, 758–764. [Google Scholar] [CrossRef] [Green Version]
- Paik, I.S.; Lee, Y.I.; Kim, H.J.; Huh, M. Time, space and structure on the korea cretaceous dinosaur coast: Cretaceous stratigraphy, geochronology, and paleoenvironments. Ichnos 2012, 19, 6–16. [Google Scholar] [CrossRef]
- Mantzouka, D.; Sakala, J.; Kvacek, Z.; Koskeridou, E.; Ioakim, C. Two fossil conifer species from the neogene of alonissos island (iliodroma, greece). Geodiversitas 2019, 41, 125–142. [Google Scholar] [CrossRef] [Green Version]
- Keller, G.; Mateo, P.; Monkenbusch, J.; Thibault, N.; Punekar, J.; Spangenberg, J.E.; Abramovich, S.; Ashckenazi-Polivoda, S.; Schoene, B.; Eddy, M.P.; et al. Mercury linked to deccan traps volcanism, climate change and the end-cretaceous mass extinction. Glob. Planet. Chang. 2020, 194, 17. [Google Scholar] [CrossRef]
- Eiserhardt, W.L.; Borchsenius, F.; Sandel, B.; Kissling, W.D.; Svenning, J.C. Late cenozoic climate and the phylogenetic structure of regional conifer floras world-wide. Glob. Ecol. Biogeogr. 2015, 24, 1136–1148. [Google Scholar] [CrossRef]
- Zheng, Y.; Liu, J.; Feng, X.Y.; Gong, X. The distribution, diversity, and conservation status of cycas in China. Ecol. Evol. 2017, 7, 3212–3224. [Google Scholar] [CrossRef]
- McCallum, M.L. Turtle biodiversity losses suggest coming sixth mass extinction. Biodivers. Conserv. 2021, 30, 1257–1275. [Google Scholar] [CrossRef]
- Elewa, A.M.T.; Abdelhady, A.A. Past, present, and future mass extinctions. J. Afr. Earth Sci. 2020, 162, 103678. [Google Scholar] [CrossRef]
- Barnosky, A.D.; Matzke, N.; Tomiya, S.; Wogan, G.O.U.; Swartz, B.; Quental, T.B.; Marshall, C.; McGuire, J.L.; Lindsey, E.L.; Maguire, K.C.; et al. Has the earth’s sixth mass extinction already arrived? Nature 2011, 471, 51–57. [Google Scholar] [CrossRef]
- Cursach, J.; Moragues, E.; Rita, J. The key role of accompanying species in the response of the critically endangered Naufraga balearica (Apiaceae) to climatic factors. Plant Ecol. 2018, 219, 561–576. [Google Scholar] [CrossRef]
- Qiu, C.J.; Shen, Z.H.; Peng, P.H.; Mao, L.F.; Zhang, X.S. How does contemporary climate versus climate change velocity affect endemic plant species richness in China? Chin. Sci. Bull. 2014, 59, 4660–4667. [Google Scholar] [CrossRef]
- Yang, Z.B.; Bai, Y.; Alatalo, J.M.; Huang, Z.D.; Yang, F.; Pu, X.Y.; Wang, R.B.; Yang, W.; Guo, X.Y. Spatio-temporal variation in potential habitats for rare and endangered plants and habitat conservation based on the maximum entropy model. Sci. Total Environ. 2021, 784, 13. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudik, M. Modeling of species distributions with maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Merow, C.; Smith, M.J.; Silander, J.A. A practical guide to maxent for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Garza, G.; Rivera, A.; Barrera, C.S.V.; Martinez-Avalos, J.G.; Dale, J.; Arroyo, T.P.F. Potential Effects of Climate Change on the Geographic Distribution of the Endangered Plant Species Manihot walkerae. Forests 2020, 11, 689. [Google Scholar] [CrossRef]
- Wu, Y.M.; Shen, X.L.; Tong, L.; Lei, F.W.; Mu, X.Y.; Zhang, Z.X. Impact of past and future climate change on the potential distribution of an endangered montane shrub Lonicera oblata and its conservation implications. Forests 2021, 12, 125. [Google Scholar] [CrossRef]
