
Soil Type Influences Rhizosphere Bacterial Community Assemblies of Pecan Plantations, a Case Study of Eastern China

Abstract
:1. Introduction
- Are the diversity and community composition of rhizosphere bacteria in pecan plantations affected by soil type?
- What are the compositions and biomarkers of the rhizosphere bacterial communities with pecan plantations grown in three soil types?
- What are the main soil factors driving the rhizosphere bacterial community of pecan plantations?
- What are the main functional types of rhizosphere bacteria in pecan plantations?
2. Materials and Methods
2.1. Study Area
2.2. Experimental Design and Sample Collection
2.3. Soil Property Determination
2.4. Soil DNA Extraction and PCR Amplification
2.5. Illumina MiSeq Sequencing
2.6. Processing of Sequencing Data
3. Results
3.1. Differences in Rhizosphere Soil Properties
3.2. The α-Diversity Analysis
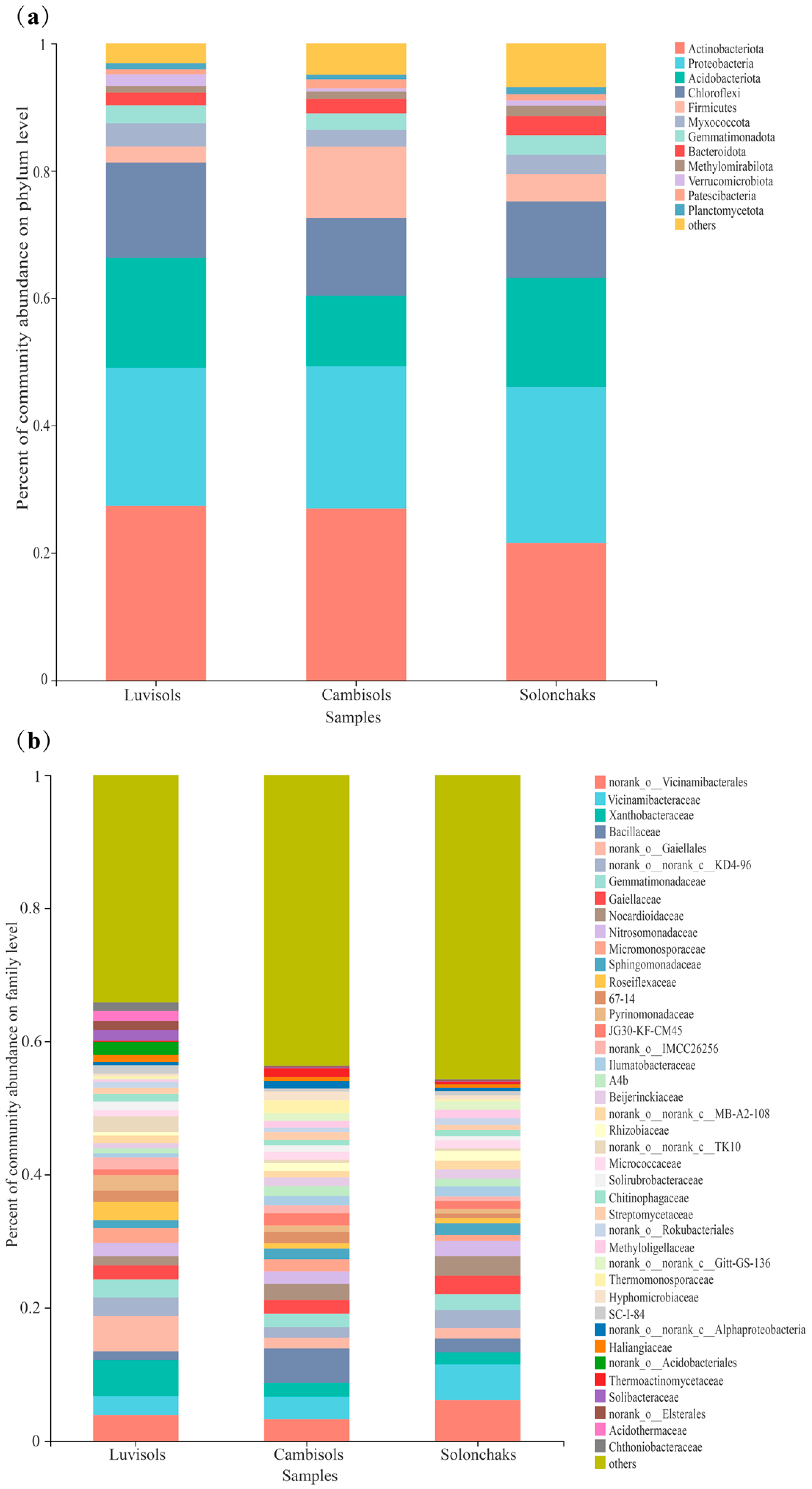
3.3. Compositions of the Rhizosphere Bacterial Communities
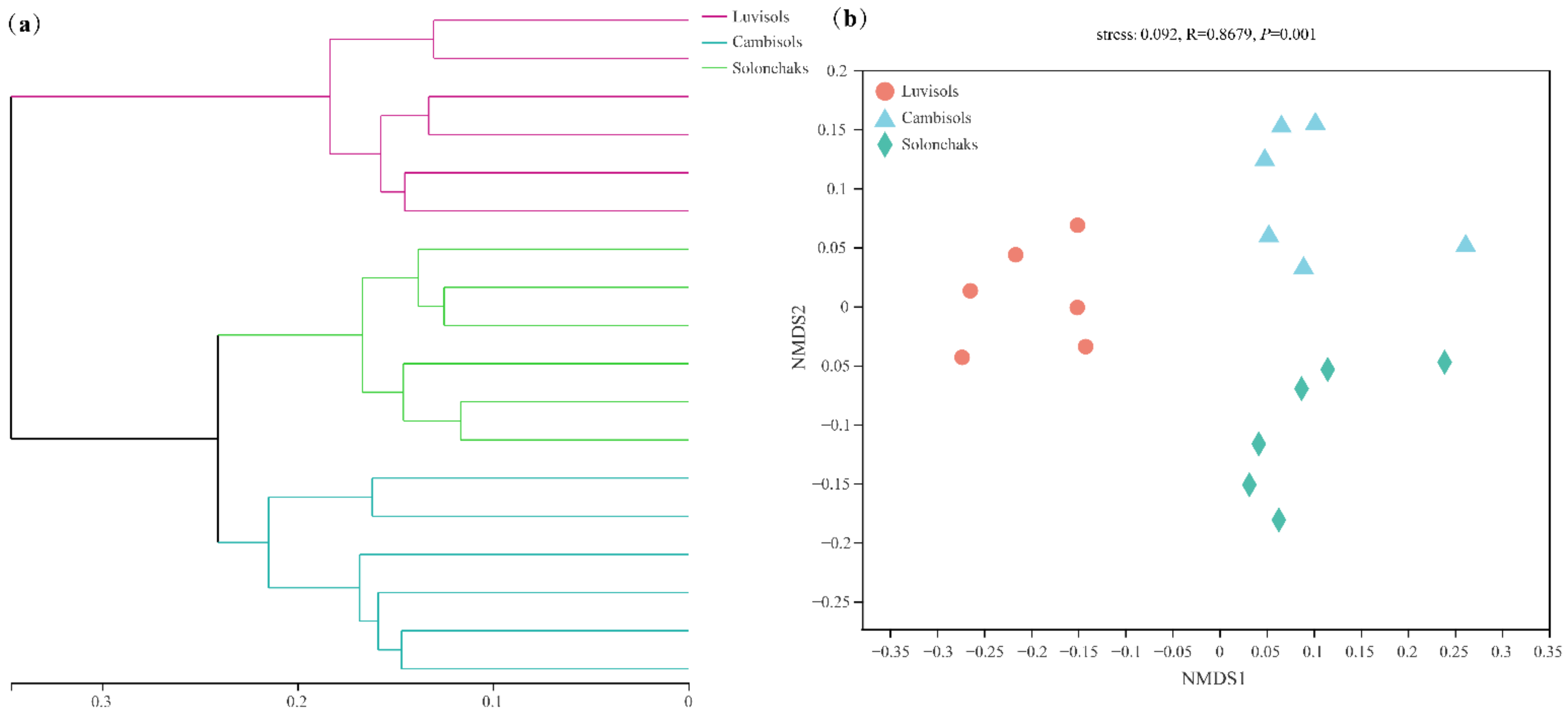
3.4. Differences in Rhizosphere Bacterial Community Structure
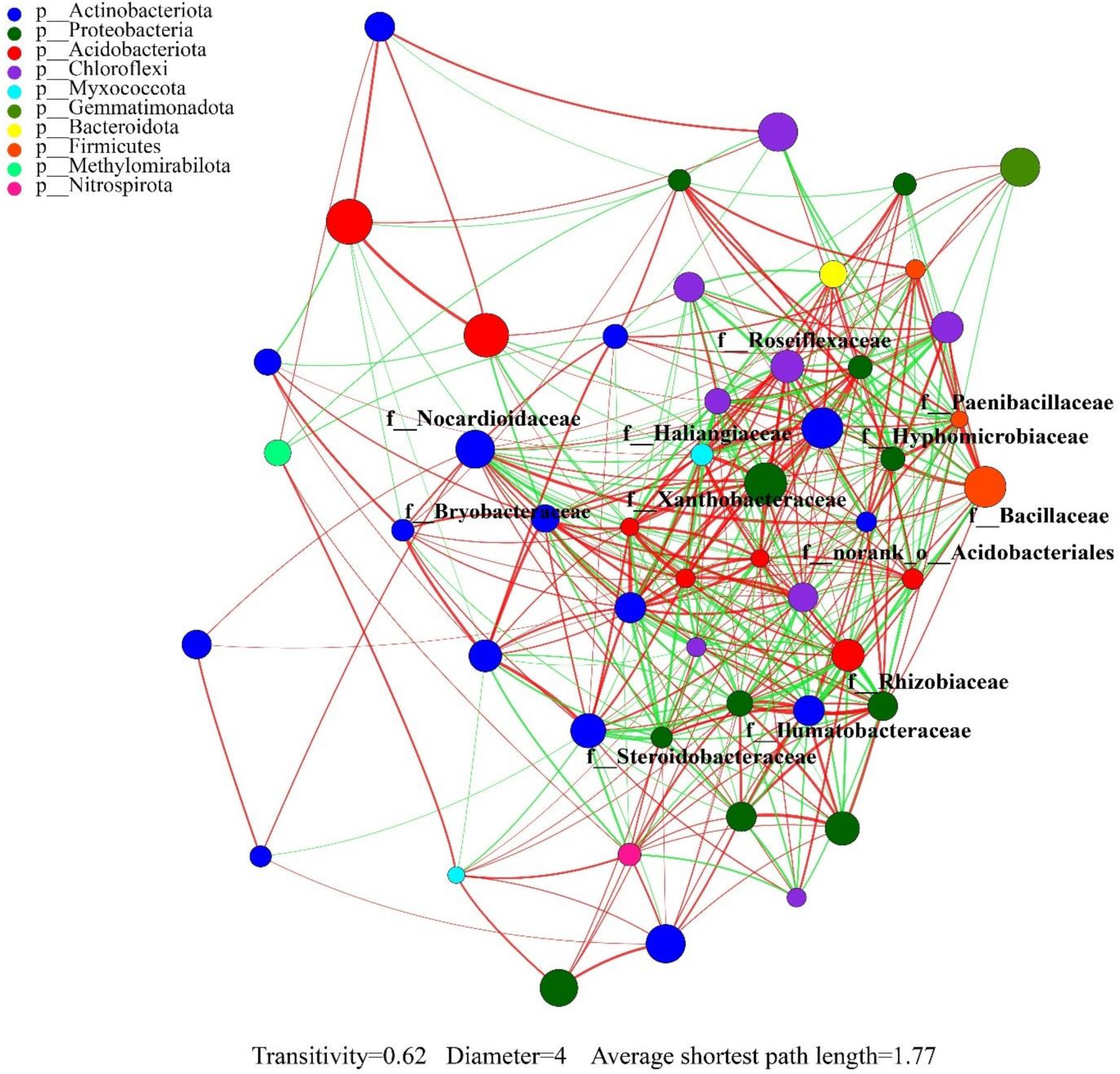
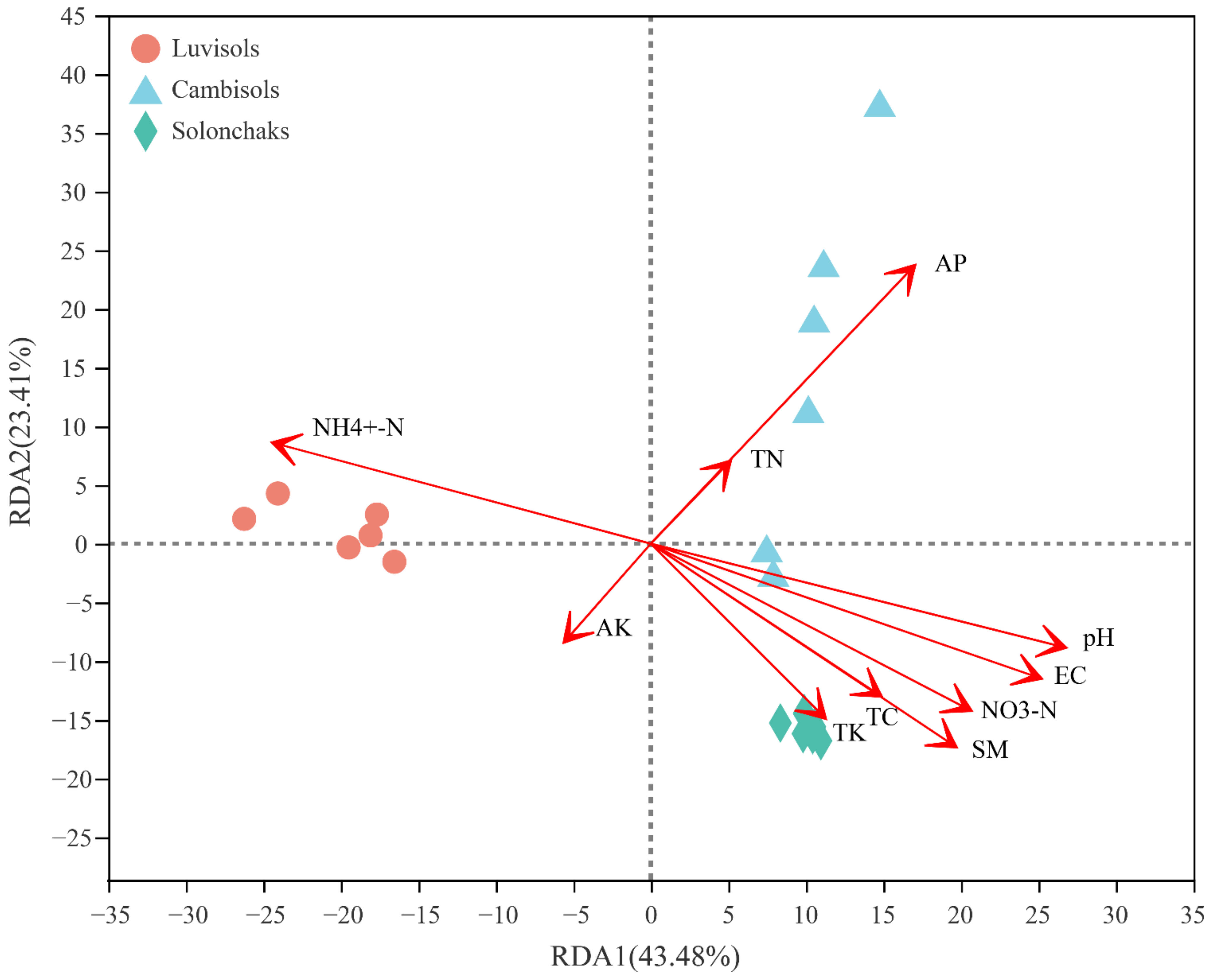
3.5. Correlation Analysis of Rhizosphere Soil Factors and Bacteria
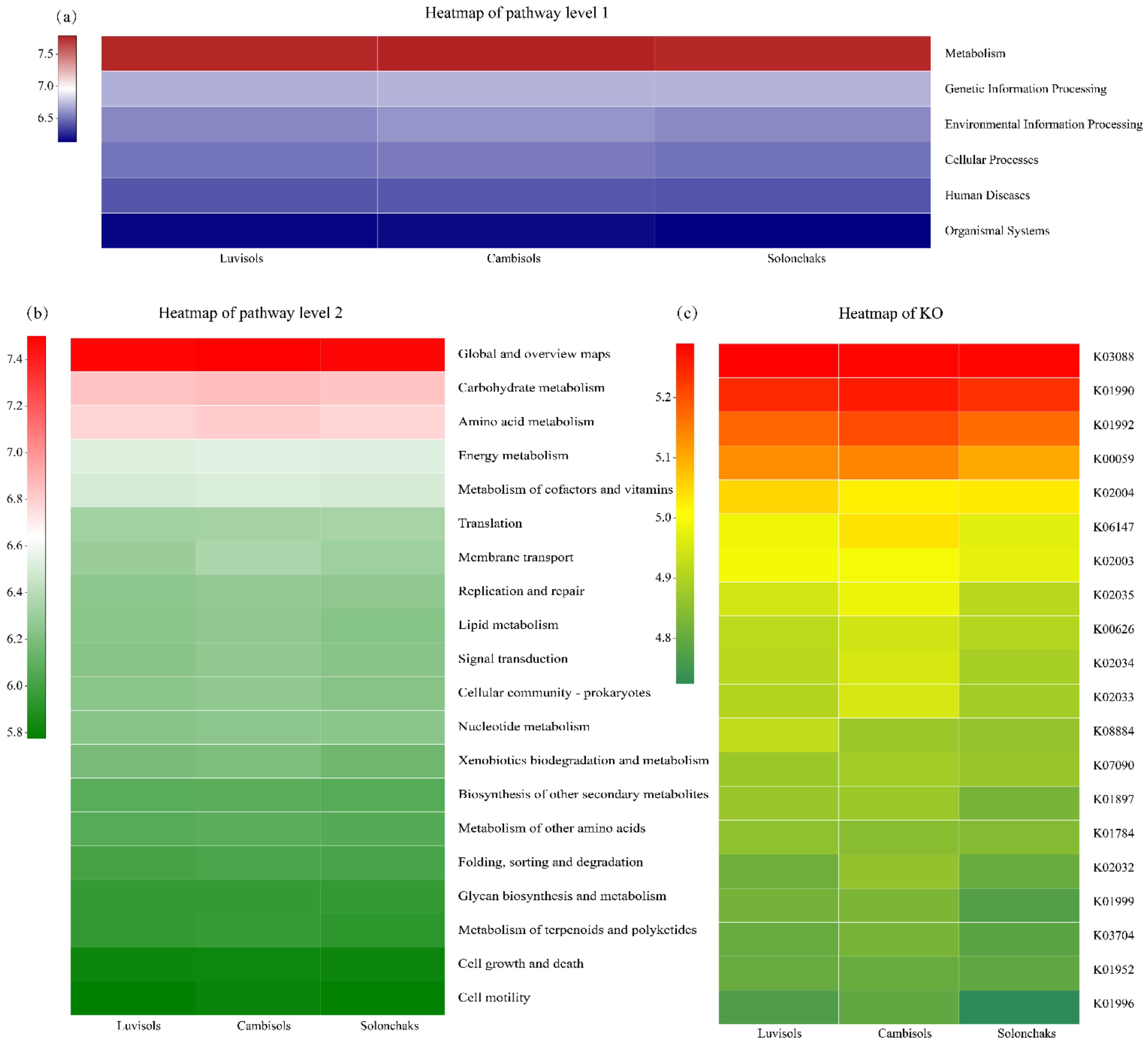
3.6. Functional Prediction of Rhizosphere Bacteria
4. Discussion
4.1. Effects of Different Soil Environments on the Rhizosphere Bacterial Diversity and Community Structure of Pecan Plantations
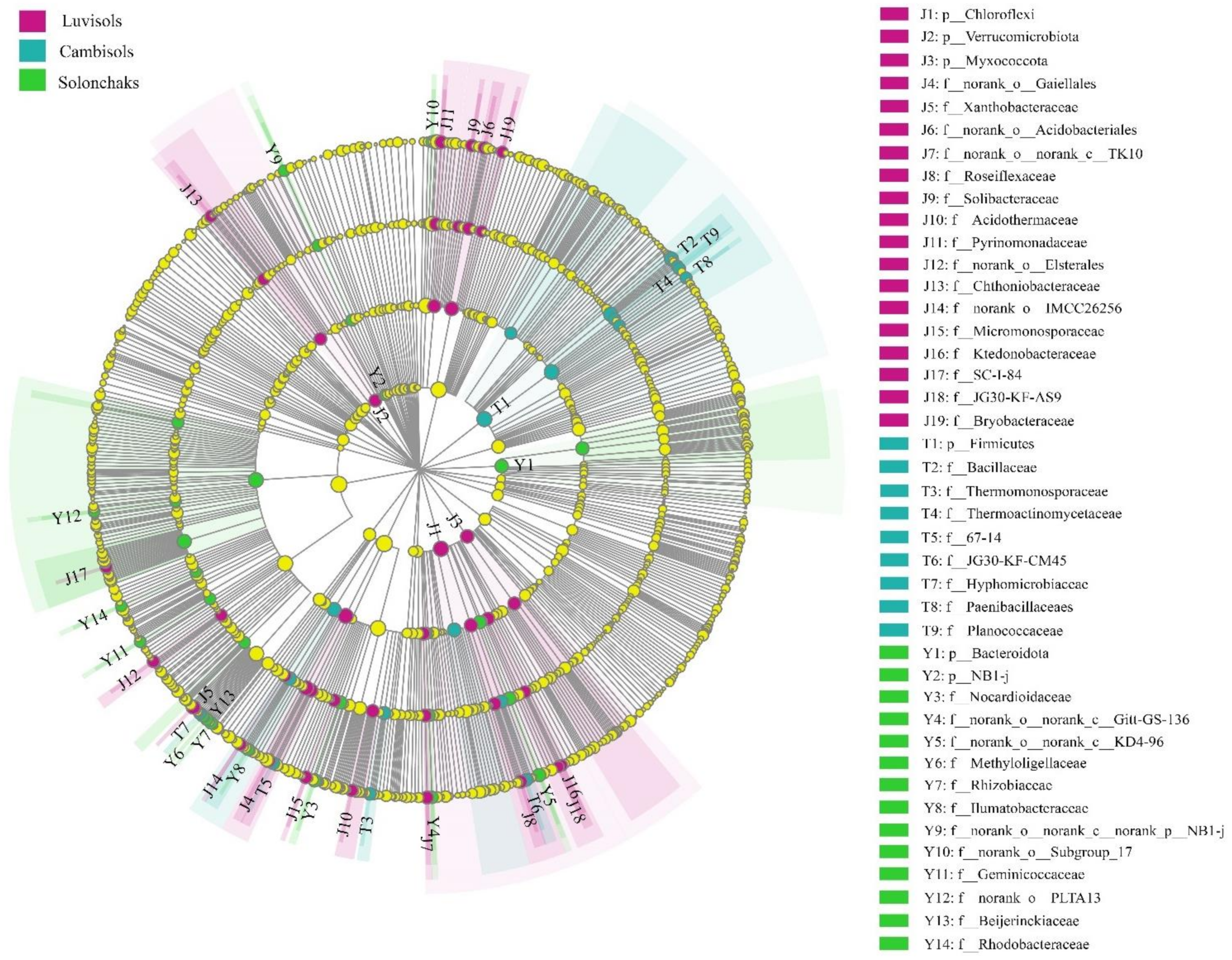
4.2. Biomarkers of Rhizosphere Bacteria in the Different Soil Types of Pecan Plantations
4.3. Potential Functions of Rhizosphere Bacteria in the Different Soil Types of Pecan Plantations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hartmann, A.; Rothballer, M.; Schmid, M. Lorenz Hiltner, a pioneer in rhizosphere microbial ecology and soil bacteriology research. Plant Soil 2008, 312, 7–14. [Google Scholar] [CrossRef]
- Peiffer, J.A.; Spor, A.; Koren, O.; Jin, Z.; Tringe, S.G.; Dangl, J.L.; Buckler, E.S.; Ley, R.E. Diversity and heritability of the maize rhizosphere microbiome under field conditions. Proc. Natl. Acad. Sci. USA 2013, 110, 6548–6553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonanomi, G.; De Filippis, F.; Cesarano, G.; La Storia, A.; Ercolini, D.; Scala, F. Organic farming induces changes in soil microbiota that affect agro-ecosystem functions. Soil Biol. Biochem. 2016, 103, 327–336. [Google Scholar] [CrossRef]
- Mendes, R.; Garbeva, P.; Raaijmakers, J.M. The rhizosphere microbiome: Significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiol. Rev. 2013, 37, 634–663. [Google Scholar] [CrossRef]
