
Response of Functional Diversity of Soil Microbial Community to Forest Cutting and Regeneration Methodology in a Chinese Fir Plantation

 , ,
, ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
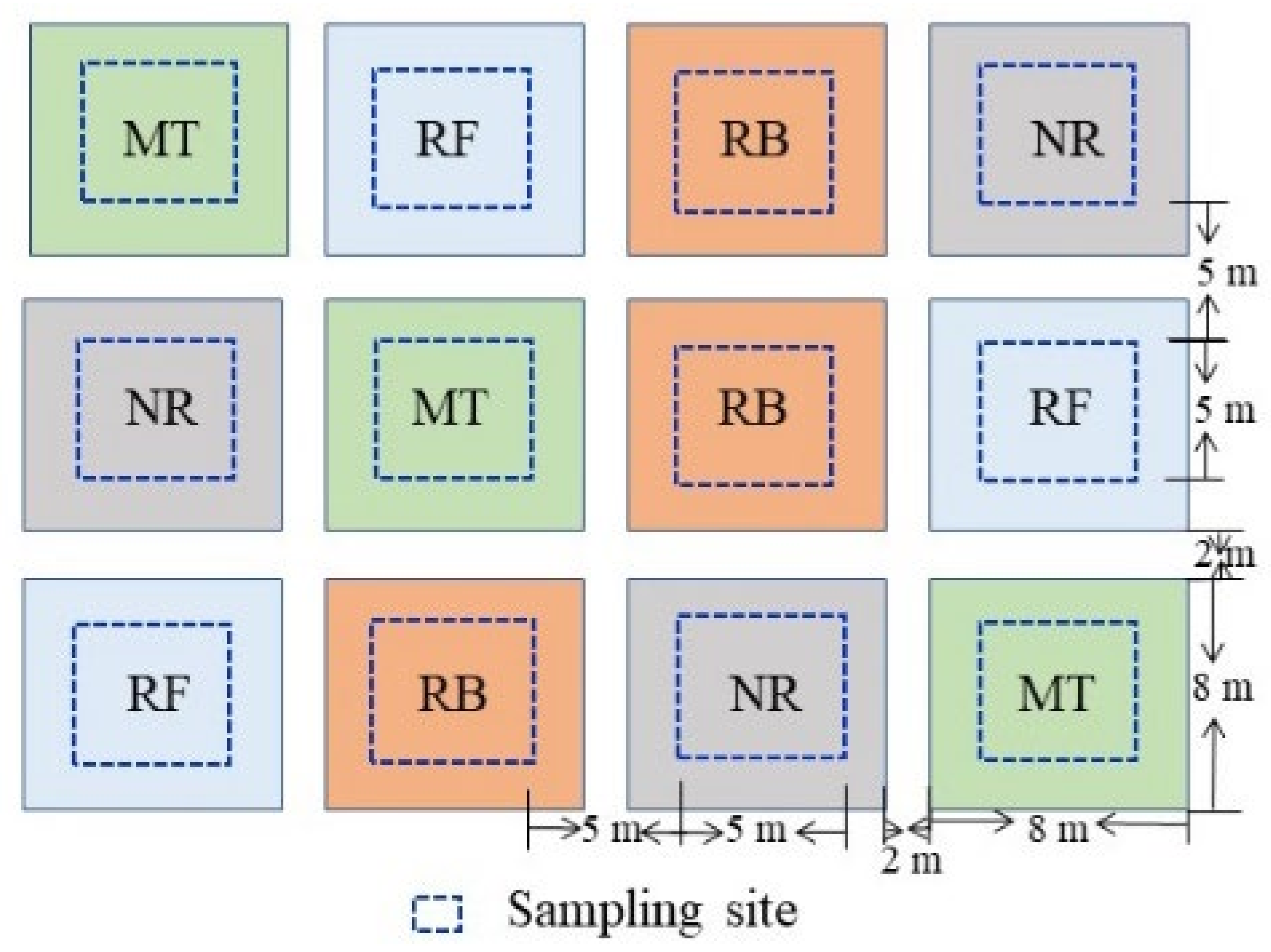
2.1. Study Areas
2.2. Treatments at the Experiment
2.3. Collection and Determination of Soil Samples
2.4. Data Analysis
3. Results
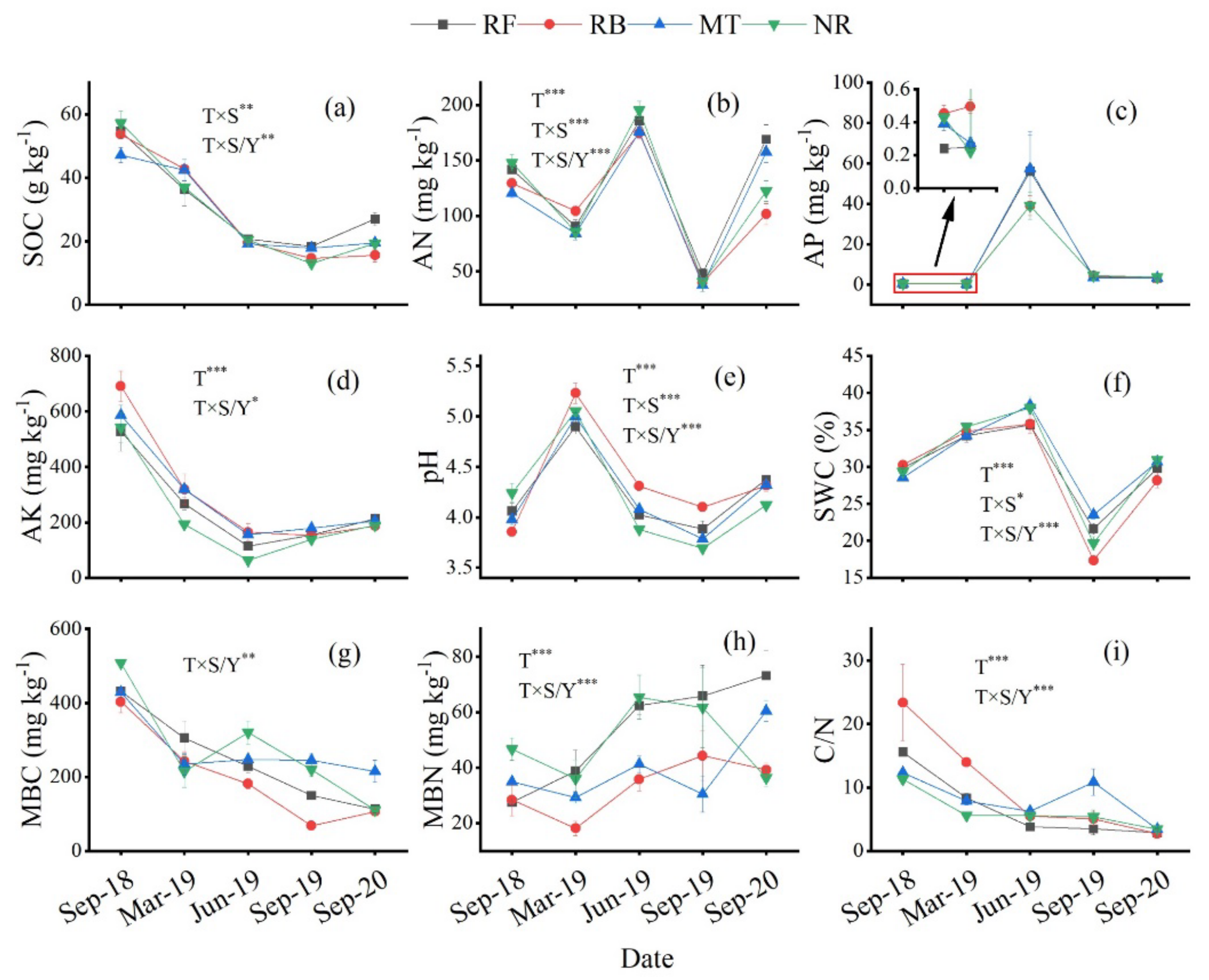
3.1. Dynamic Characteristics of Soil Physical and Chemical Properties
3.2. Dynamic Characteristics of Soil Physical and Chemical Properties
3.3. Dynamic Characteristics of Functional Diversity of Soil Microorganisms
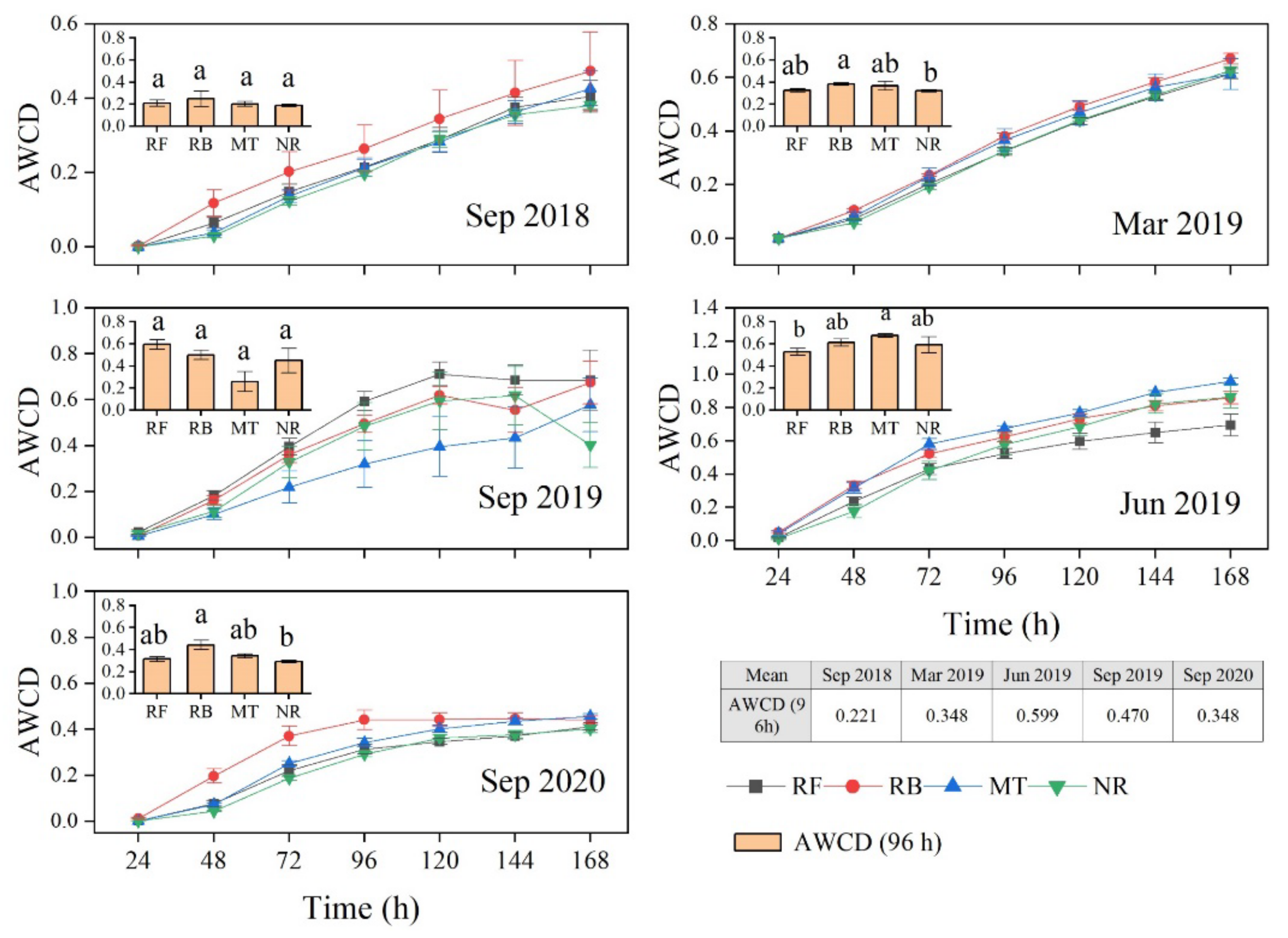
3.3.1. The Average Well Color Development
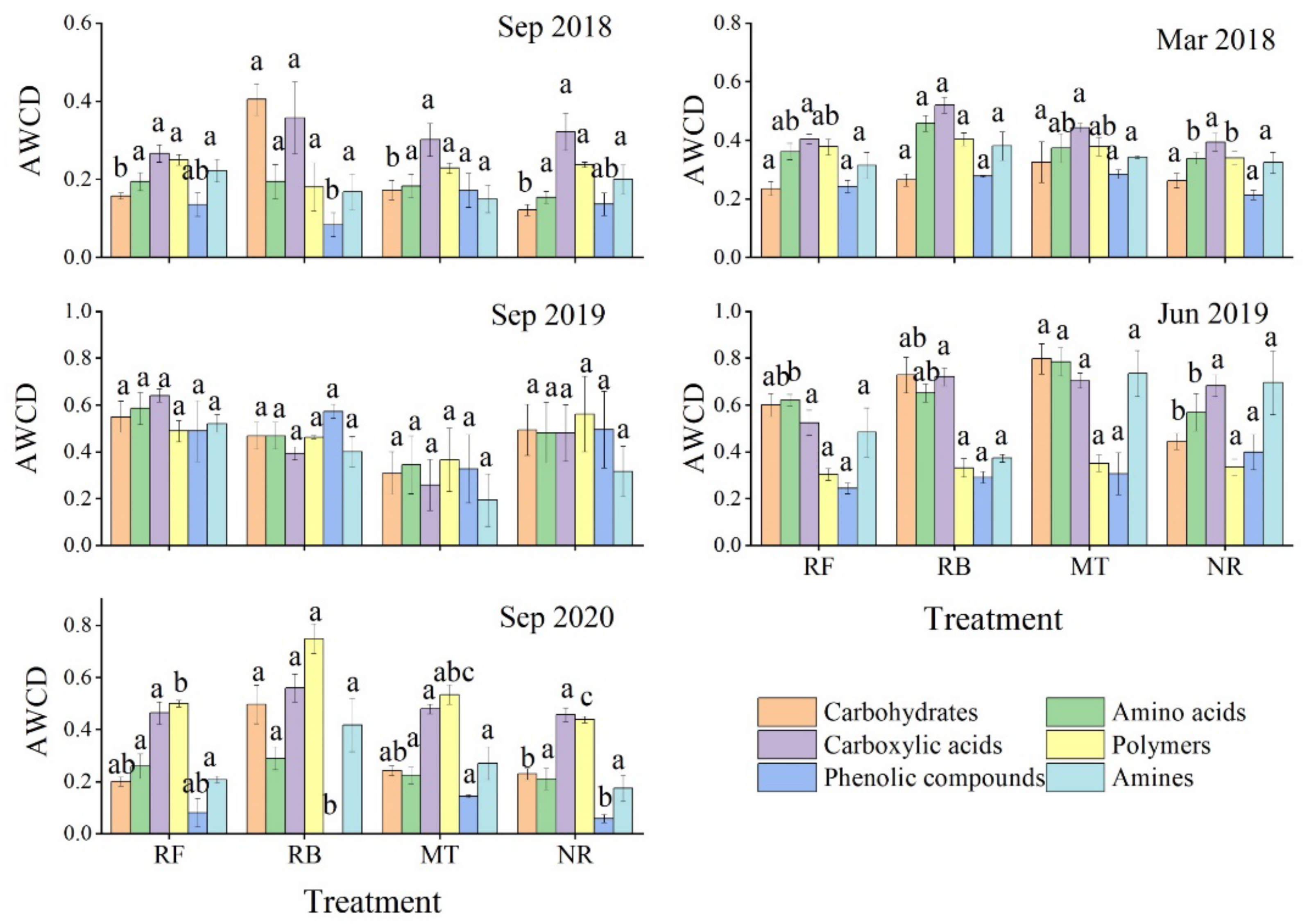
3.3.2. Utilization of Various Carbon Substrates by Soil Microbial Community
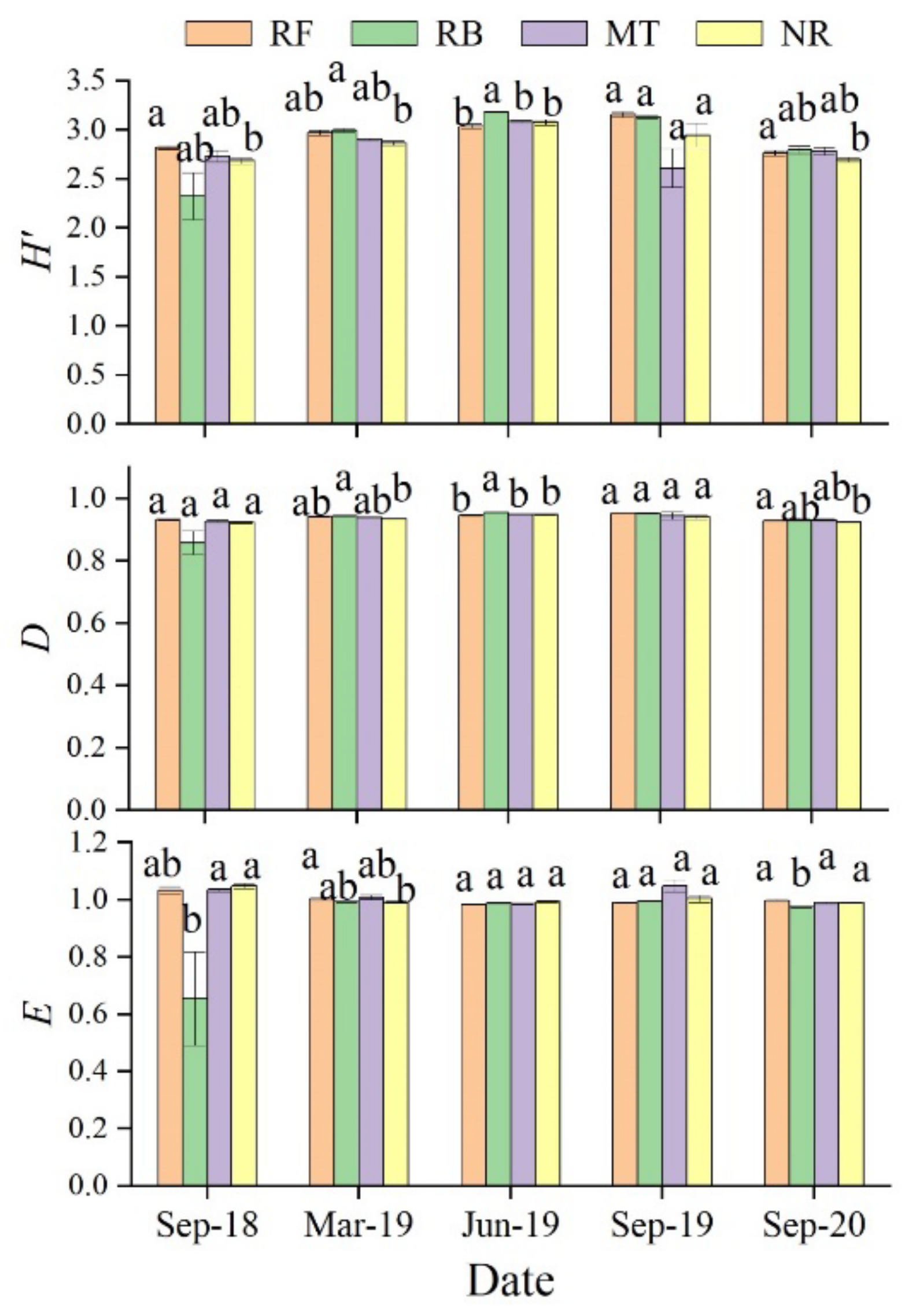
3.3.3. Diversity Index of Soil Microorganisms
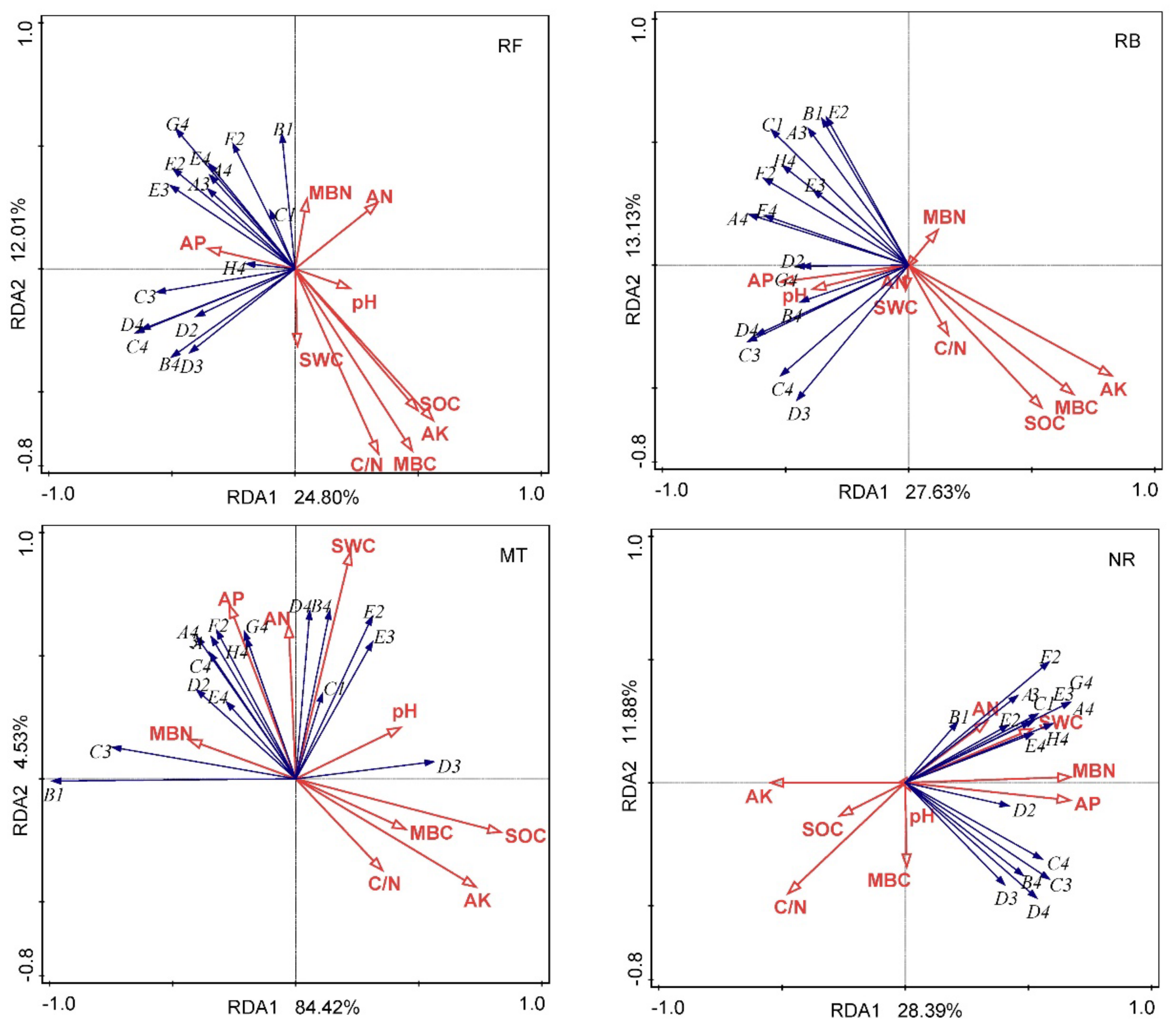
3.3.4. PCA of Carbon Utilization by Soil Microorganisms
3.4. Contribution of Soil Factors to Soil Microbial Community Functional Diversity Variations
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | T | % | S | % | S/Y | % | T × S | % | T × S/Y | % |
|---|---|---|---|---|---|---|---|---|---|---|
| Physicochemical index 1 | ||||||||||
| SOC | 1.91 | 0.46 | 100.75 *** | 16.10 | 420.61 *** | 67.20 | 3.56 ** | 1.70 | 3.68 ** | 1.76 |
| AN | 31.58 *** | 5.90 | 341.39 *** | 42.49 | 248.06 | 30.88 | 7.89 *** | 2.95 | 20.98 *** | 7.84 |
| AP | 0.51 | 0.43 | 91.34 *** | 51.89 | 0.42 | 0.24 | 1.17 | 1.99 | 0.00 | 0.01 |
| AK | 6.55 *** | 2.34 | 60.39 *** | 14.36 | 260.25 *** | 61.89 | 0.94 | 0.67 | 2.42 * | 1.72 |
| pH | 18.57 *** | 3.39 | 612.58 *** | 74.44 | 30.32 *** | 3.68 | 7.97 *** | 2.91 | 16.08 *** | 5.86 |
| SWC | 6.69 *** | 1.20 | 464.76 *** | 55.62 | 252.28 *** | 30.19 | 2.65 * | 0.95 | 6.87 *** | 2.47 |
| Microbial biomass 2 | ||||||||||
| MBC | 1.10 | 1.00 | 1.78 | 1.07 | 66.68 *** | 40.28 | 1.62 | 2.94 | 3.52 ** | 6.37 |
| MBN | 10.38 *** | 11.45 | 11.71 *** | 8.61 | 9.46 *** | 6.96 | 0.75 | 1.67 | 5.64 *** | 12.45 |
| MBC/MBN | 6.19 *** | 4.83 | 7.09 ** | 3.69 | 71.07 ** | 37.00 | 1.50 | 2.35 | 6.70 *** | 10.47 |
| Date | RF | RB | MT | NR |
|---|---|---|---|---|
| 18 September | 0.209 ± 0.027 | 0.248 ± 0.070 | 0.201 ± 0.022 | 0.188 ± 0.010 |
| 19 March | 0.325 ± 0.015 | 0.381 ± 0.012 | 0.367 ± 0.041 | 0.321 ± 0.010 |
| 19 June | 0.527 ± 0.034 | 0.612 ± 0.031 | 0.676 ± 0.015 | 0.591 ± 0.071 |
| 19 September | 0.594 ± 0.041 | 0.500 ± 0.043 | 0.258 ± 0.091 | 0.449 ± 0.111 |
| 20 September | 0.314 ± 0.022 | 0.441 ± 0.042 | 0.343 ± 0.018 | 0.292 ± 0.012 |
| Index | S | Y | ||||
|---|---|---|---|---|---|---|
| Spring | Summer | Fall | 0 | 1 | 2 | |
| AWCD | 0.348 ± 0.012 c | 0.599 ± 0.021 a | 0.470 ± 0.042 b | 0.221 ± 0.018 b | 0.470 ± 0.042 a | 0.348 ± 0.016 c |
| Carbohydrates | 0.272 ± 0.020 c | 0.649 ± 0.035 a | 0.456 ± 0.043 b | 0.196 ± 0.027 b | 0.456 ± 0.043 a | 0.276 ± 0.024 b |
| Phenolic compounds | 0.285 ± 0.016 a | 0.365 ± 0.037 a | 0.398 ± 0.058 a | 0.150 ± 0.017 b | 0.398 ± 0.058 a | 0.064 ± 0.014 b |
| Polymers | 0.375 ± 0.013 ab | 0.335 ± 0.017 b | 0.420 ± 0.042 a | 0.233 ± 0.020 c | 0.420 ± 0.042 b | 0.562 ± 0.025 a |
| Carboxylic acids | 0.422 ± 0.014 b | 0.659 ± 0.024 a | 0.482 ± 0.048 b | 0.305 ± 0.023 b | 0.482 ± 0.048 a | 0.491 ± 0.015 a |
| Amines | 0.356 ± 0.014 b | 0.598 ± 0.051 a | 0.383 ± 0.044 b | 0.186 ± 0.019 b | 0.383 ± 0.044 a | 0.262 ± 0.030 b |
