
Soil Microbial Community Responds to Elevation Gradient in an Arid Montane Ecosystem in Northwest China

Abstract
:1. Introduction
2. Materials and Methods
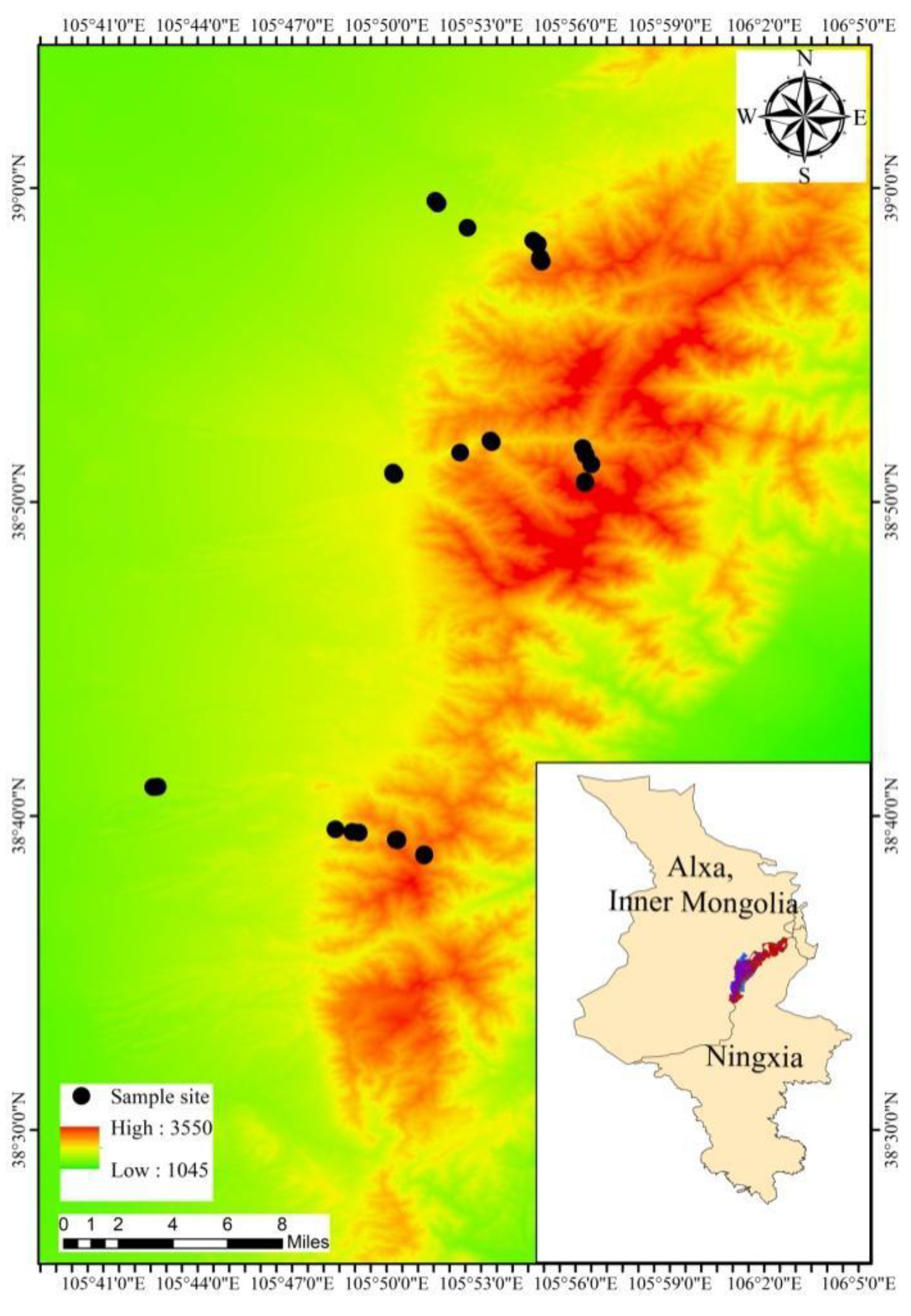
2.1. Study Area
2.2. Plots and Collecting of Samples
2.3. Soil Physical and Chemical Properties
2.4. Plant Diversity
2.5. Climate and Topography
2.6. High-Throughput Sequencing of Fungi and Bacteria
2.7. Data Analysis
3. Results
3.1. Soil, Climate, and Plant Diversity along the Elevation Gradient
3.2. Soil Microbial Diversity and Composition along the Elevation Gradient
3.2.1. Fungal and Bacterial Diversity
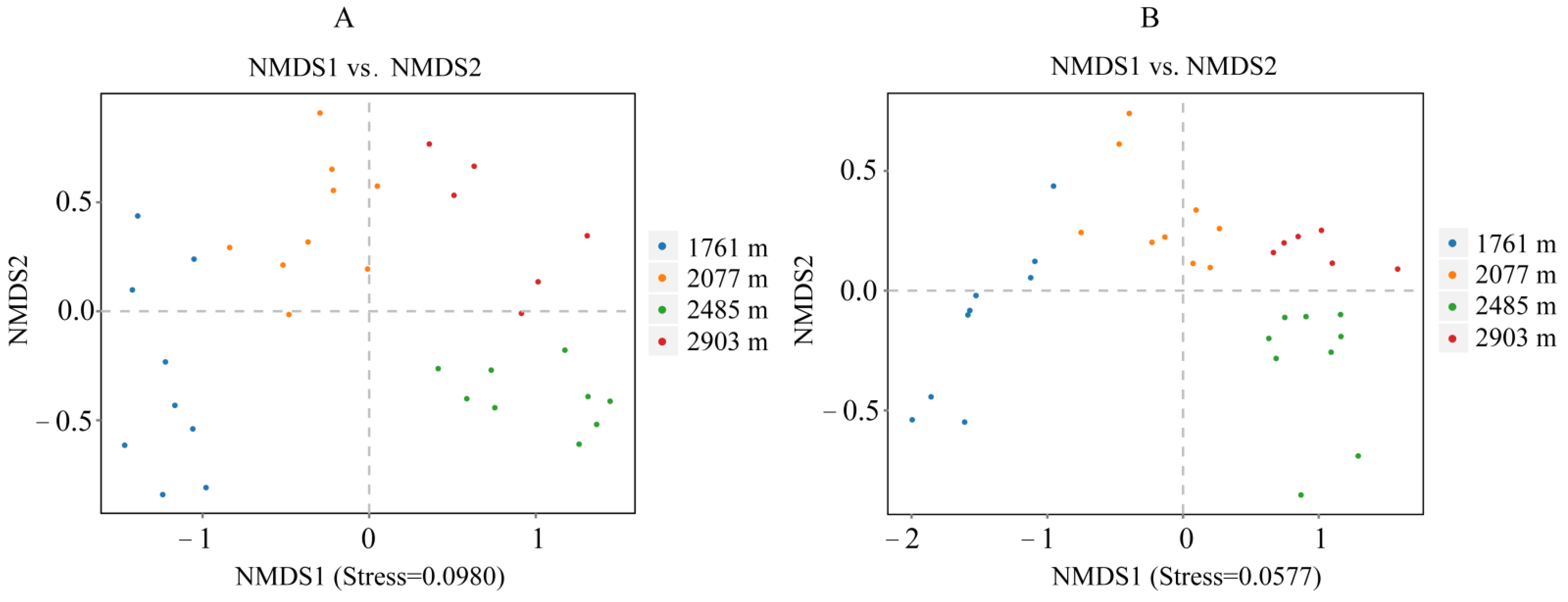
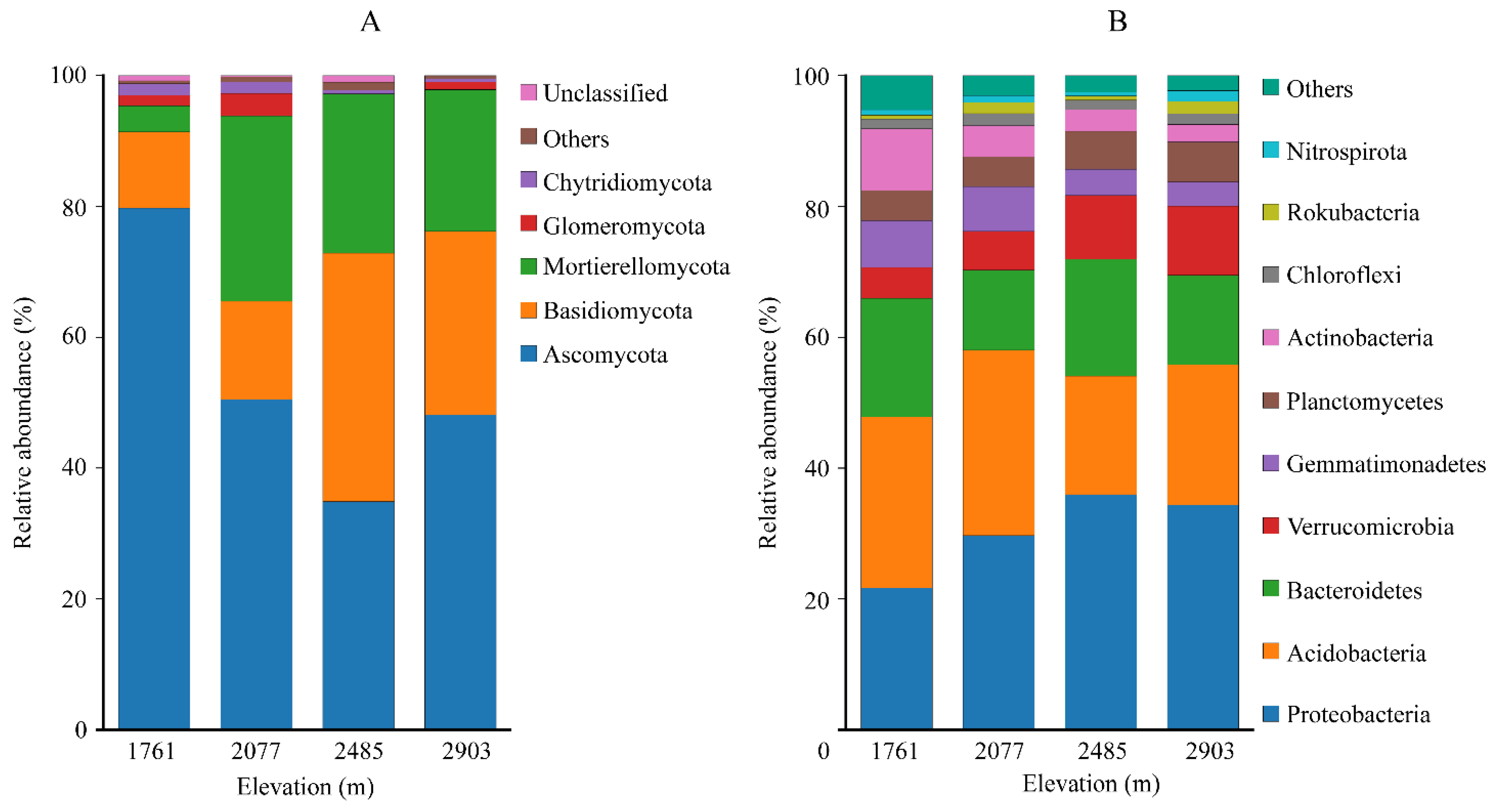
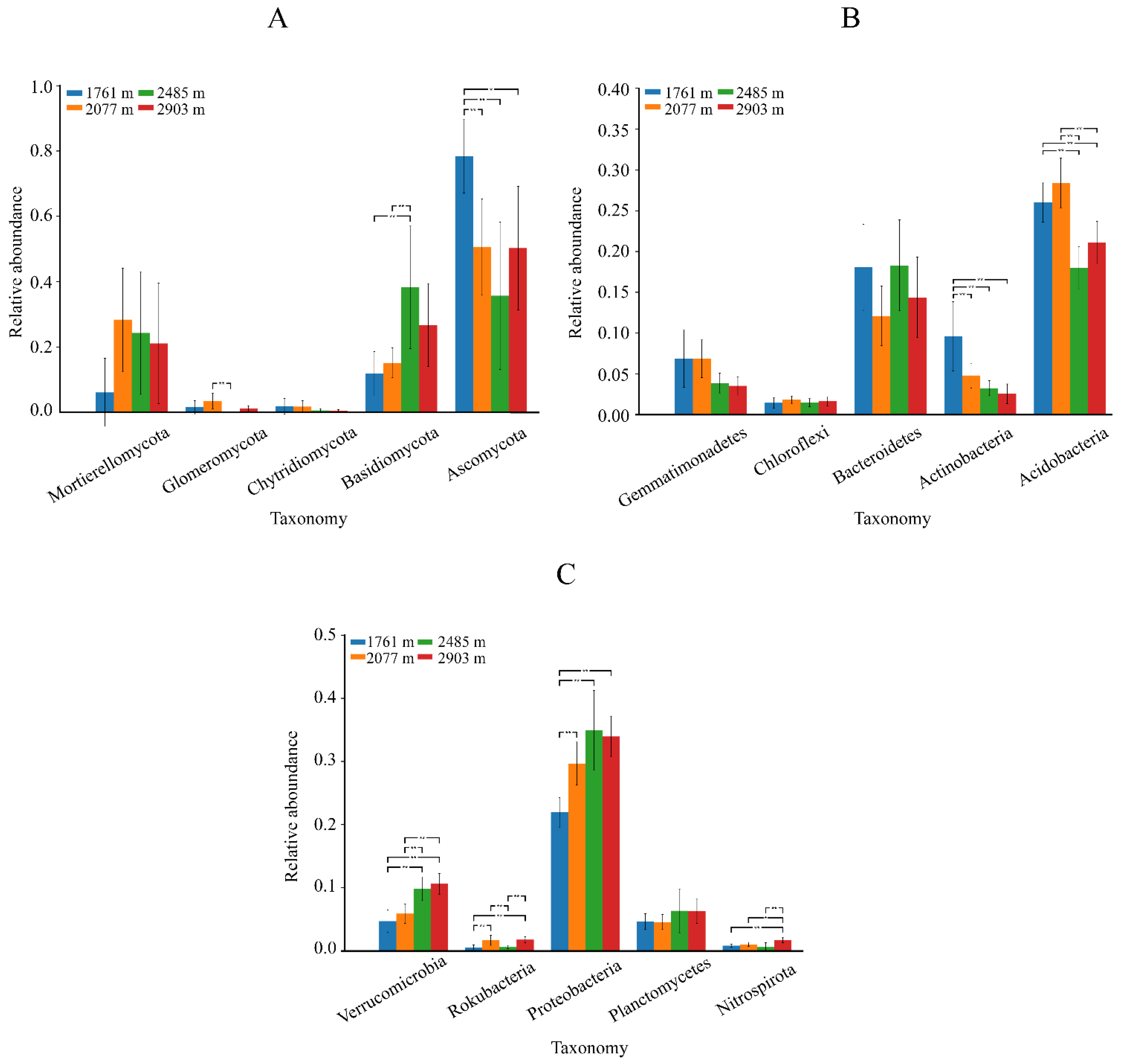
3.2.2. Fungal and Bacterial Community Composition
3.3. Soil Microbial Community and Environmental Factors
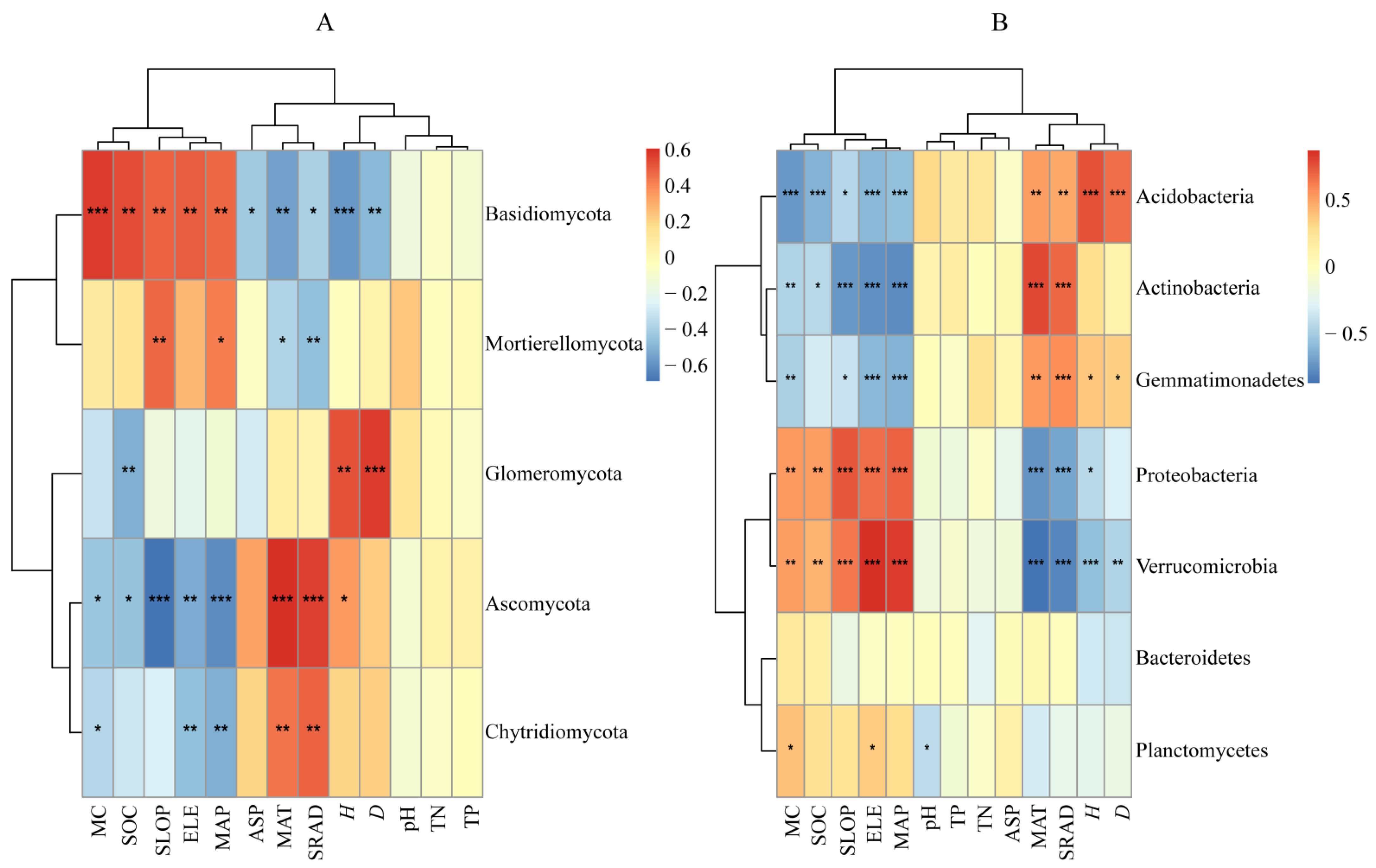
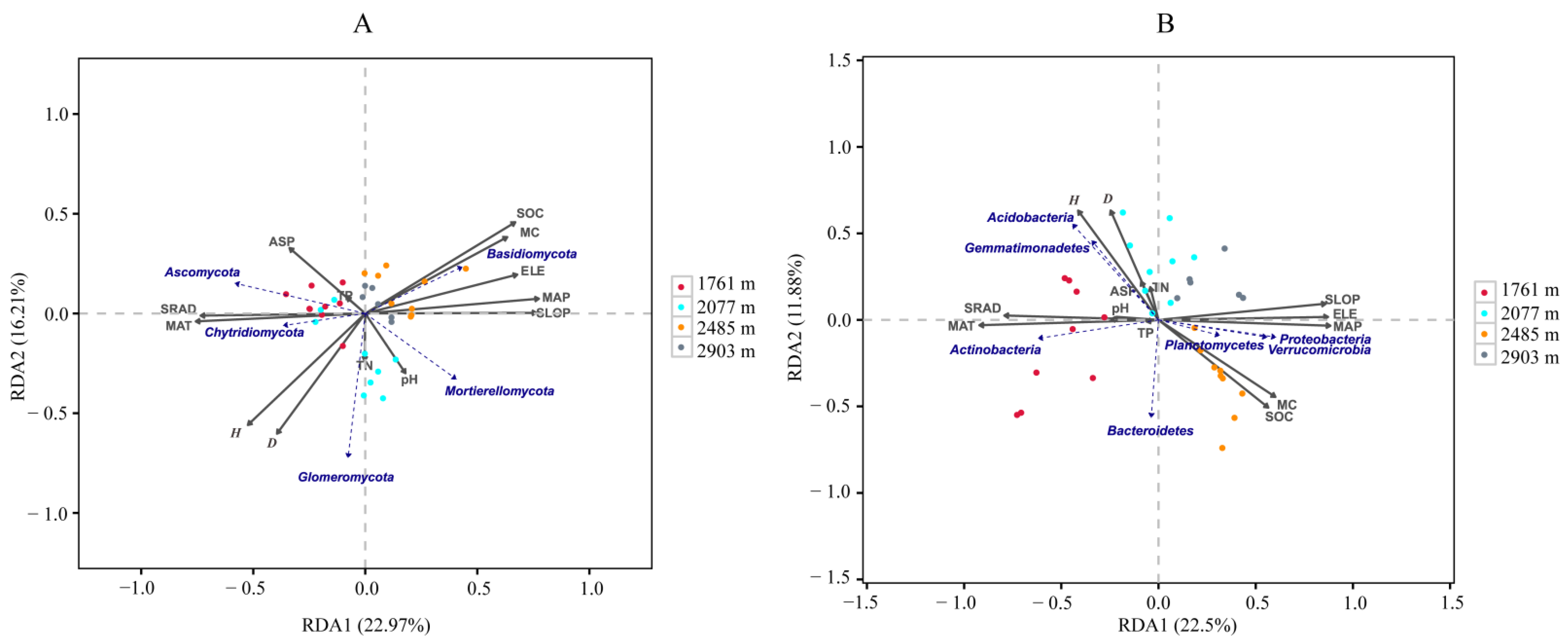
3.3.1. Relationships between Soil Microbial Composition and Environmental Factors
3.3.2. Relationships between Soil Microbial Diversity and Environmental Factors
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Van der Heijden, M.G.A.; Bardgett, R.D.; van Straalen, N.M. The unseen majority: Soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol. Lett. 2008, 11, 296–310. [Google Scholar] [CrossRef] [PubMed]
- Martiny, J.; Bohannan, B.J.M.; Brown, J.H.; Colwell, R.K.; Fuhrman, J.; Green, J.L.; Horner-Devine, M.C.; Kane, M.; Krumins, J.A.; Kuske, C.R.; et al. Microbial biogeography: Putting microorganisms on the map. Nat. Rev. Genet. 2006, 4, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Albright, M.B.N.; Johansen, R.; Thompson, J.; Lopez, D.; Gallegos-Graves, L.V.; Kroeger, M.E.; Runde, A.; Mueller, R.C.; Washburne, A.; Munsky, B.; et al. Soil bacterial and fungal richness forecast patterns of early pine litter decomposition. Front. Microbiol. 2020, 11, 542220. [Google Scholar] [CrossRef]