- Nacke, H.; Thurmer, A.; Wollherr, A.; Will, C.; Hodac, L.; Herold, N.; Schoning, I.; Schrumpf, M.; Daniel, R. Pyrosequencing-based assessment of bacterial community structure along different management types in German forest and grassland soils. PLoS ONE 2011, 6, e17000. [Google Scholar] [CrossRef] [Green Version]
- Bai, Y.; Muller, D.B.; Srinivas, G.; Garrido-Oter, R.; Potthoff, E.; Rott, M.; Dombrowski, N.; Munch, P.C.; Spaepen, S.; Remus-Emsermann, M.; et al. Functional overlap of the Arabidopsis leaf and root microbiota. Nature 2015, 528, 364–369. [Google Scholar] [CrossRef]
- Gottel, N.R.; Castro, H.F.; Kerley, M.; Yang, Z.M.; Pelletier, D.A.; Podar, M.; Karpinets, T.; Uberbacher, E.; Tuskan, G.A.; Vilgalys, R.; et al. Distinct microbial communities within the endosphere and rhizosphere of Populus deltoides roots across contrasting soil types. Appl. Env. Microbiol. 2011, 77, 5934–5944. [Google Scholar] [CrossRef] [Green Version]
- Lebeis, S.L.; Rott, M.; Dangl, J.L.; Schulze-Lefert, P. Culturing a plant microbiome community at the cross-Rhodes. New Phytol. 2012, 196, 341–344. [Google Scholar] [CrossRef]
- Berg, G.; Smalla, K. Plant species and soil type cooperatively shape the structure and function of microbial communities in the rhizosphere. FEMS Microbiol. Ecol. 2009, 68, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Seaton, F.M.; George, P.B.; Lebron, I.; Jones, D.L.; Creer, S.; Robinson, D.A. Soil textural heterogeneity impacts bacterial but not fungal diversity. Soil Biol. Biochem. 2020, 144, 107766. [Google Scholar] [CrossRef]
- Philippot, L.; Raaijmakers, J.M.; Lemanceau, P.; van der Putten, W.H. Going back to the roots: The microbial ecology of the rhizosphere. Nat. Rev. Microbiol. 2013, 11, 789–799. [Google Scholar] [CrossRef]
- Casales, F.G.; Watt, E.V.D.; Coetzer, G.M. Propagation of Pecan (Carya illinoensis): A review. Afr. J. Biotechnol. 2018, 17, 586–605. [Google Scholar] [CrossRef]
- Luo, X.; Li, Z.; Sun, Z.; Wan, X. Analysis of pecan cultivars Mahan and Western in East China. Genet. Mol. Res. 2016, 15, 1–12. [Google Scholar] [CrossRef]
- Zhang, R.; Peng, F.R.; Li, Y.R. Pecan production in China. Sci. Hortic. 2015, 197, 719–727. [Google Scholar] [CrossRef]
- Shapiro-Ilan, D.I.; Gardner, W.A.; Wells, L.; Cottrell, T.E.; Behle, R.W.; Wood, B.W. Effects of entomopathogenic fungus species, and impact of fertilizers, on biological control of pecan weevil (Coleoptera: Curculionidae). Env. Entomol. 2013, 42, 253–261. [Google Scholar] [CrossRef]
- Colombo, R.; Recchi, M.; Silvani, V.; Pérgola, M.; Martínez, A.; Godeas, A.M. Detection of arbuscular mycorrhizal fungi associated with pecan (Carya illinoinensis) trees by molecular and morphological approaches. MycoKeys 2018, 42, 73–88. [Google Scholar] [CrossRef]