| Amino acids | 0.383 ± 0.017 c | 0.687 ± 0.030 a | 0.493 ± 0.051 b | 0.181 ± 0.015 b | 0.493 ± 0.051 a | 0.246 ± 0.021 b |
| H’ | 2.934 ± 0.014 b | 3.090 ± 0.013 a | 2.961 ± 0.067 b | 2.632 ± 0.073 b | 2.961 ± 0.067 a | 2.760 ± 0.017 b |
| D | 0.940 ± 0.001 b | 0.949 ± 0.001 a | 0.947 ± 0.004 a | 0.909 ± 0.011 c | 0.947 ± 0.004 a | 0.928 ± 0.001 b |
| E | 0.999 ± 0.002 ab | 0.988 ± 0.001 b | 1.010 ± 0.007 a | 0.934 ± 0.053 a | 1.010 ± 0.007 a | 0.988 ± 0.002 a |
| Treatment | Variables | Explains % | Contribution % | Pseudo-F | p |
|---|---|---|---|---|---|
| RF | AK | 12.7 | 27.9 | 6.3 | 0.002 |
| pH | 8.7 | 19.1 | 5.0 | 0.004 | |
| MBC | 5.8 | 12.6 | 3.6 | 0.008 | |
| SWC | 5.1 | 11.2 | 2.7 | 0.012 | |
| AN | 4.4 | 9.6 | 2.2 | 0.028 | |
| MBN | 3.9 | 8.5 | 2.5 | 0.012 | |
| RB | AK | 21.5 | 43.3 | 11.8 | 0.002 |
| SOC | 5.9 | 11.8 | 3.4 | 0.006 | |
| AN | 7.7 | 15.4 | 5.0 | 0.002 | |
| MT | SOC | 58.8 | 64.6 | 61.4 | 0.002 |
| AP | 8.9 | 9.8 | 19.1 | 0.002 | |
| SWC | 7.6 | 8.3 | 9.5 | 0.002 | |
| MBN | 6.0 | 6.6 | 8.9 | 0.002 | |
| MBC | 6.0 | 6.6 | 18.3 | 0.002 | |
| AK | 2.0 | 2.2 | 7.1 | 0.002 | |
| AN | 0.8 | 0.9 | 3.1 | 0.03 | |
| NR | MBN | 13.4 | 25.9 | 6.7 | 0.002 |
| SWC | 10.6 | 20.5 | 5.9 | 0.004 | |
| MBC | 6.3 | 12.2 | 3.7 | 0.004 | |
| AP | 5.7 | 11.0 | 3.5 | 0.014 | |
| pH | 5.1 | 9.8 | 3.4 | 0.014 | |
| SOC | 4.5 | 8.7 | 3.3 | 0.008 |
References
- Engell, G.; Liejon, B. Effects of different levels of biomass removal in thinning on short-term production of Pinus sylvestris and Picea abies stands. Scan. J. For. Res. 1997, 12, 17–26. [Google Scholar]
- Bi, J.; Blanco, J.A.; Seely, B.; Kimmins, J.P.; Ding, Y.; Welham, C. Yield decline in Chinese-fir plantations: A simulation investigation with implications for model complexity. Can. J. For. Res. 2007, 37, 1615–1630. [Google Scholar] [CrossRef]
- Walmsley, J.D.; Jones, D.L.; Reynolds, B.; Price, M.H.; Healey, J.R. Whole tree harvesting can reduce second rotation forest productivity. For. Ecol. Manag. 2009, 257, 1104–1111. [Google Scholar] [CrossRef]
- Rocha, J.H.T.; Gonçalves, J.L.D.M.; Brandani, C.B.; Ferraz, A.D.V.; Franci, A.F.; Marques, E.R.G.; Junior, J.C.A.; Hubner, A. Forest residue removal decreases soil quality and affects wood productivity even with high rates of fertilizer application. For. Ecol. Manag. 2018, 430, 188–195. [Google Scholar] [CrossRef]
- Naeem, S.; Li, S. Biodiversity enhances ecosystem reliability. Nature 1997, 390, 507–509. [Google Scholar] [CrossRef]
- Fuhrman, J.A. Microbial community structure and its functional implications. Nature 2009, 459, 193–199. [Google Scholar] [CrossRef]
- Colombo, F.; Macdonald, C.A.; Jeffries, T.C.; Powell, J.R.; Singh, B.K. Impact of forest management practices on soil bacterial diversity and consequences for soil processes. Soil Biol. Biochem. 2016, 94, 200–210. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Grinyer, J.; Reich, P.; Singh, B. Relative importance of soil properties and microbial community for soil functionality: Insights from a microbial swap experiment. Funct. Ecol. 2016, 30, 1862–1873. [Google Scholar] [CrossRef]
- Sparling, G.P. Soil Microbial Biomass, Activity and Nutrient Cycling as Indicators of Soil Health. In Biological Indicators of Soil Health; Pankhurst, C.E., Doube, B.M., Gupta, V.V.S.R., Eds.; CAB International: Wallingford, CT, USA, 1997; pp. 97–119. [Google Scholar]