- Su, H.Y.; Bista, M.; Li, M.S. Mapping habitat suitability for Asiatic black bear and red panda in Makalu Barun National Park of Nepal from Maxent and GARP models. Sci. Rep. 2021, 11, 14135. [Google Scholar] [CrossRef]
- Ye, X.Z.; Zhao, G.H.; Zhang, M.Z.; Cui, X.Y.; Fan, H.H.; Liu, B. Distribution pattern of endangered plant Semiliquidambar cathayensis (Hamamelidaceae) in response to climate change after the last interglacial period. Forests 2020, 11, 125. [Google Scholar] [CrossRef]
- Mai, J.F.; Xian, Y.Y.; Liu, G.L. Predicting potential rainfall-triggered landslides sites in Guangdong Province (China) using MaxEnt model under climate changes scenarios. J. Geo-Inf. Sci. 2021, 23, 2042–2054. [Google Scholar] [CrossRef]
- Pandit, K.; Smith, J.; Quesada, T.; Villari, C.; Johnson, D.J. Association of Recent Incidence of Foliar Disease in Pine Species in the Southeastern United States with Tree and Climate Variables. Forests 2020, 11, 1155. [Google Scholar] [CrossRef]
- Ning, H.; Tang, M.; Chen, H. Impact of climate change on potential distribution of chinese white pine beetle dendroctonus armandi in China. Forests 2021, 12, 14. [Google Scholar] [CrossRef]
- Yang, X.H.; Jin, X.B.; Zhou, Y.K. Wildfire Risk Assessment and Zoning by Integrating Maxent and GIS in Hunan Province, China. Forests 2021, 12, 1299. [Google Scholar] [CrossRef]
- De Sousa, V.A.; Reeves, P.A.; Reilley, A.; de Aguiar, A.V.; Stefenon, V.M.; Richards, C.M. Genetic diversity and biogeographic determinants of population structure in Araucaria angustifolia (bert.) o. Ktze. Conserv. Genet. 2020, 21, 217–229. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y.D.; Li, X.Y.; Worth, J.R.P.; Lin, X.Y.; Ruhsam, M.; Chen, L.; Wu, X.T.; Wang, M.Q.; Thomas, P.I.; Wen, Y.F. Development of chloroplast microsatellite markers for Glyptostrobus pensilis (cupressaceae). Appl. Plant Sci. 2019, 7, 6. [Google Scholar] [CrossRef] [Green Version]
- Richards, K.; Mudie, P.; Rochon, A.; Athersuch, J.; Bolikhovskaya, N.; Hoogendoorn, R.; Verlinden, V. Late pleistocene to holocene evolution of the Emba Delta, Kazakhstan, and coastline of the north-eastern Caspian Sea: Sediment, ostracods, pollen and dinoflagellate cyst records. Palaeogeogr. Palaeocl. 2017, 468, 427–452. [Google Scholar] [CrossRef]
- Wu, X.; Ruhsam, M.; Wen, Y.; Thomas, P.; Worth, J.R.P.; Lin, X.; Wang, M.; Li, X.; Chen, L.; Lamxay, V.; et al. The last primary forests of the tertiary relict Glyptostrobus pensilis contain the highest genetic diversity. Forestry 2020, 93, 359–375. [Google Scholar] [CrossRef]
- Li, F.G.; Xia, N.H. The geographical distribution and cause of threat to Glyptostrobus pensilis (Taxodiaceae). J. Trop. Subtrop. Bot. 2004, 12, 13–20. [Google Scholar] [CrossRef]
- Averyanov, L.V.; Phan, K.L.; Nguyen, T.H.; Nguyen, S.K.; Nguyen, T.V.; Pham, T.D. Preliminary observation of native Glyptostrobus pensilis (Taxodiaceae) stands in Vietnam. Taiwania 2009, 54, 191–212. [Google Scholar] [CrossRef]
- Lepage, B.A. The taxonomy and biogeographic history of Glyptostrobus Endlicher (Cupressaceae). Bull. Peabody Mus. Nat. Hist. 2007, 48, 359–426. [Google Scholar] [CrossRef]
- Chen, Y.Q.; Wang, R.J.; Zhu, S.S.; Jiang, A.L.; Zhou, L.X. Population status and conservation strategy of the rare and endangered plant Glyptostrobus pensilis in Guangzhou. Trop. Geogr. 2016, 36, 944–951. [Google Scholar] [CrossRef]