- Iyer, R.; Damania, A. Draft genome sequence of Rhizobium sp. GHKF11, isolated from farmland soil in pecan Grove, Texas. Genome Announc. 2016, 4, e616–e682. [Google Scholar] [CrossRef] [Green Version]
- Riley, D.; Barber, S.A. Bicarbonate accumulation and pH changes at the soybean [Glycine max (L.) Merr.] root-soil interface. Soil Sci. Soc. Am. J. 1969, 33, 905–908. [Google Scholar] [CrossRef]
- Riley, D.; Barber, S.A. Salt accumulation at the soybean [Glycine max (L.) Merr.] root-soil interface. Soil Sci. Soc. Am. J. 1970, 34, 154–155. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, M.; Huang, S.Y.; Zha, X. Assessing spatial variability of soil organic carbon and total nitrogen in eroded hilly region of subtropical China. PLoS ONE 2020, 15, e244322. [Google Scholar] [CrossRef]
- Doane, T.A.; Horwáth, W.R. Spectrophotometric determination of nitrate with a single reagent. Anal. Lett. 2003, 36, 2713–2722. [Google Scholar] [CrossRef]
- Foster, J.C. Soil sampling, handling, storage and analysis. In Methods in Applied Soil Microbiology and Biochemistry; Academic Press: New York, NY, USA, 1995; pp. 49–121. [Google Scholar] [CrossRef]
- Woods, E.C.; McBride, S.M. Regulation of antimicrobial resistance by extracytoplasmic function (ECF) sigma factors. Microbes Infect. 2017, 19, 238–248. [Google Scholar] [CrossRef] [Green Version]
- Ponte-Sucre, A. Availability and applications of ATP-binding cassette (ABC) transporter blockers. Appl. Microbiol. Biotechnol. 2007, 76, 279–286. [Google Scholar] [CrossRef]
- Xian, J.P.; Wang, Y.; Niu, K.J.; Ma, H.L.; Ma, X. Transcriptional regulation and expression network responding to cadmium stress in a Cd-tolerant perennial grass Poa Pratensis. Chemosphere 2020, 250, 126158. [Google Scholar] [CrossRef]
- Ke, P.J.; Miki, T.; Ding, T.S. The soil microbial community predicts the importance of plant traits in plant-soil feedback. New Phytol. 2015, 206, 329–341. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.X.; Bing, H.J.; Fang, L.C.; Jiang, M.; Shen, G.T.; Yu, J.L.; Wang, X.; Zhu, H.; Wu, Y.H.; Zhang, X.C. Extracellular enzyme stoichiometry reveals the carbon and phosphorus limitations of microbial metabolisms in the rhizosphere and bulk soils in alpine ecosystems. Plant Soil 2019, 458, 7–14. [Google Scholar] [CrossRef]
- Erktan, A.; Or, D.; Scheu, S. The physical structure of soil: Determinant and consequence of trophic interactions. Soil Biol. Biochem. 2020, 148, 107876. [Google Scholar] [CrossRef]
- Liu, J.; Liu, M.; Wu, M.; Jiang, C.Y.; Chen, X.F.; Cai, Z.J.; Wang, B.R.; Zhang, J.; Zhang, T.L.; Li, Z.P. Soil pH rather than nutrients drive changes in microbial community following long-term fertilization in acidic Ultisols of southern China. J. Soils Sediments 2018, 18, 1853–1864. [Google Scholar] [CrossRef]