- Anderson, T.H. Microbial eco-physiological indicators to asses soil quality. Agric. Ecosyst. Environ. 2003, 98, 285–293. [Google Scholar] [CrossRef]
- Jesus, E.C.; Marsh, T.L.; Tiedje, J.M.; De, S.J. Changes in land use alter the structure of bacterial communities in Western Amazon soils. ISME J. 2009, 3, 1004–1011. [Google Scholar] [CrossRef]
- Garau, G.; Morillas, L.; Roales, J.; Castaldi, P.; Mangia, N.P.; Spano, D.; Mereu, S. Effect of monospecific and mixed Mediterranean tree plantations on soil microbial community and biochemical functioning. Appl. Soil Ecol. 2019, 140, 78–88. [Google Scholar] [CrossRef]
- Berkelmann, D.; Schneider, D.; Meryandini, A.; Daniel, R. Unravelling the effects of tropical land use conversion on the soil microbiome. Environ. Microbiome 2020, 15, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mushinski, R.M.; Zhou, Y.; Gentry, T.J.; Boutton, T.W. Bacterial metataxonomic profile and putative functional behavior associated with C and N cycle processes remain altered for decades after forest harvest. Soil Biol. Biochem. 2018, 119, 184–193. [Google Scholar] [CrossRef]
- Rutgers, M.; Wouterse, M.; Drost, S.M.; Breure, A.M.; Mulder, C.; Stone, D.; Bloem, J. Monitoring soil bacteria with community-level physiological profiles using Biolog™ ECO-plates in the Netherlands and Europe. Appl. Soil Ecol. 2016, 97, 23–35. [Google Scholar] [CrossRef]
- Selvaraj, S.; De la Rosa, J.M.; Zhijun, H.; Guo, F.; Ma, X. Effects of ageing and successive slash-and-burn practice on the chemical composition of charcoal and yields of stable carbon. Catena 2018, 162, 141–147. [Google Scholar]
- Grier, C.C.; Cole, D.W. Influence of slash burning on ion transport in a forest soil. Northwest Sci. 1971, 45, 100–106. [Google Scholar]
- Prats, S.A.; Abrantes, J.R.; Crema, I.P.; Keizer, J.J.; de Lima, J.L. Runoff and soil erosion mitigation with sieved forest residue mulch strips under controlled laboratory conditions. For. Ecol. Manag. 2017, 396, 102–112. [Google Scholar] [CrossRef]
- DeBano, L.F.; Neary, D.G.; Ffolliot, P.F. Fire’s Effects on Ecosystems, 1st ed.; Wiley: New York, NY, USA, 1998. [Google Scholar]
- Glenn, N.F.; Finley, C.D. Fire and vegetation type effects on soil hydrophobicity and infiltration in the sagebrush-steppe steppe: I. Field analysis. J. Arid. Environ. 2010, 74, 653–659. [Google Scholar] [CrossRef]
- Fultz, L.M.; Moore-Kucera, J.; Dathe, J.; Davinic, M.; Perry, G.; Wester, D.; Rideout-Hanzak, S. Forest wildfire and grassland prescribed fire effects on soil biogeochemical processes and microbial communities: Two case studies in the semi-arid Southwest. Appl. Soil Ecol. 2016, 99, 118–128. [Google Scholar] [CrossRef]
- Bååth, E.; Frostegård, Å.; Pennanen, E.; Fritze, H. Microbial community structure and pH response in relation to soil organic matter quality in wood-ash fertilized, clear-cut or burned coniferous forest soils. Soil Biol. Biochem. 1995, 27, 229–240. [Google Scholar] [CrossRef] [Green Version]
- Suominen, M.; Junninen, K.; Heikkala, O.; Kouki, J. Burning harvested sites enhances polypore diversity on stumps and slash. For. Ecol. Manag. 2018, 414, 47–53. [Google Scholar] [CrossRef]
- Pietikäinen, J.; Fritze, H. Clear-cutting and prescribed burning in coniferous forest: Comparison of efects on soil fungal and total microbial biomass, respiration activity and nitrification. Soil Biol. Biochem. 1995, 27, 101–109. [Google Scholar] [CrossRef]