- Tang, C.Q.; Yang, Y.C.; Momohara, A.; Wang, H.C.; Luu, H.T.; Li, S.F.; Song, K.; Qian, S.H.; LePage, B.; Dong, Y.F.; et al. Forest characteristics and population structure of Glyptostrobus pensilis, a globally endangered relict species of southeastern China. Plant Divers. 2019, 41, 237–249. [Google Scholar] [CrossRef]
- Ma, S.Y. Study on Protection and Breeding of Germplasm Resources and Application in Plant Landscaping of the Glyptostrobus pensilis; Zhejiang University: Hangzhou, China, 2020; pp. 1–43. [Google Scholar]
- Zhang, Y.M.; Yin, R.T.; Jia, R.R.; Yang, E.H.; Xu, H.M.; Tan, N.H. A new abietane diterpene from Glyptostrobus pensilis. Fitoterapia 2010, 81, 1202–1204. [Google Scholar] [CrossRef]
- Mao, D.H.; Wang, Z.M.; Wu, J.G.; Wu, B.F.; Zeng, Y.; Song, K.S.; Yi, K.P.; Luo, L. China’s wetlands loss to urban expansion. Land Degrad. Dev. 2018, 29, 2644–2657. [Google Scholar] [CrossRef]
- Dorken, V.M.; Rudall, P.J. Understanding the cone scale in Cupressaceae: Insights from seed-cone teratology in Glyptostrobus pensilis. Peer J. 2018, 6, e4948. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.Q.; Liu, J.F.; Wu, Z.Y.; Fu, D.L.; He, Z.S.; Dai, L.C.; Lu, J.C. The interspecific competition of main population in Glyptostrobus pensilis natural forest in Pingnan County. J. Fujian Coll. For. 2008, 28, 216–219. [Google Scholar] [CrossRef]
- Zheng, S.Q.; Wu, Z.Y.; Liu, J.F.; Hong, W.; He, Z.S.; Xu, D.W. The endangering causes and protection strategies for Glyptostrobus pensilis, an endemic relict plant in China. Subtrop. Agric. Res. 2011, 7, 217–220. [Google Scholar] [CrossRef]
- Muscarella, R.; Galante, P.J.; Soley-Guardia, M.; Boria, R.A.; Kass, J.M.; Uriarte, M.; Anderson, R.P. Enmeval: An r package for conducting spatially independent evaluations and estimating optimal model complexity for maxent ecological niche models. Methods Eco. Evol. 2014, 5, 1198–1205. [Google Scholar] [CrossRef]
- Wu, Z.Y. Study on Conservation Biology and Restoration Technique of the Relict Plant Glyptostrobus pensilis; Fujian Agriculture and Forestry University: Fuzhou, China, 2011; pp. 25–39. [Google Scholar]
- Loc, P.K.; The, P.V.; Long, P.K.; Regalado, J.; Averyanov, L.V.; Maslin, B. Native conifers of Vietnam—A review. Pak. J. Bot. 2017, 49, 2037–2068. [Google Scholar]
- Gent, P.R.; Danabasoglu, G.; Donner, L.J.; Holland, M.M.; Hunke, E.C.; Jayne, S.R.; Lawrence, D.M.; Neale, R.B.; Rasch, P.J.; Vertenstein, M.; et al. The community climate system model version 4. J. Clim. 2011, 24, 4973–4991. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Chao, B.X.; Dong, P.; Zhang, D.A.; Yu, W.W.; Hu, W.J.; Ma, Z.Y.; Chen, G.C.; Liu, Z.H.; Chen, B. Simulating spatial change of mangrove habitat under the impact of coastal land use: Coupling maxent and dyna-clue models. Sci. Total Environ. 2021, 788, 147914. [Google Scholar] [CrossRef] [PubMed]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Clim. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Shi, X.D.; Yin, Q.; Sang, Z.Y.; Zhu, Z.L.; Jia, Z.K.; Ma, L.Y. Prediction of potentially suitable areas for the introduction of Magnolia wufengensis under climate change. Ecol. Indic. 2021, 127, 14. [Google Scholar] [CrossRef]
- R Development Core Team. 2019. Available online: https://Cran.R-Project.Org/Manuals.Html (accessed on 5 January 2021).