- Xun, W.B.; Huang, T.; Zhao, J.; Ran, W.; Wang, B.R.; Shen, Q.R.; Zhang, R.F. Environmental conditions rather than microbial inoculum composition determine the bacterial composition, microbial biomass and enzymatic activity of reconstructed soil microbial communities. Soil Biol. Biochem. 2015, 90, 10–18. [Google Scholar] [CrossRef]
- Cui, Y.X.; Fang, L.C.; Guo, X.B.; Wang, X.; Zhang, Y.J.; Li, P.F.; Zhang, X.C. Ecoenzymatic stoichiometry and microbial nutrient limitation in rhizosphere soil in the arid area of the northern Loess Plateau, China. Soil Biol. Biochem. 2018, 116, 11–21. [Google Scholar] [CrossRef]
- Zeng, Q.; Dong, Y.; An, S. Bacterial community responses to soils along a latitudinal and vegetation gradient on the Loess Plateau, China. PLoS ONE 2016, 11, e0152894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiao, P.; Li, Z.; Yang, L.; He, J.; Chang, X.; Xiao, H.; Nie, X.; Tong, D. Bacteria are more sensitive than fungi to moisture in eroded soil by natural grass vegetation restoration on the Loess Plateau. Sci. Total Environ. 2021, 756, 143899. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Xiao, X.; Huang, H.Y.; Jing, J.Q.; Zhao, H.J.; Wang, L.; Long, X.E. Contrasting beneficial and pathogenic microbial communities across consecutive cropping fields of greenhouse strawberry. Appl. Microbiol. Biotechnol. 2018, 102, 5717–5729. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Yu, M.; Lu, X.; Tang, C.; Liu, X.; Brookes, P.C.; Xu, J. Combined application of biochar and nitrogen fertilizer benefits nitrogen retention in the rhizosphere of soybean by increasing microbial biomass but not altering microbial community structure. Sci. Total Environ. 2018, 640, 1221–1230. [Google Scholar] [CrossRef]
- Meng, S.; Zhang, C.X.; Su, L.; Li, Y.M.; Cao, Y.; Zhao, Z. Distinct effect of pH on N uptake and assimilation in two conifer species. Trees 2016, 30, 1607–1618. [Google Scholar] [CrossRef]
- Mahoney, A.K.; Yin, C.T.; Hulbert, S.H. Community structure, species variation, and potential functions of rhizosphere-associated bacteria of different winter wheat (Triticum aestivum) Cultivars. Front. Plant Sci. 2017, 8, 132. [Google Scholar] [CrossRef] [Green Version]
- Janssen, P.H. Identifying the dominant soil bacterial taxa in libraries of 16S rRNA and 16S rRNA genes. Appl. Environ. Microbiol. 2006, 72, 1719–1728. [Google Scholar] [CrossRef] [Green Version]
- De Menezes, A.B.; Prendergast-Miller, M.T.; Poonpatana, P.; Farrell, M.; Bissett, A.; Macdonald, L.M.; Toscas, P.; Richardson, A.E.; Thrall, P.H. C/N ratio drives soil actinobacterial cellobiohydrolase gene diversity. Appl. Environ. Microbiol. 2015, 81, 3016–3028. [Google Scholar] [CrossRef] [Green Version]