- Marinho, F.; Oehl, F.; da Silva, I.R.; Coyne, D.; Veras, J.S.D.N.; Maia, L.C. High diversity of arbuscular mycorrhizal fungi in natural and anthropized sites of a Brazilian tropical dry forest (Caatinga). Fungal Ecol. 2019, 40, 82–91. [Google Scholar] [CrossRef]
- Da Silva, C.A.; Londe, V.; Andrade, S.A.L.; Joly, C.A.; Vieira, S.A. Fine root-arbuscular mycorrhizal fungi interaction in Tropical Montane Forests: Effects of cover modifications and season. For. Ecol. Manag. 2020, 476, 118478. [Google Scholar] [CrossRef]
- Liu, S.; Moora, M.; Vasar, M.; Zobel, M.; Öpik, M.; Koorem, K. Arbuscular mycorrhizal fungi promote small-scale vegetation recovery in the forest understorey. Oecologia 2021, 197, 685–697. [Google Scholar] [CrossRef]
- Van Ruijven, J.; Ampt, E.; Francioli, D.; Mommer, L. Do soil-borne fungal pathogens mediate plant diversity–productivity relationships? Evidence and future opportunities. J. Ecol. 2020, 108, 1810–1821. [Google Scholar] [CrossRef]
- Finlay, B.J. Global dispersal of free-living microbial Eukaryote species. Science 2002, 296, 1061–1063. [Google Scholar] [CrossRef]
- O’Malley, M.A. The nineteenth century roots of ‘everything is everywhere’. Nat. Rev. Genet. 2007, 5, 647–651. [Google Scholar] [CrossRef]
- Fierer, N.; Jackson, R.B. The diversity and biogeography of soil bacterial communities. Proc. Natl. Acad. Sci. USA 2006, 103, 626–631. [Google Scholar] [CrossRef] [Green Version]
- Leff, J.W. Biogeography of Plant and Soil Microbial Communities; ProQuest Dissertations Publishing: Ann Arbor, MI, USA, 2016. [Google Scholar]
- O’Brien, S.L.; Gibbons, S.; Owens, S.M.; Hampton-Marcell, J.; Johnston, E.; Jastrow, J.; Gilbert, J.A.; Meyer, F.; Antonopoulos, D.A. Spatial scale drives patterns in soil bacterial diversity. Environ. Microbiol. 2016, 18, 2039–2051. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, L.; Lu, J.; Chen, W.; Wei, G.; Lin, Y. Topography affects the soil conditions and bacterial communities along a restoration gradient on Loess-Plateau. Appl. Soil Ecol. 2019, 150, 103471. [Google Scholar] [CrossRef]