- Bautista, S.; Robichaud, P.R.; Bladé, C. Post-fire mulching. In Fire Effects on Soils and Restoration Strategies; Cerdá, A., Robichaud, P., Eds.; Science Publishers: Enfield, NH, USA, 2009; pp. 353–372. [Google Scholar]
- Díaz-Raviña, M.; Martín, M.; Barreiro, A.; Lombao, A.; Iglesias, L.; Díaz-Fierros, F.; Carballas, T. Mulching and seeding treatments for post-fire soil stabilisation in NW Spain: Short-term effects and effectiveness. Geoderma 2012, 191, 31–39. [Google Scholar] [CrossRef]
- Suominen, M.; Junninen, K.; Kouki, J. Diversity of fungi in harvested forests 10 years after logging and burning: Polypore assemblages on different woody substrates. For. Ecol. Manag. 2019, 446, 63–70. [Google Scholar] [CrossRef]
- Savill, P.; Eanns, J.; Auclair, D. Plantation Silviculture in Europe; University Press: Oxford, UK, 1997. [Google Scholar]
- Liu, B.; Daryanto, S.; Wang, L.; Li, Y.; Liu, Q.; Zhao, C.; Wang, Z. Excessive Accumulation of Chinese Fir Litter Inhibits Its Own Seedling Emergence and Early Growth—A Greenhouse Perspective. Forests 2017, 8, 341. [Google Scholar] [CrossRef] [Green Version]
- Premer, M.I.; Froese, R.E.; Vance, E.D. Whole-tree harvest and residue recovery in commercial aspen: Implications to forest growth and soil productivity across a rotation. For. Ecol. Manag. 2019, 447, 130–138. [Google Scholar] [CrossRef]
- Muñoz Delgado, B.L.; Kenefic, L.S.; Weiskittel, A.R.; Fernandez, I.J.; Benjamin, J.G.; Dibble, A.C. Northern mixed wood composition and productivity 50 years after whole-tree and stem-only harvesting with and without post-harvest prescribed burning. For. Ecol. Manag. 2019, 441, 155–166. [Google Scholar] [CrossRef]
- Tew, D.T.; Morris, L.A.; Allen, H.L.; Wells, C.G. Estimates of nutrient removal, displacement and loss resulting from harvest and site preparation of a Pinus taeda plantation in the piedmont of North Carolina. For. Ecol. Manag. 1986, 15, 257–267. [Google Scholar]
- Mueller, T.; Jensen, L.S.; Nielsen, N.E.; Magid, J. Turnover of carbon and nitrogen in a sandy loam soil following incorporation of chopped maize plants, barley straw and blue grass in the field. Soil Biol. Biochem. 1998, 30, 561–571. [Google Scholar] [CrossRef]
- Chevanan, N.; Womac, A.R.; Bitra, V.; Igathinathane, C.; Sokhansanj, S. Bulk density and compaction behavior of knife mill chopped switchgrass, wheat straw, and corn stover. Bioresour. Technol. 2009, 101, 207–214. [Google Scholar] [CrossRef]
- Sinkeviiene, A.; Jodaugiene, D.; Pupaliene, R.; Urboniene, M. The influence of organic mulches on soil properties and crop yield. Agron. Res. 2009, 7, 485–491. [Google Scholar]
- Palmieri, N.; Forleo, M.B.; Giannoccaro, G.; Suardi, A. Environmental impact of cereal straw management: An on-farm assessment. J. Clean. Prod. 2017, 142, 2950–2964. [Google Scholar] [CrossRef]
- Peng, Z. Rapid Growth and High Yield Cultivation of Chinese Fir; Anhui Science and Technology Press: Hefei, China, 1984; pp. 3–4. [Google Scholar]
- FAO. Global Forests Resources Assessment 2010-Main report. FAO For. Pap. 2010, 163, 44–48. [Google Scholar]
- Wu, Z. Chinese Fir; China Forestry Publishing House: Beijing, China, 1984; pp. 8–20. (In Chinese) [Google Scholar]
- Sheng, W. On the maintenance of long-term productivity of plantation in China. For. Res. 2018, 31, 1–14. [Google Scholar]
- Bao, S.D. Soil Agrochemical Analysis, 3rd ed.; China Agriculture Press: Beijing, China, 2000. [Google Scholar]