- Yang, L.; Li, H.E.; Li, Q.; Guo, Q.Q.; Li, J.R. Genetic diversity analysis and potential distribution prediction of Sophora moorcroftiana endemic to Qinghai–Tibet Plateau, China. Forests 2021, 12, 1106. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Roberts, D.R.; Bahn, V.; Ciuti, S.; Boyce, M.S.; Elith, J.; Guillera-Arroita, G.; Hauenstein, S.; Lahoz-Monfort, J.J.; Schroder, B.; Thuiller, W.; et al. Cross-validation strategies for data with temporal, spatial, hierarchical, or phylogenetic structure. Ecography 2017, 40, 913–929. [Google Scholar] [CrossRef]
- Xu, Y.D.; Huang, Y.; Zhao, H.R.; Yang, M.L.; Zhuang, Y.Q.; Ye, X.P. Modelling the Effects of Climate Change on the Distribution of Endangered Cypripedium japonicum in China. Forests 2021, 12, 429. [Google Scholar] [CrossRef]
- Zhang, Y.P.; Liu, Y.L.; Qin, H.; Meng, Q.X. Prediction on spatial migration of suitable distribution of Elaeagnus mollis under climate change conditions in Shanxi Province, China. Chin. J. Appl. Ecol. 2019, 30, 496–502. [Google Scholar] [CrossRef]
- Wang, S.H. No Quaternary glacier ever existed in Fujian. J. Subtrop. Resour. Environ. 2008, 3, 83–88. [Google Scholar] [CrossRef]
- Liu, S.R.; Qin, C.F.; Peng, H. The discussion on the condition for glacier whether developed in Guangdong. Sci. Geogr. Sin. 2000, 20, 375–380. [Google Scholar]
- Kong, X.H. A floristic analysis on the Gymnosperms of Fujian Province. Plant Sci. J. 2004, 22, 514–522. [Google Scholar] [CrossRef]
- Kong, X.H. Studies on flora and geographical distribution of Gymnosperm in Wuyi Mountains, Fujian Province, China. J. Trop. Subtrop. Bot. 2011, 19, 33–39. [Google Scholar] [CrossRef]
- Yue, Y.; Zheng, Z.; Huang, K.; Chevalier, M.; Chase, B.M.; Carre, M.; Ledru, M.-P.; Cheddadi, R. A continuous record of vegetation and climate change over the past 50,000 years in the Fujian province of eastern subtropical China. Palaeogeogr. Palaeocl. 2012, 365, 115–123. [Google Scholar] [CrossRef]
- Zhao, Y.P.; Fan, G.Y.; Yin, P.P.; Sun, S.; Li, N.; Hong, X.N.; Hu, G.; Zhang, H.; Zhang, F.M.; Han, J.D.; et al. Resequencing 545 ginkgo genomes across the world reveals the evolutionary history of the living fossil. Nat. Commun. 2019, 10, 4201. [Google Scholar] [CrossRef] [Green Version]
- Xiao, J.Y.; Shang, Z.Y.; Shu, Q.; Yin, J.J.; Wu, X.S. The vegetation feature and palaeoenvironment significance in the mountainous interior of southern China from the last glacial maximum. Sci. China Earth Sci. 2018, 61, 71–81. [Google Scholar] [CrossRef]
- Li, Q.; Wu, H.; Yu, Y.; Sun, A.; Luo, Y. Quantifying regional vegetation changes in China during three contrasting temperature intervals since the Last Glacial Maximum. J. Asian Earth Sci. 2019, 174, 23–36. [Google Scholar] [CrossRef]
- Wang, M.Y.; Zheng, Z.; Man, M.L.; Hu, J.F.; Gao, Q.Z. Branched gdgt-based paleotemperature reconstruction of the last 30,000 years in humid monsoon region of southeast China. Chem. Geol. 2017, 463, 94–102. [Google Scholar] [CrossRef]
- Li, G.X.; Li, P.; Liu, Y.; Qiao, L.L.; Ma, Y.Y.; Xu, J.S.; Yang, Z.G. Sedimentary system response to the global sea level change in the East China Seas since the last glacial maximum. Earth-Sci. Rev. 2014, 139, 390–405. [Google Scholar] [CrossRef]