- Barka, E.A.; Vatsa, P.; Sanchez, L.; Gaveau-Vaillant, N.; Jacquard, C.; Meier-Kolthoff, J.P.; Klenk, H.P.; Clément, C.; Ouhdouch, Y.; van Wezel, G.P. Taxonomy, physiology, and natural products of Actinobacteria. Microbiol. Mol. Biol. Rev. 2015, 80, 1–43. [Google Scholar] [CrossRef] [Green Version]
- Nelson, M.B.; Martiny, A.C.; Martiny, J.B. Global biogeography of microbial nitrogen-cycling traits in soil. Proc. Natl. Acad. Sci. USA 2016, 113, 8033–8040. [Google Scholar] [CrossRef] [Green Version]
- Karimi, B.; Terrat, S.; Dequiedt, S.; Saby, N.P.A.; Horriguel, W.; Lelievre, M.; Nowak, V.; Jolivet, C.; Arrouays, D.; Wincker, P.; et al. Biogeography of soil bacteria and archaea across France. Sci. Adv. 2018, 4, eaat1808. [Google Scholar] [CrossRef] [Green Version]
- Lebeis, S.L. The potential for give and take in plant-microbiome relationships. Front. Plant Sci. 2014, 5, 287. [Google Scholar] [CrossRef] [Green Version]
- Prober, S.M.; Leff, J.W.; Bates, S.T.; Borer, E.T.; Firn, J.; Harpole, W.S.; Lind, E.M.; Seabloom, E.W.; Adler, P.B.; Bakker, J.D.; et al. Plant diversity predicts beta but not alpha diversity of soil microbes across grasslands worldwide. Ecol. Lett. 2015, 18, 85–95. [Google Scholar] [CrossRef]
- Vacheron, J.; Desbrosses, G.; Bouffaud, M.L.; Touraine, B.; Moenne-Loccoz, Y.; Muller, D.; Legendre, L.; Wisniewski-Dye, F.; Prigent-Combaret, C. Plant growth-promoting rhizobacteria and root system functioning. Front. Plant Sci. 2013, 4, 356. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.Q.; Liu, Y.; Wang, Y.; Gao, F.; Zhao, J.H.; Li, Y.; Li, X.D. Effects of soil tillage, management practices, and mulching film application on soil health and peanut yield in a continuous cropping system. Front. Microbiol. 2020, 11, 570924. [Google Scholar] [CrossRef]
- Kanokratana, P.; Uengwetwanit, T.; Rattanachomsri, U.; Bunterngsook, B.; Nimchua, T.; Tangphatsornruang, S.; Plengvidhya, V.; Champreda, V.; Eurwilaichitr, L. Insights into the phylogeny and metabolic potential of a primary tropical peat swamp forest microbial community by metagenomic analysis. Microb. Ecol. 2011, 61, 518–528. [Google Scholar] [CrossRef]
- Lu, S.P.; Gischkat, S.; Reiche, M.; Akob, D.M.; Hallberg, K.B.; Kusel, K. Ecophysiology of Fe-cycling bacteria in acidic sediments. Appl. Environ. Microbiol. 2010, 76, 8174–8183. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Chen, Z.; Qian, J.; Wei, F.; Zhang, G.; Wang, Y.; Wei, G.; Hu, Z.; Dong, L.; Chen, S. Composition and function of rhizosphere microbiome of Panax notoginseng with discrepant yields. Chin. Med. 2020, 15, 85–97. [Google Scholar] [CrossRef]