- Brookes, P.C.; Landman, A.; Pruden, G.; Jenkinson, D.S. Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biol. Biochem. 1985, 17, 837–842. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Joergensen, R.G.; Mueller, T. The fumigation-extraction method to estimate soil microbial biomass: Calibration of the kEN value. Soil Biol. Biochem. 1996, 28, 33–37. [Google Scholar] [CrossRef]
- Classen, A.T.; Boyle, S.I.; Haskins, K.E.; Overby, S.T.; Hart, S.C. Community-level physiological profiles of bacteria and fungi: Plate type and incubation temperature influences on contrasting soils. FEMS. Microbiol. Ecol. 2003, 44, 319–328. [Google Scholar] [CrossRef] [Green Version]
- Garland, J.L.; Mills, A.L. Classification and characterization of heterotrophic microbial communities on the basis of patterns of community-level sole-carbon-source utilization. Appl. Environ. Microb. 1991, 57, 2351–2359. [Google Scholar] [CrossRef] [Green Version]
- Insam, H. A New Set of Substrates Proposed for Community Characterization in Environmental Samples. In Microbial Communities; Insam, H., Rangger, A., Eds.; Springer: Berlin, Germany, 1997; pp. 259–260. [Google Scholar]
- Zak, J.C.; Willig, M.R.; Moorhead, D.L.; Wildman, H.G. Functional diversity of microbial communities: A quantitative approach. Soil Biol. Biochem. 1994, 26, 1101–1108. [Google Scholar] [CrossRef]
- Magguran, A.E. Ecological Diversity and Its Measurement; Princeton University Press: Princeton, NJ, USA, 1988; pp. 141–162. [Google Scholar]
- Fontúrbel, M.T.; Barreiro, A.; Vega, J.A.; Martin, A.; Jiménez, E.; Carballas, T.; Fernández, C.; Diaz-Ravina, M. Effects of an experimental fire and post-fire stabilization treatments on soil microbial communities. Geoderma 2012, 191, 51–60. [Google Scholar] [CrossRef]
- Ma, S.; Concilio, A.; Oakley, B.; North, M.; Chen, J. Spatial variability in microclimate in a mixed-conifer forest before and after thinning and burning treatments. For. Ecol. Manag. 2010, 259, 904–915. [Google Scholar] [CrossRef]
- Chen, J.; Franklin, J.F.; Spies, T.A. Contrasting microclimates among clearcut, edge, and interior of old-growth Douglas-fir forest. Agric. For. Meteorol. 1993, 63, 219–237. [Google Scholar] [CrossRef]
- D’Ascoli, R.; Rutigliano, F.A.; De Pascale, R.A.; Gentile, A.; De Santo, V.A. Functional diversity of the microbial community in Mediterranean maquis soils as affected by fires. Int. J. Wildland Fire 2005, 14, 355–363. [Google Scholar] [CrossRef]
- Neary, D.G.; Klopatek, C.C.; DeBano, L.F.; Ffolliott, P.F. Fire effects on belowground sustainability: A review and synthesis. For. Ecol. Manag. 1999, 122, 70–71. [Google Scholar] [CrossRef]
- Rocha, J.H.T.; Gonçalves, J.L.D.M.; Gava, J.L.; Godinho, T.D.O.; Melo, E.A.S.C.; Bazani, J.H.; Wichert, M.P. Forest residue maintenance increased the wood productivity of a Eucalyptus plantation over two short rotations. For. Ecol. Manag. 2016, 379, 1–10. [Google Scholar] [CrossRef]
- Staddon, W.J.; Duchesne, L.C.; Trevors, J.T. Impact of clear-cutting and prescribed burning on microbial diversity and community structure in a Jack pine (Pinus banksiana Lamb.) clear-cut using Biolog Gram-negative microplates. World J. Microbiol. Biotechnol. 1997, 14, 119–123. [Google Scholar] [CrossRef]
- Yang, Y.; Guo, J.; Chen, G.; He, Z.; Xie, J. Effect of Slash Burning on Nutrient Removal and Soil Fertility in Chinese Fir and Evergreen Broadleaved Forests of Mid-Subtropical China. Pedosphere 2003, 13, 89–96. [Google Scholar]