- Jiang, D.B.; Lang, X.M.; Tian, Z.P.; Guo, D.L. Last glacial maximum climate over China from PMIP simulations. Palaeogeogr. Palaeocl. 2011, 309, 347–357. [Google Scholar] [CrossRef]
- Xu, H.Y.; Chang, F.M.; Luo, Y.L.; Sun, X.J. Palaeoenvironmental changes from pollen record in deep sea core PC-1 from northern Okinawa Trough, East China Sea during the past 24 ka. Chin. Sci. Bull. 2009, 54, 3117–3126. [Google Scholar] [CrossRef]
- Zheng, Z.; Huang, K.Y.; Deng, Y.; Cao, L.L.; Yu, S.H.; Suc, J.P.; Berne, S.; Guichard, F. A–200 ka pollen record from Okinawa Trough: Paleoenvironment reconstruction of glacial-interglacial cycles. Sci. China Earth Sci. 2013, 43, 1231. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.T.; Lin, Z.S. Causes of submerged forests at Qianhu Bay, Zhangpu County, Fujian Province. Mar. Sci. Bull. 2009, 28, 84–89. [Google Scholar]
- Vickulin, S.V.; Ma, Q.W.; Zhilin, S.G.; Li, C.S. On cuticular compressions of Glyptostrobus europaeus (Taxodiaceae) from kaydagul formation (lower miocene) of the central kazakhstan. Acta Bot. Sin. 2003, 45, 673–680. [Google Scholar]
- Miller, C.N., Jr. Mesozoic conifers. Bot. Rev. 1977, 43, 217–280. [Google Scholar] [CrossRef]
- Yu, Y.F. Origin, evolution and distribution of the Taxodiacea. Acta Phytotaxon. Sin. 1995, 33, 362–389. [Google Scholar]
- Li, L.; Jin, J.H.; Manchester, S.R. Cupressaceae fossil remains from the paleocene of carneyville, wyoming. Rev. Palaeobot. Palynol. 2018, 251, 1–13. [Google Scholar] [CrossRef]
- Fauquette, S.; Suc, J.P.; Popescu, S.M.; Guillocheau, F.; Violette, S.; Jost, A.; Robin, C.; Briais, J.; Baby, G. Pliocene uplift of the Massif Central (France) constrained by the palaeoelevation quantified from the pollen record of sediments preserved along the Cantal Stratovolcano (Murat area). J.Geol. Soc. 2020, 177, 923–938. [Google Scholar] [CrossRef]
- Zheng, Z.; Peng, H.H.; Zheng, Y.W. Development and migration of Glyptostrobus pensilis and its rapid extinction in late Holocene. In Proceedings of the 27th Annual Conference of Palaeontological Society of China, Guilin, China, 1 November 2013; pp. 211–212. [Google Scholar]
- Sun, J.; Ma, C.M.; Cao, X.Y.; Zhao, Y.T.; Deng, Y.K.; Zhao, L.; Zhu, C. Quantitative precipitation reconstruction in the east-central monsoonal China since the late glacial period. Quatern. Int. 2019, 521, 175–184. [Google Scholar] [CrossRef]
- Tang, J.H. Holocene Profile Sediment of Tianhushan in Fujian and its Paleoclimate Significance; Fujian Normal University: Fuzhou, China, 2018; pp. 19–39. [Google Scholar]
- Peng, H.H.; Zheng, Z.; Zheng, Y.W.; Huang, K.Y.; Wei, J.H. Holocene vegetation changes and human activities revealed by a peat sediment core in Gaoyao, Zhaoqing. Quat. Sci. 2015, 35, 742–754. [Google Scholar] [CrossRef]
- Zheng, Z.; Ma, T.; Roberts, P.; Li, Z.; Yue, Y.F.; Peng, H.H.; Huang, K.Y.; Han, Z.Y.; Wan, Q.C.; Zhang, Y.Z.; et al. Anthropogenic impacts on Late Holocene land-coverchange and floristic biodiversity loss in tropical southeastern Asia. Proc. Natl. Acad. Sci. USA 2021, 118, e2022210118. [Google Scholar] [CrossRef]