- Mokni-Tlili, S.; Mehri, I.; Ghorbel, M.; Hassen, W.; Hassen, A.; Jedidi, N.; Hamdi, H. Community-level genetic profiles of actinomycetales in long-term biowaste-amended soils. Arch. Microbiol. 2020, 202, 2607–2617. [Google Scholar] [CrossRef]
- Wu, S.J.; Fotso, S.; Li, F.C.; Qin, S.; Laatsch, H. Amorphane sesquiterpenes from a marine Streptomyces sp. J. Nat. Prod. 2007, 70, 304–306. [Google Scholar] [CrossRef]
- Mandic-Mulec, I.; Stefanic, P.; van Elsas, J.D. Ecology of Bacillaceae. Microbiol. Spectr. 2015, 3, 17–2013. [Google Scholar] [CrossRef] [Green Version]
- Sandeep, C.; Raman, R.V.; Radhika, M.; Tejus, M.; Sanjeev, P.; Tejaswini, G.; Suresh, C.; Mulla, S.R. Effect of inoculation of Bacillus megaterium isolates on growth, biomass and nutrient content of Peppermint. J. Phytol. 2011, 3, 19–24. [Google Scholar]
- Ivanova, A.A.; Zhelezova, A.D.; Chernov, T.I.; Dedysh, S.N. Linking ecology and systematics of acidobacteria: Distinct habitat preferences of the Acidobacteriia and Blastocatellia in tundra soils. PLoS ONE 2020, 15, e230157. [Google Scholar] [CrossRef] [Green Version]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef]
- Luo, J.P.; Guo, X.Y.; Liang, J.B.; Song, Y.C.; Liu, Y.K.; Li, J.X.; Du, Y.L.; Mu, Q.L.; Jiang, Y.; Zhao, H.P.; et al. The influence of elevated CO2 on bacterial community structure and its co-occurrence network in soils polluted with Cr2O3 nanoparticles. Sci. Total Environ. 2021, 779, 146430. [Google Scholar] [CrossRef]
- Petters, S.; Groß, V.; Söllinger, A.; Pichler, M.; Reinhard, A.; Bengtsson, M.M.; Urich, T. The soil microbial food web revisited: Predatory myxobacteria as keystone taxa? ISME J. 2021, 15, 2665–2675. [Google Scholar] [CrossRef]
- López-Lozano, N.E.; Echeverría Molinar, A.; Ortiz Durán, E.A.; Hernandez Rosales, M.; Souza, V. Bacterial diversity and interaction networks of Agave lechuguilla rhizosphere differ significantly from bulk soil in the Oligotrophic Basin of Cuatro Cienegas. Front. Plant Sci. 2020, 11, 1028. [Google Scholar] [CrossRef]
- Cordovez, V.; Dini-Andreote, F.; Carrion, V.J.; Raaijmakers, J.M. Ecology and evolution of plant microbiomes. Annu. Rev. Microbiol. 2019, 73, 69–88. [Google Scholar] [CrossRef]
- Langille, M.G.I.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Thurber, R.L.V.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef] [PubMed]
- Landesman, W.J.; Nelson, D.M.; Fitzpatrick, M.C. Soil properties and tree species drive-diversity of soil bacterial communities. Soil Biol. Biochem. 2014, 76, 201–209. [Google Scholar] [CrossRef]
