
The Effects of Biochar on Microbial Community Composition in and Beneath Biological Soil Crusts in a Pinus massoniana Lamb. Plantation

 ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Study Site
2.2. Experimental Design and Soil Sampling
2.3. Determination of Soil Physicochemical Properties and Enzyme Activities
2.4. Soil DNA Extraction, PCR Amplification, and High-Throughput Sequencing
2.5. Statistical Analysis
3. Results and Analysis
3.1. Soil Physicochemical Properties and Enzyme Activities
3.2. Soil Microbial Diversity
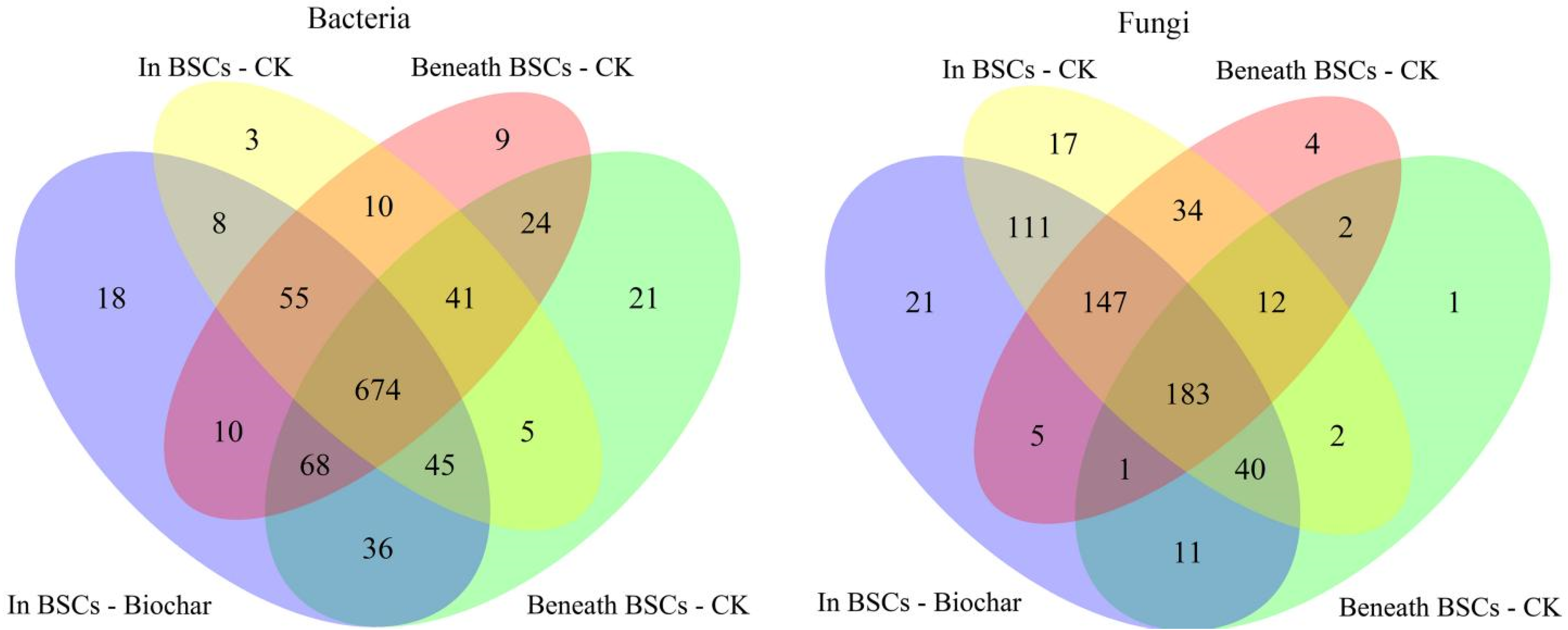
3.3. Soil Microbial Composition
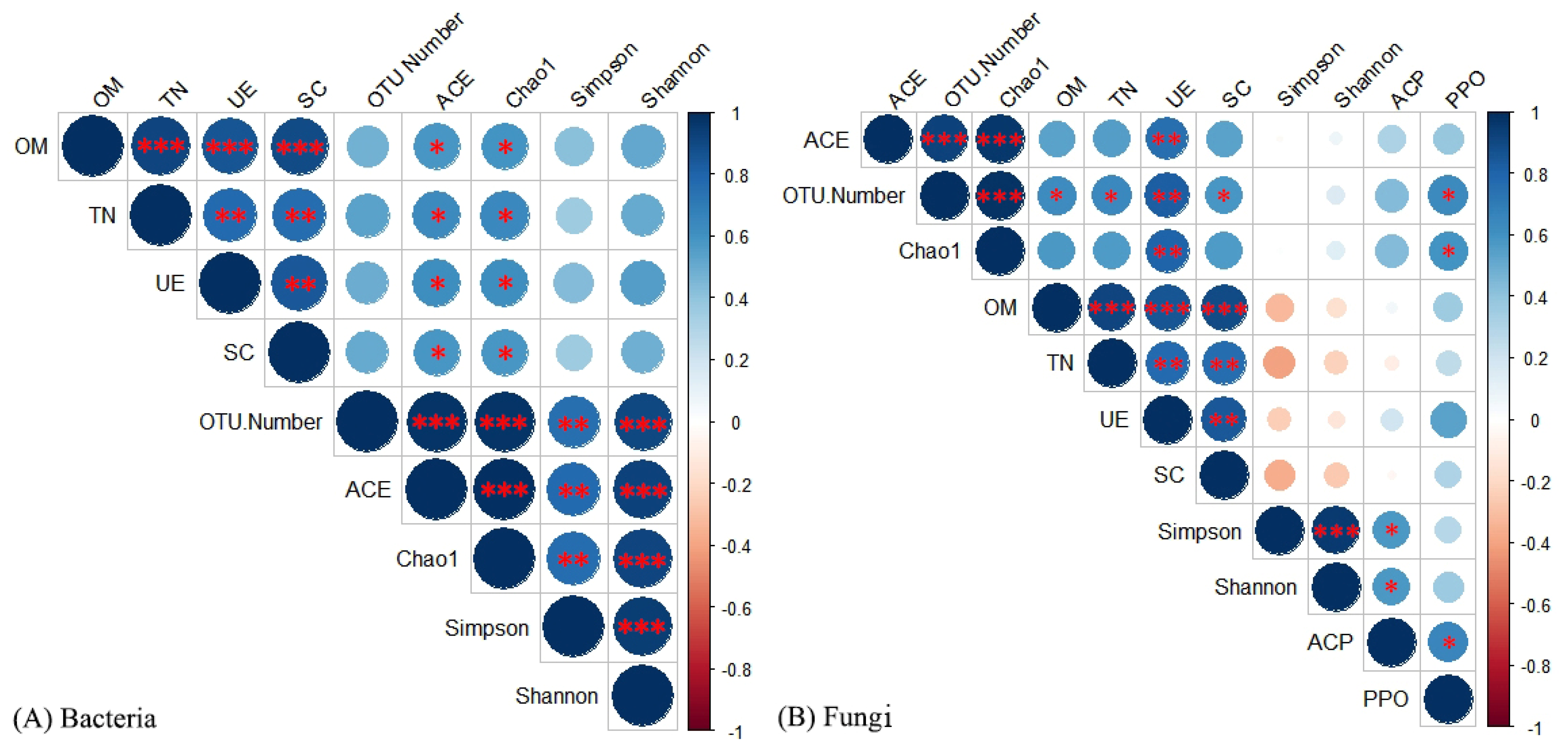
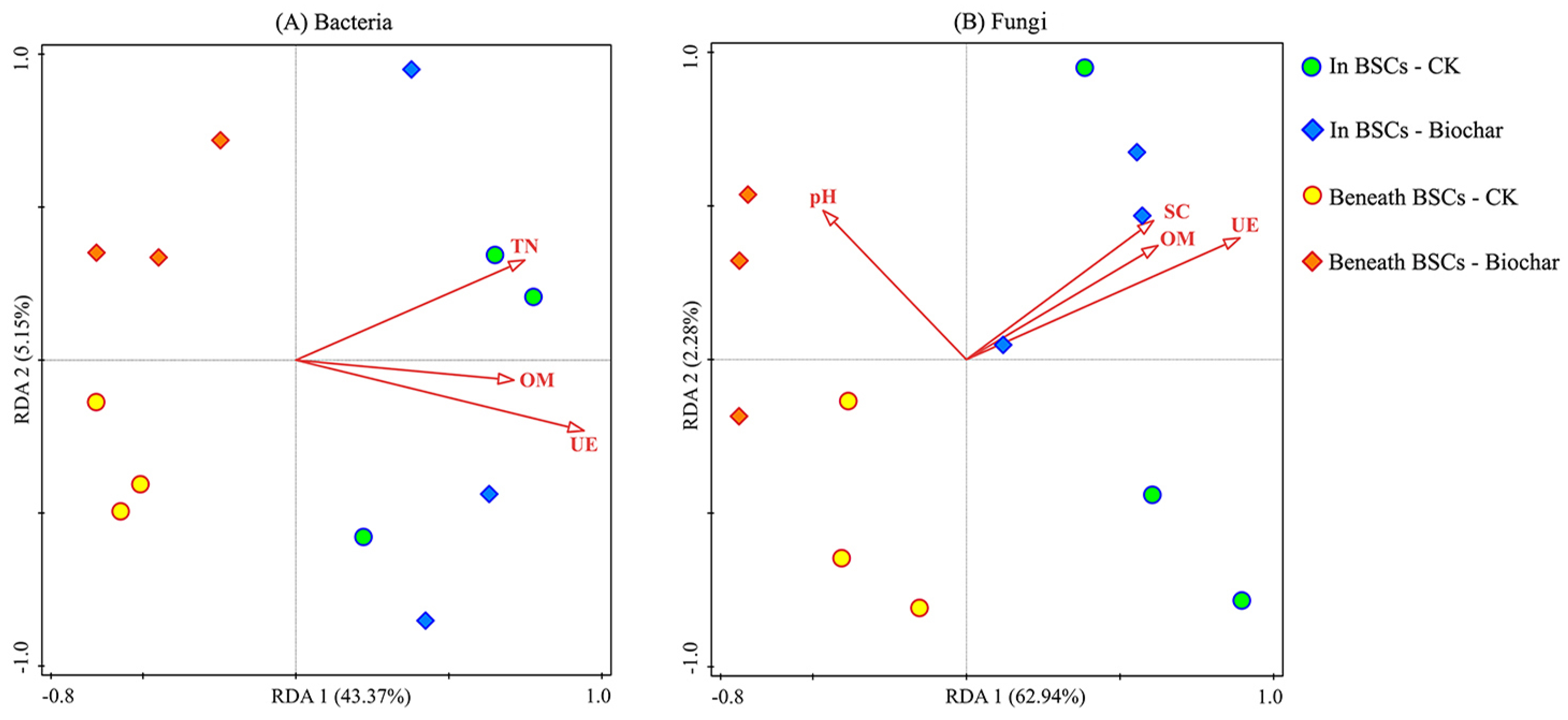
3.4. Relationship between Soil Microorganisms and Soil Environmental Factors
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Corbin, J.D.; Thiet, R.K. Temperate biocrusts: Mesic counterparts to their better-known dryland cousins. Front. Ecol. Environ. 2020, 18, 456–464. [Google Scholar] [CrossRef]
- Belnap, J.; Weber, B.; Büdel, B. Biological soil crusts as an organizing principle in drylands. In Biological Soil Crusts: An Organizing Principle in Drylands; Springer: Cham, Switzerland, 2016; pp. 3–13. [Google Scholar]
- Weber, B.; Belnap, J.; Büdel, B.; Antoninka, A.J.; Barger, N.N.; Chaudhary, V.B.; Darrouzet-Nardi, A.; Eldridge, D.J.; Faist, A.M.; Ferrenberg, S.; et al. What is a biocrust? A refined, contemporary definition for a broadening research community. Biol. Rev. 2022. Available online: https://onlinelibrary.wiley.com/doi/full/10.1111/brv.12862 (accessed on 20 May 2022).