- Cao, B.; Bai, C.; Xue, Y.; Yang, J.; Gao, P.; Liang, H.; Zhang, L.; Che, L.; Wang, J.; Xu, J.; et al. Wetlands rise and fall: Six endangered wetland species showed different patterns of habitat shift under future climate change. Sci. Total Environ. 2020, 731, 138518. [Google Scholar] [CrossRef] [PubMed]
- Qin, A.L.; Liu, B.; Guo, Q.S.; Bussmann, R.W.; Ma, F.Q.; Jian, Z.J.; Xu, G.X.; Pei, S.X. Maxent modeling for predicting impacts of climate change on the potential distribution of Thuja sutchuenensis Franch, an extremely endangered conifer from southwestern China. Glob. Ecol. Conserv. 2017, 10, 139–146. [Google Scholar] [CrossRef]
- Wu, C.Y.; Chen, D.S.; Shen, J.P.; Sun, X.M.; Zhang, S.G. Estimating the distribution and productivity characters of Larix kaempferi in response to climate change. J. Environ. Manag. 2021, 280, 111633. [Google Scholar] [CrossRef]
- Ding, Y.L.; Shi, Y.T.; Yang, S.H. Molecular regulation of plant responses to environmental temperatures. Mol. Plant 2020, 13, 544–564. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, T.; Kimura, S. Plant temperature sensors. Sensors 2018, 18, 4365. [Google Scholar] [CrossRef] [Green Version]
- Susila, H.; Nasim, Z.; Ahn, J.H. Ambient temperature-responsive mechanisms coordinate regulation of flowering time. Int. J. Mol. Sci. 2018, 19, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, G.B.; Liu, X.S.; Liang, W.B. Megasporogenesis, Female Gametophyte Development and Embryogenesis in Critically Endangered Glyptostrobus pensilis. Sci. Silvae Sin. 2015, 51, 50–62. [Google Scholar] [CrossRef]
- Huang, X. Study on Spatial Distribution Pattern and Suitability of the Original Metasequoia Glyptostroboides Population; Hubei Minzu University: Enshi, China, 2020; pp. 38–45. [Google Scholar]
- Wu, J.G. Risk and Uncertainty of Losing Suitable Habitat Areas Under Climate Change Scenarios: A Case Study for 109 Gymnosperm Species in China. Environ. Manag. 2020, 65, 517–533. [Google Scholar] [CrossRef]
- Cai, H.H.; Yangjin, Y.L.; Zhu, W.J.; Zhang, Z.L.; Wu, Y.B.; Zhuang, X.Y. Experiments of Effects of Water and Copper Stresses on the Seeding Growth of Glyptostrobus pensilis. J. Fujian For. Sci. Technol. 2011, 38, 46. [Google Scholar] [CrossRef]
- Wu, Z.Y.; Liu, J.F.; Hong, W.; Pan, D.M.; Zheng, S.Q. Genetic diversity of natural and planted Glyptostrobus pensilis populations: A comparative study. Chin. J. Appl. Ecol. 2011, 22, 873–879. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.Y. Development of EST–SSR Markers and Population Genetic Variation in Glyptostrobus pensilis; Central South University of Forestry and Technology: Changsha, China, 2018; pp. 33–62. [Google Scholar]
- Tam, N.M.; Duy, V.D.; Xuan, B.T.T.; Duc, N.M. Genetic variation and population structure in Chinese water pine (Glyptostrobus pensilis): A threatened species. Indian J. Biotechnol. 2013, 12, 499–503. [Google Scholar]
- Thomas, P.; LePage, B.A. The end of an era?—The conservation status of redwoods and other members of the former Taxodiaceae in the 21st century. Jpn. J. Histor. Bot. 2011, 19, 89–100. [Google Scholar]
- Li, G.; Xiao, N.W.; Luo, Z.L.; Liu, D.M.; Zhao, Z.P.; Guan, X.; Zang, C.X.; Li, J.S.; Shen, Z.H. Identifying conservation priority areas for Gymnosperm species under climate changes in China. Biol. Conserv. 2021, 253, 108914. [Google Scholar] [CrossRef]