- Zhang, B.; Liang, C.; He, H.; Zhang, X.; Smidt, H. Variations in soil microbial communities and residues along an altitude gradient on the northern slope of Changbai mountain, China. PLoS ONE 2013, 8, e66184. [Google Scholar] [CrossRef]
- Lei, T.; Si, G.; Wang, J.; Zhang, G. Microbial communities and associated enzyme activities in alpine wetlands with increasing altitude on the Tibetan Plateau. Wetlands 2017, 37, 401–412. [Google Scholar]
- Li, G.; Ma, K. Progress in the study of elevational patterns of soil microbial diversity. Acta Ecol. Sin. 2018, 38, 1521–1529. [Google Scholar]
- Singh, D.; Lee-Cruz, L.; Kim, W.-S.; Kerfahi, D.; Chun, J.-H.; Adams, J.M. Strong elevational trends in soil bacterial community composition on Mt. Halla, South Korea. Soil Biol. Biochem. 2014, 68, 140–149. [Google Scholar] [CrossRef]
- Wu, Z.; Lin, W.; Chen, Z.; Fang, C.; Zhang, Z.; Wu, L.; Zhou, M.; Chen, T. Variations of soil microbial community diversity along an elevational gradient in mid-subtropical forest. Chin. J. Plant Ecol. 2013, 37, 397–406. [Google Scholar] [CrossRef]
- Li, G.; Xu, G.; Shen, C.; Tang, Y.; Zhang, Y.; Ma, K. Contrasting elevational diversity patterns for soil bacteria between two ecosystems divided by the treeline. Sci. China Life Sci. 2016, 59, 1177–1186. [Google Scholar] [CrossRef]
- Su, C.; Zhang, X.; Wenhong, M. Altitudinal pattern and environmental interpretation of species diversity of Scrub communty in the Helan Mountains, China. Mt. Res. 2018, 36, 699–708. [Google Scholar]
- Su, C.; Ma, W.-H.; Zhang, X.-Y.; Su, Y.; Min, Y.-E.; Zhang, J.-Y.; Zhao, L.-Q.; Liang, C.-Z. Species composition and structure characteristics of the major montane shrub communities in the west slope of Helan Mountains, Nei Mongol, China. Chin. J. Plant Ecol. 2018, 42, 1050–1054. [Google Scholar] [CrossRef]