- Rahman, M.S.; Quadir, Q.F.; Rahman, A.; Asha, M.N.; Chowdhury, M.A.K. Screening and characterization of phosphorus solubilizing bacteria and their effect on rice seedlings. Res. Agric. Livest. Fish. 2015, 1, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Polónia, A.R.M.; Cleary, D.F.R.; Duarte, L.N.; de Voogd, N.J.; Gomes, N.C.M. Composition of Archaea in seawater, sediment, and sponges in the Kepulauan Seribu reef system, Indonesia. Microb. Ecol. 2014, 67, 553–567. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.X.; Hao, Z.P.; Sun, Y.Q.; Guo, L.P.; Huang, L.Q.; Zeng, Y.; Wang, Y.; Yang, L.; Chen, B.D. Comparison on the structure and function of the rhizosphere microbial community between healthy and root-rot Panax notoginseng. Appl. Soil Ecol. 2016, 107, 99–107. [Google Scholar] [CrossRef]
- Lee, S.H.; Ka, J.O.; Cho, J.C. Members of the phylum Acidobacteria are dominant and metabolically active in rhizosphere soil. FEMS Microbiol. Lett. 2008, 285, 263–269. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Type | Sampling Area | Altitude (m) | Soil Texture | Soil Bulk Density (g·cm−3) | Soil Porosity (%) | Average Tree Height (m) | Average Diameter at Breast Height (cm) | Average Crown (m) |
|---|---|---|---|---|---|---|---|---|
| Luvisols | Jurong | 24.21 | Loam | 1.39~1.50 | 44~48 | 7.47 | 9.21 | 3.95 |
| Cambisols | Taizhou | 7.85 | Sandy Loam | 1.31~1.46 | 45~51 | 7.21 | 9.76 | 4.03 |
| Solonchaks | Yancheng | 3.26 | Silt Loam | 1.12~1.42 | 46~58 | 6.23 | 8.53 | 3.79 |
| Soil Type | EC (μS·cm−1) | pH | SM (%) | TC (g·kg−1) | TN (g·kg−1) | NH4+-N (mg·kg−1) | NO3−-N (mg·kg−1) | AP (mg·kg−1) | AK (mg·kg−1) | TP (g·kg−1) | TK (g·kg−1) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Luvisols | 58.03 ± 18.74 b | 6.49 ± 0.64 b | 11.99 ± 1.39 b | 11.97 ± 1.78 b | 1.15 ± 0.62 a | 10.00 ± 1.09 a | 3.68 ± 0.69 b | 43.73 ± 8.28 b | 185.08 ± 68.41 a | 1.29 ± 0.08 c | 10.03 ± 0.63 b |
| Cambisols | 221.75 ± 84.20 a | 7.36 ± 0.07 a | 16.54 ± 2.66 b | 12.12 ± 2.09 b | 1.32 ± 0.89 a | 6.73 ± 2.22 b | 21.69 ± 14.63 a | 393.28 ± 123.97 a | 133.19 ± 35.15 a | 5.45 ± 0.31 a | 10.38 ± 0.45 b |
| Solonchaks | 268.42 ± 64.63 a | 7.76 ± 0.06 a | 26.92 ± 6.23 a | 16.38 ± 0.97 a | 1.28 ± 0.40 a | 5.45 ± 0.63 b | 39.32 ± 16.48 a | 72.23 ± 7.47 b | 186.61 ± 20.14 a | 1.60 ± 0.07 b | 14.24 ± 4.38 a |
| Soil Type | Sobs | Shannon | Simpson | Ace | Chao | Coverage |
|---|---|---|---|---|---|---|
| Luvisols | 3122.83 ± 191.28 b | 6.8481 ± 0.0997 b | 0.0026 ± 0.0003 a | 5048.02 ± 666.50 a | 4516.73 ± 294.67 b | 0.9582 ± 0.0034 a |
| Cambisols | 3597.83 ± 94.47 a | 7.0267 ± 0.0692 a | 0.0024 ± 0.0007 a | 5236.60 ± 169.37 a | 5207.75 ± 150.89 a | 0.9511 ± 0.0016 b |
| Solonchaks | 3474.83 ± 119.76 a | 7.0227 ± 0.0656 a | 0.0021 ± 0.0002 a | 5304.12 ± 580.52 a | 5049.36 ± 195.47 a | 0.9532 ± 0.0024 b |