- Chen, Y.Q.; Zhu, S.S.; Wang, G.T.; Wen, X.Y.; Huang, X.X.; Zhou, L.X.; Wang, R.J. Phylogenetic diversity analysis of the community of extremely small populations of Glyptostrobus pensilis. Plant Sci. J. 2017, 35, 667–678. [Google Scholar] [CrossRef]
- De Kort, H.; Prunier, J.G.; Ducatez, S.; Honnay, O.; Baguette, M.; Stevens, V.M.; Blanchet, S. Life history, climate and biogeography interactively affect worldwide genetic diversity of plant and animal populations. Nat. Commun. 2021, 12, 516. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environmental Variables | PC | PI(%) | TRGw | TRGo | TGw | TGo | AUCw | AUCo |
|---|---|---|---|---|---|---|---|---|
| Bio2 | 40.59 | 9.79 | 2.02 | 1.03 | 2.63 | 1.19 | 0.98 | 0.89 |
| Bio6 | 32.86 | 46.17 | 1.92 | 1.00 | 2.64 | 1.11 | 0.98 | 0.86 |
| Bio12 | 9.24 | 14.65 | 1.94 | 1.25 | 2.36 | 1.69 | 0.97 | 0.94 |
| Bio9 | 7.79 | 21.44 | 2.01 | 0.98 | 2.53 | 1.27 | 0.97 | 0.90 |
| Bio14 | 4.84 | 3.52 | 2.07 | 0.69 | 2.61 | 1.04 | 0.97 | 0.86 |
| Bio18 | 2.46 | 0.08 | 2.18 | 0.70 | 2.82 | 0.91 | 0.98 | 0.87 |
| Bio15 | 1.56 | 2.4 | 2.13 | 0.19 | 2.70 | 0.38 | 0.98 | 0.76 |
| Bio1 | 0.67 | 1.95 | 2.17 | 0.71 | 2.74 | 1.11 | 0.98 | 0.90 |
| Period | Highly Suitable Habitats | Moderately Suitable Habitats | Generally Suitable Habitats | Total |
|---|---|---|---|---|
| Last interglacial | 9.82 (−14.31) | 45.80 (34.07) | 62.72 (−35.76) | 118.34 (−17.69) |
| Last glacial maximum | 12.04 (5.06) | 35.88 (4.12) | 122.47 (25.43) | 170.39 (18.52) |
| Middle holocene | 12.73 (11.08) | 28.52 (−17.24) | 94.40 (−3.32) | 135.64 (−5.65) |
| Current | 11.46 | 34.46 | 97.64 | 143.77 |
| RCP2.6-2050s | 8.86 (−22.69) | 37.28 (8.18) | 96.72 (−0.94) | 142.86 (−0.63) |
| RCP8.5-2050s | 10.97 (−4.28) | 49.12 (42.54) | 107.83 (10.44) | 167.92 (16.80) |
| RCP2.6-2070s | 12.99 (13.35) | 49.14 (42.60) | 105.96 (8.52) | 168.09 (16.92) |
| RCP8.5-2070s | 10.32 (−9.95) | 55.33 (60.56) | 97.88 (0.25) | 163.53 (13.74) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, X.; Zhang, M.; Yang, Q.; Ye, L.; Liu, Y.; Zhang, G.; Chen, S.; Lai, W.; Wen, G.; Zheng, S.; et al. Prediction of Suitable Distribution of a Critically Endangered Plant Glyptostrobus pensilis. Forests 2022, 13, 257. https://doi.org/10.3390/f13020257
Ye X, Zhang M, Yang Q, Ye L, Liu Y, Zhang G, Chen S, Lai W, Wen G, Zheng S, et al. Prediction of Suitable Distribution of a Critically Endangered Plant Glyptostrobus pensilis. Forests. 2022; 13(2):257. https://doi.org/10.3390/f13020257
Chicago/Turabian StyleYe, Xingzhuang, Mingzhu Zhang, Qianyue Yang, Liqi Ye, Yipeng Liu, Guofang Zhang, Shipin Chen, Wenfeng Lai, Guowei Wen, Shiqun Zheng, and et al. 2022. "Prediction of Suitable Distribution of a Critically Endangered Plant Glyptostrobus pensilis" Forests 13, no. 2: 257. https://doi.org/10.3390/f13020257
APA StyleYe, X., Zhang, M., Yang, Q., Ye, L., Liu, Y., Zhang, G., Chen, S., Lai, W., Wen, G., Zheng, S., Ruan, S., Zhang, T., & Liu, B. (2022). Prediction of Suitable Distribution of a Critically Endangered Plant Glyptostrobus pensilis. Forests, 13(2), 257. https://doi.org/10.3390/f13020257