- Rodriguez-Caballero, E.; Belnap, J.; Büdel, B.; Crutzen, P.J.; Andreae, M.O.; Pöschi, U.; Weber, B. Dry land photoautotrophic soil surface communities endangered by global change. Nat. Geosci. 2018, 11, 185–189. [Google Scholar] [CrossRef]
- Zhou, X.; Zhao, Y.; Belnap, J.; Zhang, B.; Bu, C.; Zhang, Y. Practices of biological soil crust rehabilitation in China: Experiences and challenges. Restor. Ecol. 2020, 28, S45–S55. [Google Scholar] [CrossRef]
- Belnap, J.; Lange, O.L. Biological Soil Crusts: Structure, Function, and Management; Springer: Heidelberg, Germany, 2003. [Google Scholar]
- Barger, N.N.; Harrick, J.E.; Zee, J.V.; Belnap, J. Impacts of biological soil crust disturbance and composition on C and N loss from water erosion. Biogeochemistry 2006, 77, 247–263. [Google Scholar] [CrossRef]
- Belnap, J. The world at your feet: Desert biological soil crusts. Front. Ecol. Environ. 2003, 1, 181. [Google Scholar] [CrossRef]
- Kuske, C.R.; Yeager, C.M.; Johnson, S.; Ticknor, L.O.; Belnap, J. Response and resilience of soil BSC bacterial communities to chronic physical disturbance in arid shrublands. ISME J. 2012, 6, 886–897. [Google Scholar] [CrossRef] [Green Version]
- Maestre, F.T.; Huesca, M.; Zaady, E.; Bautista, S.; Cortina, J. Infiltration, penetration resistance and microphytic crust composition in contrasted microsites within a Mediterranean semi-arid steppe. Soil Biol. Biochem. 2002, 34, 895–898. [Google Scholar] [CrossRef]
- Godinez-Alvarez, H.; Morin, C.; Rivera-Aguilar, V. Germination, survival and growth of three vascular plants on biological soil crusts from a Mexican tropical desert. Plant Biol. 2012, 14, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Bowker, M.A.; Reed, S.C.; Maestre, F.T.; Eldridge, D.J. Biocrusts: The living skin of the earth. Plant Soil 2018, 429, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Elbert, W.; Weber, B.; Burrow, S.; Steinkamp, J.; Büdel, B.; Andreae, M.O.; Pöschl, U. Contribution of cryptogamic covers to the global cycles of carbon and nitrogen. Nat. Geosci. 2012, 5, 459–462. [Google Scholar] [CrossRef]
- Lan, S.; Wu, L.; Zhang, D. Successional stages of biological soil crusts and their microstructure variability in Shapotou region (China). Environ. Earth Sci. 2012, 65, 77–88. [Google Scholar] [CrossRef]
- Büdel, B.; Williams, W.J.; Reichenberger, H. Annual net primary productivity of a cyanobacteria-dominated biological soil crust in the Gulf Savannah, Queensland, Australia. Biogeosciences 2018, 15, 491–505. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhao, L.; Wang, Z.; Liu, L.; Zhang, P.; Sun, J.; Wang, B.; Song, G.; Li, X. Changes in functional gene structure and metabolic potential of the microbial community in biological soil crusts along a revegetation chronosequence in the Tengger Desert. Soil Biol. Biochem. 2018, 126, 40–48. [Google Scholar] [CrossRef]
- Zhao, L.; Liu, Y.; Wang, Z.; Yuan, S.; Qi, J.; Zhang, W.; Wang, Y.; Li, X. Bacteria and fungi differentially contribute to carbon and nitrogen cycles during biological soil crust succession in arid ecosystems. Plant Soil 2020, 447, 379–392. [Google Scholar] [CrossRef]
- You, Y.; Aho, K.; Lohse, K.A.; Schwabedissen, S.G.; Ledbetter, R.N.; Magnuson, T.S. Biological soil crust bacterial communities vary along climatic and shrub cover gradients within a sagebrush steppe ecosystem. Front. Microbiol. 2021, 12, 569791. [Google Scholar] [CrossRef] [PubMed]
- Seitz, S.; Nebel, M.; Goebes, P.; Käppeler, K.; Schmidt, K.; Shi, X.; Song, Z.; Webber, C.L.; Weber, B.; Scholten, T. Bryophyte-dominated biological soil crusts mitigate soil erosion in an early successional Chinese subtropical forest. Biogeosciences 2017, 14, 5775–5788. [Google Scholar] [CrossRef] [Green Version]
- Song, Z.; Seitz, S.; Li, J.; Goebes, P.; Schmidt, K.; Kühn, P.; Shi, X.; Scholten, T. Tree diversity reduced soil erosion by affecting tree canopy and biological soil crust development in a subtropical forest experiment. Forest Ecol. Manag. 2019, 444, 69–77. [Google Scholar] [CrossRef]