- Jiang, L.; Geng, Z.; Zhang, W.; Chen, X. Soil fertility under different forest types in the Helan and Liupan Mountain ranges of Ningxia Province. Acta Ecol. Sin. 2013, 33, 1982–1993. [Google Scholar] [CrossRef]
- Liu, B.; Zhang, X.; Hu, T.; Li, W. Soil microbial diversity under typical vegetation zones along an elevation gradient in Helan Mountains. Acta Ecol. Sin. 2013, 33, 7211–7220. [Google Scholar]
- Ma, J.; Pang, D.; Chen, L.; Wan, H.Y.; Chen, G.L.; Li, X.B. Characteristics of soil microbial community structure under vegetation at different altitudes in Helan Mountains. Acta Ecol. Sin. 2022, 42, 667–676. [Google Scholar]
- Yang, H.; Wang, P.; Li, X.; Wang, J.; Yang, J. Distribution of soil fungal diversity and community structure in different vegetation types on the eastern slopes of Helan Mountains. Ecol. Environ. Sci. 2022, 31, 239–247. [Google Scholar]
- Zheng, J.-G.; Chen, Y.-W.; Wu, G.-X. Association of Vegetation Patterns and Environmental Factors on the Arid Western Slopes of the Helan Mountains, China. Mt. Res. Dev. 2013, 33, 323–331. [Google Scholar] [CrossRef]
- Gregorich, E.G.; Carter, M.R. Soil Sampling and Methods of Analysis; CRC Press: Boca Raton, Fl, USA, 2007. [Google Scholar]
- Sun, Y.; Xia, F.; He, H.; Liu, B.; Wang, G.; Li, L. Community structure features of the larch-birch secondary forest on the northeastern slope of Changbai Mountain, Northeast China. J. Beijing For. Univ. 2016, 38, 28–38. [Google Scholar]
- Bellemain, E.; Carlsen, T.; Brochmann, C.; Coissac, E.; Taberlet, P.; Kauserud, H. ITS as an environmental DNA barcode for fungi: An in silico approach reveals potential PCR biases. BMC Microbiol. 2010, 10, 189. [Google Scholar] [CrossRef] [Green Version]
- Xu, N.; Tan, G.; Wang, H.; Gai, X. Effect of biochar additions to soil on nitrogen leaching, microbial biomass and bacterial community structure. Eur. J. Soil Biol. 2016, 74, 1–8. [Google Scholar] [CrossRef]