- Borchers, S.L.; Perry, D.A. Growth and ectomycorrhiza formation of douglas-fir seedlings grown in soils collected at different distances from pioneering hardwoods in southwest Oregon clear-cuts. Can. J. For. Res. 1990, 20, 712–721. [Google Scholar] [CrossRef]
- Certini, G. Effects of fire on properties of forest soils: A review. Oecologia 2005, 143, 1–10. [Google Scholar] [CrossRef]
- Alcañiz, M.; Outeiro, L.; Francos, M.; Úbeda, X. Effects of prescribed fires on soil properties: A review. Sci. Total Environ. 2018, 613–614, 944–957. [Google Scholar] [CrossRef] [PubMed]
- Blankenship, B.A.; Arthur, M.A. Soil Nutrient and Microbial Response to Prescribed Fire in an Oak-Pine Ecosystem in Eastern Kentucky. In Proceedings of the 12th Central Hardwood Forest Conference, USDA For. Serv. Gen. Tech. Rep. SRS-24. Southern Research Station, Asheville, NC, USA, 28 March 1999; pp. 39–47. [Google Scholar]
- Gray, D.M.; Dighton, J. Nutrient utilization by pine seedlings and soilmicrobes in oligotrophic pine barrens forest soils subjected to prescribed fire treatment. Soil Biol. Biochem. 2009, 41, 1957–1965. [Google Scholar] [CrossRef]
- Marschner, P.; Kandeler, E.; Marschner, B. Structure and function of the soil microbial community in a long-term fertilizer experiment. Soil Biol. Biochem. 2003, 35, 453–461. [Google Scholar] [CrossRef]
- Carney, K.M.; Matson, P.A. Plant communities, soil microorganisms, and soil carbon cycling: Does altering the world belowground matter to ecosystem functioning? Ecosystems 2005, 8, 928–940. [Google Scholar] [CrossRef]
- Chen, X.; Wang, D.; Chen, X.; Wang, J.; Diao, J.; Zhang, J.; Guan, Q. Soil microbial functional diversity and biomass as affected by different thinning intensities in a Chinese fir plantation. Appl. Soil Ecol. 2015, 92, 35–44. [Google Scholar] [CrossRef]
- Wang, Z.; Pan, L.; Xue, L. Effects of forest gaps on soil microbial diversity in a Cunninghamia lanceolata Stand after a Severe Ice Storm. Int. J. Agric. Biol. 2018, 20, 1161–1166. [Google Scholar]
- Wang, Q.; Wang, S.; Fan, B.; Yu, X. Litter production, leaf litter decomposition and nutrient return in Cunninghamia lanceolata plantations in south China: Effect of planting conifers with broadleaved species. Plant Soil 2007, 297, 201–211. [Google Scholar] [CrossRef]
- Zhou, L.; Shalom, A.-D.D.; Wu, P.; Li, S.; Jia, Y.; Ma, X. Litterfall production and nutrient return in different-aged Chinese fir (Cunninghamia lanceolata) plantations in South China. J. For. Res. 2015, 26, 79–89. [Google Scholar] [CrossRef]
- Rietl, A.J.; Jackson, C.R. Effects of the ecological restoration practices of prescribed burning and mechanical thinning on soil microbial enzyme activities and leaf litter decomposition. Soil Biol. Biochem. 2012, 50, 47–57. [Google Scholar] [CrossRef]
- Huang, Z.; Xu, Z.; Chen, C. Effect of mulching on labile soil organic matter pools, microbial community functional diversity and nitrogen transformations in two hardwood plantations of subtropical Australia. Appl. Soil Ecol. 2008, 40, 229–239. [Google Scholar] [CrossRef]
- Chen, J.; Saunders, S.C.; Crow, T.R.; Naiman, R.J.; Brosofske, K.D.; Mroz, G.D.; Brookshire, B.L.; Franklin, J.F. Microclimate in forest ecosystem and landscape ecology: Variations in local climate can be used to monitor and compare the effects of different management regimes. BioScience 1999, 49, 288–297. [Google Scholar] [CrossRef] [Green Version]