| Phylum | Family | Group | LDA | Degree | Clustering | Degree Centrality | Closeness Centrality | Betweenness Centrality |
|---|---|---|---|---|---|---|---|---|
| Proteobacteria | Xanthobacteraceae | Luvisols | 4.27 | 24 | 0.66 | 0.49 | 0.65 | 0.01 |
| Acidobacteriota | norank_o__Acidobacteriales | Luvisols | 4.02 | 25 | 0.66 | 0.51 | 0.66 | 0.02 |
| Chloroflexi | Roseiflexaceae | Luvisols | 3.92 | 24 | 0.64 | 0.49 | 0.64 | 0.02 |
| Firmicutes | Bacillaceae | Cambisols | 4.28 | 21 | 0.59 | 0.43 | 0.62 | 0.02 |
| Proteobacteria | Hyphomicrobiaceae | Cambisols | 3.65 | 25 | 0.60 | 0.49 | 0.65 | 0.02 |
| Firmicutes | Paenibacillaceae | Cambisols | 3.54 | 24 | 0.56 | 0.49 | 0.65 | 0.03 |
| Actinobacteriota | Nocardioidaceae | Solonchaks | 3.92 | 23 | 0.58 | 0.47 | 0.61 | 0.04 |
| Proteobacteria | Rhizobiaceae | Solonchaks | 3.67 | 22 | 0.68 | 0.45 | 0.62 | 0.01 |
| Actinobacteriota | Ilumatobacteraceae | Solonchaks | 3.66 | 24 | 0.63 | 0.49 | 0.64 | 0.02 |
| Index | EC | pH | SM | TC | TN | NH4+-N | NO3−-N | AP | AK | TK |
|---|---|---|---|---|---|---|---|---|---|---|
| Sobs | 0.733 ** | 0.788 ** | 0.463 | 0.226 | 0.203 | −0.653 ** | 0.485 * | 0.561 * | −0.187 | 0.132 |
| Shannon | 0.724 ** | 0.812 ** | 0.535 * | 0.352 | 0.144 | −0.657 ** | 0.480* | 0.330 | −0.054 | 0.200 |
| Simpson | −0.500 * | −0.492 * | −0.352 | −0.193 | 0.125 | 0.256 | −0.415 | 0.280 | −0.027 | −0.277 |
| Ace | 0.337 | 0.515 * | 0.269 | 0.077 | 0.252 | −0.194 | 0.294 | 0.061 | 0.026 | 0.286 |
| Chao | 0.750 ** | 0.781 ** | 0.484 * | 0.190 | 0.163 | −0.638 ** | 0.528 * | 0.567 * | −0.229 | 0.279 |
| Coverage | −0.722 ** | −0.747 ** | −0.418 | −0.111 | −0.170 | 0.578 * | −0.512 * | −0.563 * | 0.254 | −0.175 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, Y.; Liu, J.; Bao, J.; Chu, G.; Peng, F. Soil Type Influences Rhizosphere Bacterial Community Assemblies of Pecan Plantations, a Case Study of Eastern China. Forests 2022, 13, 363. https://doi.org/10.3390/f13030363
Tang Y, Liu J, Bao J, Chu G, Peng F. Soil Type Influences Rhizosphere Bacterial Community Assemblies of Pecan Plantations, a Case Study of Eastern China. Forests. 2022; 13(3):363. https://doi.org/10.3390/f13030363
Chicago/Turabian StyleTang, Yujie, Junping Liu, Jiashu Bao, Guolin Chu, and Fangren Peng. 2022. "Soil Type Influences Rhizosphere Bacterial Community Assemblies of Pecan Plantations, a Case Study of Eastern China" Forests 13, no. 3: 363. https://doi.org/10.3390/f13030363
APA StyleTang, Y., Liu, J., Bao, J., Chu, G., & Peng, F. (2022). Soil Type Influences Rhizosphere Bacterial Community Assemblies of Pecan Plantations, a Case Study of Eastern China. Forests, 13(3), 363. https://doi.org/10.3390/f13030363