- Sun, Q.; Wu, H.; Chen, F.; Kang, J. Fungal community diversity and structure in rhizosphere soil of different crops in the arid zone of central Ningxia. Microbiol. China 2019, 46, 2963–2972. [Google Scholar]
- Hammer, D.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics software package for education and data analysis. Palacontologia Electron. 2021, 4, 9. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Ping, Y.; Han, D.; Wang, N.; Hu, Y.; Mu, L.; Feng, F. Vertical zonation of soil fungal community structure in a Korean pine forest on Changbai Mountain, China. World J. Microbiol. Biotechnol. 2016, 33, 12. [Google Scholar] [CrossRef] [PubMed]
- Man, B.; Xiang, X.; Luo, Y.; Mao, X.; Zhang, C.; Sun, B.; Wang, X. Characteristics and influencing factors of soil fungal community of typical vegetation types in Mount Huangshan, East China. Mycosystema 2021, 40, 2735–2751. [Google Scholar]
- Dassen, S.; Cortois, R.; Martens, H.; de Hollander, M.; Kowalchuk, G.A.; van der Putten, W.H.; De Deyn, G.B. Differential responses of soil bacteria, fungi, archaea and protists to plant species richness and plant functional group identity. Mol. Ecol. 2017, 26, 4085–4098. [Google Scholar] [CrossRef]
- Shen, C.; Wang, J.; He, J.-Z.; Yu, F.-H.; Ge, Y. Plant Diversity Enhances Soil Fungal Diversity and Microbial Resistance to Plant Invasion. Appl. Environ. Microbiol. 2021, 87, e00251-21. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Hou, L.; Zhang, Z.; Zhang, J.; Cheng, J.; Wei, G.; Lin, Y. Soil microbial diversity during 30 years of grassland restoration on the Loess Plateau, China: Tight linkages with plant diversity. Land Degrad. Dev. 2019, 30, 1172–1182. [Google Scholar] [CrossRef]
- Shen, C.; Liang, W.; Shi, Y.; Lin, X.; Zhang, H.; Wu, X.; Xie, G.; Chain, P.; Grogan, P.; Chu, H. Contrasting elevational diversity patterns between eukaryotic soil microbes and plants. Ecology 2014, 95, 3190–3202. [Google Scholar] [CrossRef]