- Athy, E.R.; Keiffer, C.H.; Stevens, M.H. Effects of mulch on seedlings and soil on a closed landfill. Restor. Ecol. 2006, 14, 233–241. [Google Scholar] [CrossRef]
- DeDorest, J.L.; Chen, J.; Mcnulty, S.G. Leaf litter is an important mediator of soil respiration in an oak-dominated forest. Int. J. Biometeorol. 2009, 53, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Fernández, C.; Vega, J.A.; Jiménez, E.; Fonturbel, T. Effectiveness of three postfire treatments at reducing soil erosion in Galicia (NW Spain). Int. J. Wildland Fire 2011, 20, 104–114. [Google Scholar] [CrossRef] [Green Version]
- Fernández, C.; Vega, J.A.; Jiménez, E.; Vieira, D.C.S.; Merino, A.; Ferreiro, A.; Fonturbel, T. Seeding and mulching plus seeding effects on post-fire runoff, soil erosion and species diversity in Galicia (NW Spain). Land Degrad. Dev. 2012, 23, 150–156. [Google Scholar] [CrossRef]
- Chantigny, M.H. Dissolved and water-extractable organic matter in soils: A review on the influence of land use and management practices. Geoderma 2003, 113, 357–380. [Google Scholar] [CrossRef]
- Sǿnsteby, A.; Nes, A.; Måge, F. Effects of bark mulch and NPK fertilizer on yield, leaf nutrien status and soil mineral nitrogen during three years of strawberry production. Acta Agric. Scand. 2004, 54, 128–134. [Google Scholar]
- Fukuoka, M. The One-Straw Revolution. In An Introduction to Natural Farming; Pearce, C., Kurosawa, T., Korn, L., Eds.; Originally published as Shizen Noho Wara Ippon No Kakumei; Hakujusha Co., Ltd.: Tokyo, Japan, 1978. [Google Scholar]
- Kesavan, P.C.; Swaminathan, M.S. Strategies andmodels for agricultural sustainability in developing Asian countries. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2008, 363, 877–891. [Google Scholar] [CrossRef]
- Budelman, A. The decomposition of the leaf mulches of Leucaena leucocephala, Gliricidia sepium and Flemingia macrophylla under humid tropical conditions. Agrofor. Syst. 1988, 7, 33–45. [Google Scholar] [CrossRef]
- Ge, T.; Chen, X.; Yuan, H.; Li, B.; Zhu, H.; Peng, P.; Li, K.; Jones, D.L.; Wu, J. Microbial biomass, activity, and community structure in horticultural soils under conventional and organic management strategies. Eur. J. Soil Biol. 2013, 58, 122–128. [Google Scholar] [CrossRef]
- Mann, L.K. Effects of whole-tree and stem-only clearcutting on postharvest hydrologic losses, nutrient capital, and regrowth. For. Sci. 1988, 34, 412–428. [Google Scholar]
- Webster, K.L.; Hazlett, P.W.; Brand, G.; Nelson, S.A.; Primavera, M.J.; Weldon, T.P. The effect of boreal jack pine harvest residue retention on soil environment and processes. For. Ecol. Manag. 2021, 497, 119517. [Google Scholar] [CrossRef]
- Hedwall, P.-O.; Grip, H.; Linder, S. Effects of clear-cutting and slash removal on soil water chemistry and forest-floor vegetation in a nutrient optimised Norway spruce stand. Silva Fenn. 2013, 47, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Ring, E.; Högbom, L.; Nohrstedt, H.-Ö.; Jacobson, S. Soil and soil-water chemistry below different amounts of logging residues at two harvested forest sites in Sweden. Silva Fenn. 2015, 49, 1265. [Google Scholar] [CrossRef] [Green Version]






| Index | T | % | S | % | S/Y | % | T × S | % | T × S/Y | % |
|---|---|---|---|---|---|---|---|---|---|---|
| Carbon sources 1 | ||||||||||
| AWCD | 2.32 | 2.01 | 48.61 *** | 28.02 | 29.74 *** | 17.14 | 1.78 | 3.08 | 2.61 * | 4.51 |