- Johnson, D.; Booth, R.E.; Whiteley, A.S.; Bailey, M.J.; Read, D.J.; Grime, J.P.; Leake, J.R. Plant community composition affects the biomass, activity and diversity of microorganisms in limestone grassland soil. Eur. J. Soil Sci. 2003, 54, 671–678. [Google Scholar] [CrossRef]
- Shigyo, N.; Umeki, K.; Hirao, T. Plant functional diversity and soil properties control elevational diversity gradients of soil bacteria. FEMS Microbiol. Ecol. 2019, 95, 1–10. [Google Scholar] [CrossRef]
- Zuo, X.; Wang, S.; Lv, P.; Zhou, X.; Zhao, X.; Zhang, T.; Zhang, J. Plant functional diversity enhances associations of soil fungal diversity with vegetation and soil in the restoration of semiarid sandy grassland. Ecol. Evol. 2015, 6, 318–328. [Google Scholar] [CrossRef]
- Díaz, S.; Cabido, M. Vive la différence: Plant functional diversity matters to ecosystem processes. Trends Ecol. Evol. 2001, 16, 646–655. [Google Scholar] [CrossRef]
- Benvenutto-Vargas, V.P.; Ochoa-Hueso, R. Effects of nitrogen deposition on the spatial pattern of biocrusts and soil microbial activity in a semi-arid Mediterranean shrubland. Funct. Ecol. 2019, 34, 923–937. [Google Scholar] [CrossRef]
- Gray, S.B.; Classen, A.T.; Kardol, P.; Yermakov, Z.; Mille, R.M. Multiple Climate Change Factors Interact to Alter Soil Microbial Community Structure in an Old-Field Ecosystem. Soil Sci. Soc. Am. J. 2011, 75, 2217–2226. [Google Scholar] [CrossRef]
- Zhao, M.; Wang, M.; Zhao, Y.; Wang, G.; Xue, Z.; Jiang, M. Variations in soil microbial communities in the sedge-dominated peatlands along an altitude gradient on the northern slope of Changbai Mountain, China. Ecol. Indic. 2021, 129, 107964. [Google Scholar] [CrossRef]
- Zhang, R.; Tian, X.; Xiang, Q.; Penttinen, P.; Gu, Y. Response of soil microbial community structure and function to different altitudes in arid valley in Panzhihua, China. BMC Microbiol. 2022, 22, 86. [Google Scholar] [CrossRef] [PubMed]
- Bryant, J.A.; Lamanna, C.; Morlon, H.; Kerkhoff, A.J.; Enquist, B.J.; Green, J.L. Microbes on mountainsides: Contrasting elevational patterns of bacterial and plant diversity. Proc. Natl. Acad. Sci. USA 2008, 105, 11505–11511. [Google Scholar] [CrossRef]
- Shen, C.; Xiong, J.; Zhang, H.; Feng, Y.; Lin, X.; Li, X.; Liang, W.; Chu, H. Soil pH drives the spatial distribution of bacterial communities along elevation on Changbai Mountain. Soil Biol. Biochem. 2013, 57, 204–211. [Google Scholar] [CrossRef]
- Li, P.; Yin, R.; Zhou, H.; Yuan, X.; Feng, Z. Soil pH drives poplar rhizosphere soil microbial community responses to ozone pollution and nitrogen addition. Eur. J. Soil Sci. 2021, 73, e13186. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | Elevation (m a.s.l.) | |||
|---|---|---|---|---|
| 1761 m | 2077 m | 2485 m | 2903 m | |
| pH | 7.86 ± 0.32 | 7.86 ± 0.43 | 7.74 ± 0.37 | 7.66 ± 0.47 |
| MC (%) | 8.55 ± 3.32 b | 8.76 ± 2.98 b | 22.38 ± 10.16 a | 18.90 ± 2.97 a |
| TN (g·kg−1) | 0.34 ± 0.14 | 0.58 ± 0.84 | 0.29 ± 0.16 | 0.38 ± 0.17 |
| TP (g·kg−1) | 0.80 ± 1.11 | 0.74 ± 0.53 | 0.84 ± 0.65 | 0.22 ± 0.22 |
| SOC (g·kg−1) | 30.75 ± 27.61 b | 51.00 ± 25.77 b | 126.50 ± 38.42 a | 47.83 ± 13.50 b |
| ALT (m) | 1761.97 ± 173.43 d | 2077.74 ± 127.18 c | 2485.27 ± 78.73 b | 2903.99 ± 78.24 a |
| MAP (mm) | 211.33 ± 13.91 d | 240.22 ± 14.22 c | 265.00 ± 9.73 b | 287.50.11 ± 4.50 a |
| SRAD (kJ·m−2·day−1) | 16522.64 ± 115.98 d | 16394.59 ± 107.57 c | 16263.94 ± 88.82 b | 16092.92 ± 4.33 a |
| MAT (°C) | 6.19 ± 1.16 a | 3.63 ± 1.05 b | 1.88 ± 0.63 c | 0.55 ± 0.72 d |
| ASP (°) | 297.90 ± 41.18 ab | 234.92 ± 126.65 ab | 204.67 ± 137.18 b | 334.69 ± 9.17 a |
| SLOP (°) | 3.77 ± 1.86 c | 22.11 ± 4.70 b | 26.81 ± 3.08 a | 24.60 ± 2.02 ab |
| H | 1.54 ± 0.28 b | 2.31 ± 0.25 a | 0.44 ± 0.09 d | 1.11 ± 0.37 c |
| D | 1.41 ± 0.49 b | 3.17 ± 0.67 a | 0.26 ± 0.02 c | 0.99 ± 0.44 b |
| Elevation (m) | Fungi | Bacteria | ||