| Carbohydrates | 4.07 ** | 3.37 | 59.05 *** | 32.59 | 21.96 *** | 12.12 | 3.37 ** | 5.59 | 2.16 | 3.57 |
| Phenolic compounds | 1.16 | 1.31 | 9.87 *** | 34.59 | 26.11 *** | 17.55 | 0.86 | 2.95 | 1.77 | 1.61 |
| Polymers | 0.86 | 1.49 | 3.70 * | 8.43 | 46.58 *** | 22.29 | 0.81 | 2.21 | 4.24 *** | 4.52 |
| Carboxylic acids | 2.68 * | 2.48 | 35.49 *** | 16.19 | 17.17 *** | 11.63 | 2.18 * | 2.69 | 2.67 * | 14.99 |
| Amines | 0.18 | 0.90 | 36.77 *** | 2.60 | 9.49 *** | 32.75 | 4.00 *** | 1.72 | 1.77 | 8.95 |
| Amino acids | 1.62 | 0.20 | 64.24 *** | 26.49 | 32.59 *** | 6.83 | 1.83 | 8.65 | 0.99 | 3.81 |
| Diversity index 2 | ||||||||||
| H’ | 2.34 | 2.78 | 22.87 *** | 24.52 | 16.43 *** | 11.86 | 1.27 | 4.52 | 7.06 *** | 5.53 |
| D | 2.14 * | 2.63 | 8.03 *** | 6.58 | 15.80 *** | 12.96 | 1.23 | 3.02 | 5.24 *** | 12.90 |
| E | 6.43 *** | 8.39 | 0.43 | 0.37 | 3.33 * | 2.90 | 2.00 | 5.23 | 7.16 *** | 18.70 |
| PC | Chemical Type | Carbon Source | Loading Value |
|---|---|---|---|
| PC1 33.98% | Amine | L-Phenylalanine C4 | 0.835 |
| Phenolic compounds | 4-Hydroxy benzoic acid D3 | 0.792 | |
| Amino acid | L-Serine D4 | 0.772 | |
| Amino acid | L-Asparagine B4 | 0.742 | |
| Carbohydrates | D-Mannitol D2 | 0.645 | |
| Phenolic compounds | 2-Hydroxy benzoic acid C3 | 0.624 | |
| Amino acid | Phenylethylamine G4 | 0.56 | |
| Amino acid | L-Threonine E4 | 0.546 | |
| Amino acid | L-Arginine A4 | 0.546 | |
| PC2 9.25% | Carboxylic acids | D-Glucosaminic acid F2 | 0.808 |
| Polymers | Tween 40 C1 | 0.747 | |
| Amine | Putrescine H4 | 0.709 | |
| Carbohydrates | D-Galactonic acid lactone A3 | 0.689 | |
| Carboxylic acids | Pyruvic acid methyl ester B1 | 0.65 | |
| Carboxylic acids | γ-Hydroxybutyric butyric acid E3 | 0.648 | |
| Carbohydrates | N-Acetyl-D-glucosamine E2 | 0.565 | |
| PC3 6.97% | Carboxylic acids | D-Malic acid H3 | 0.684 |
| Carbohydrates | D-Xylose B2 | 0.641 | |
| Polymers | Tween 80 D1 | −0.568 | |
| Carboxylic acids | D-Galacturonic acid B3 | 0.567 | |
| Carbohydrates | Glucose-1-phosphate G2 | 0.558 | |
| Carbohydrates | D-Cellobiose G1 | 0.524 | |
| PC4 5.27% | Polymers | Glycogen F1 | 0.786 |
| Polymers | α-Cyclodextrin E1 | 0.689 | |
| Carbohydrates | I-Erythritol C2 | 0.617 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Gao, S.; Chen, J.; Yao, Z.; Zhang, L.; Wu, H.; Shu, Q.; Zhang, X. Response of Functional Diversity of Soil Microbial Community to Forest Cutting and Regeneration Methodology in a Chinese Fir Plantation. Forests 2022, 13, 360. https://doi.org/10.3390/f13020360
Wang X, Gao S, Chen J, Yao Z, Zhang L, Wu H, Shu Q, Zhang X. Response of Functional Diversity of Soil Microbial Community to Forest Cutting and Regeneration Methodology in a Chinese Fir Plantation. Forests. 2022; 13(2):360. https://doi.org/10.3390/f13020360
Chicago/Turabian StyleWang, Xu, Shenghua Gao, Jiquan Chen, Zengwang Yao, Lei Zhang, Hailong Wu, Qi Shu, and Xudong Zhang. 2022. "Response of Functional Diversity of Soil Microbial Community to Forest Cutting and Regeneration Methodology in a Chinese Fir Plantation" Forests 13, no. 2: 360. https://doi.org/10.3390/f13020360
APA StyleWang, X., Gao, S., Chen, J., Yao, Z., Zhang, L., Wu, H., Shu, Q., & Zhang, X. (2022). Response of Functional Diversity of Soil Microbial Community to Forest Cutting and Regeneration Methodology in a Chinese Fir Plantation. Forests, 13(2), 360. https://doi.org/10.3390/f13020360