|---|---|---|---|---|
| Chao1 | Shannon | Chao1 | Shannon | |
| 1761 | 161.82 ± 51.61 b | 4.45 ± 1.21 b | 1276.54 ± 192.76 b | 8.77 ± 0.28 b |
| 2077 | 227.84 ± 63.34 a | 5.53 ± 0.37 a | 1659.75 ± 166.15 a | 9.10 ± 0.20 a |
| 2485 | 174.43 ± 33.06 ab | 4.45 ± 0.75 b | 1307.11 ± 150.58 b | 8.67 ± 0.32 b |
| 2903 | 165.22 ± 43.06 b | 4.60 ± 0.90 ab | 1345.40 ± 213.88 b | 8.82 ± 0.20 ab |
| Environment | Fungi | Bacteria | ||
|---|---|---|---|---|
| Chao1 | Shannon | Chao1 | Shannon | |
| pH | 0.450 ** | 0.148 | 0.034 | 0.046 |
| MC | −0.062 | −0.198 | −0.120 | −0.185 |
| TN | 0.296 | 0.189 | −0.018 | 0.100 |
| TP | 0.272 | 0.060 | 0.103 | 0.153 |
| SOC | −0.155 | −0.219 | −0.246 | −0.403 * |
| ALT | −0.029 | −0.161 | −0.016 | −0.073 |
| D | 0.306 | 0.286 | 0.461 ** | 0.461 ** |
| H | 0.356 * | 0.324 | 0.583 ** | 0.517 ** |
| MAT | −0.066 | 0.115 | −0.113 | 0.007 |
| MAP | 0.060 | −0.117 | 0.079 | −0.005 |
| SRAD | −0.108 | 0.118 | −0.130 | −0.068 |
| ASP | −0.125 | 0.033 | −0.087 | 0.186 |
| SLOP | 0.159 | 0.080 | 0.215 | 0.018 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.; Yu, J.; Wang, J.; Li, X. Soil Microbial Community Responds to Elevation Gradient in an Arid Montane Ecosystem in Northwest China. Forests 2022, 13, 1360. https://doi.org/10.3390/f13091360
Yang J, Yu J, Wang J, Li X. Soil Microbial Community Responds to Elevation Gradient in an Arid Montane Ecosystem in Northwest China. Forests. 2022; 13(9):1360. https://doi.org/10.3390/f13091360
Chicago/Turabian StyleYang, Junlong, Jian Yu, Jifei Wang, and Xiaowei Li. 2022. "Soil Microbial Community Responds to Elevation Gradient in an Arid Montane Ecosystem in Northwest China" Forests 13, no. 9: 1360. https://doi.org/10.3390/f13091360
APA StyleYang, J., Yu, J., Wang, J., & Li, X. (2022). Soil Microbial Community Responds to Elevation Gradient in an Arid Montane Ecosystem in Northwest China. Forests, 13(9), 1360. https://doi.org/10.3390/f13091360