- Harper, K.T.; Belnap, J. The influence of biological soil crusts on mineral uptake by associated vascular plants. J. Arid. Environ. 2001, 47, 347–357. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zhao, X.; Zhang, M. Functional diversity changes of microbial communities along a soil aquifer for reclaimed water recharge. FEMS Microbiol. Ecol. 2012, 80, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Yadav, K.K.; Sarkar, S. Biofertilizers, impact on soil fertility and crop productivity under sustainable agriculture. Environ. Ecol. 2019, 37, 89–93. [Google Scholar]
- Mager, D.M.; Thomas, A.D. Extracellular polysaccharides from cyanobacterial soil crusts: A review of their role in dryland soil processes. J. Arid Environ. 2011, 75, 91–97. [Google Scholar] [CrossRef]
- Garbeva, P.; van Veen, J.A.; van Elsas, J.D. Microbial diversity in soil: Selection microbial populations by plant and soil type and implications for disease suppressiveness. Annu. Rev. Phytopathol. 2004, 42, 243–270. [Google Scholar] [CrossRef] [PubMed]
- Yarwood, S.A. The role of wetland microorganisms in plant-litter decomposition and soil organic matter formation: A critical review. FEMS Microbiol. Ecol. 2018, 94, fiy175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joshi, H.; Somduttand, C.P.; Mundra, S.L. Role of effective microorganisms (EM) in sustainable agriculture. Int. J. Curr. Microbiol. Appl. Sci. 2019, 8, 172–181. [Google Scholar] [CrossRef]
- Ngosong, C.; Buse, T.; Ewald, M.; Richter, A.; Glaser, K. Influence of management intensity and environmental conditions on microbiota in biological soil crust and crust-free soil habitats of temperate forests. Soil Biol. Biochem. 2020, 144, 107761. [Google Scholar] [CrossRef]
- Singh, C.; Tiwari, S.; Gupta, V.K.; Singh, J.S. The effect of rice husk biochar on soil nutrient status, microbial biomass and paddy productivity of nutrient poor agriculture soils. Catena 2018, 171, 485–493. [Google Scholar] [CrossRef]
- Cao, X.; Ma, L.; Gao, B.; Harris, W. Dairy-manure derived biochar effectively sorbs lead and atrazine. Environ. Sci. Technol. 2009, 43, 3285–3291. [Google Scholar] [CrossRef]
- Spokas, K.A.; Koskinen, W.C.; Baker, J.M.; Reicosky, D.C. Impacts of woodchip biochar additions on greenhouse gas production and sorption/degradation of two herbicides in a Minnesota soil. Chemosphere 2009, 77, 574–581. [Google Scholar] [CrossRef]
- Atkinson, C.J.; Fitzgerald, J.D.; Hipps, N.A. Potential mechanisms for achieving agricultural benefifits from biochar application to temerate soils: A review. Plant Soil 2010, 337, 1–18. [Google Scholar] [CrossRef]
- Sohi, S.P.; Krull, E.; Lopez-Capel, E.; Bol, R. A review of biochar and its use and function in soil. Adv. Agron. 2010, 105, 47–82. [Google Scholar]
- Anderson, C.R.; Condron, L.M.; Clough, T.J.; Fiers, M.; Stewart, A.; Hill, R.A.; Sherlock, R.R. Biochar induced soil microbial community change: Implications for biogeochemical cycling of carbon, nitrogen and phosphorus. Pedobiologia 2011, 54, 309–320. [Google Scholar] [CrossRef]
- Palansooriya, K.N.; Wong, J.T.F.; Hashimoto, Y.; Huang, L.; Rinklebe, J.; Chang, S.X.; Bolan, N.; Wang, H.; Ok, Y.S. Response of microbial communities to biochar-amended soil: A critical review. Biochar 2019, 1, 3–22. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.; Yuan, W. Can biochar couple with algae to deal with desertification? J. Sustain. Bioenergy Syst. 2014, 4, 194–198. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Guo, J.; Chen, G.; Yin, Y.; Gao, R.; Lin, C. Effects of forest conversion on soil labile organic carbon fractions and aggregate stability in subtropical China. Plant Soil 2009, 323, 153–162. [Google Scholar] [CrossRef]
- Hill, R.D.; Peart, M.R. Land use, runoff, erosion and their control: A review for southern China. Hydrol. Process. 1998, 12, 2029–2042. [Google Scholar] [CrossRef]
- Guo, Q.; Hao, Y.; Liu, B. Rates of soil erosion in China: A study based on runoff plot data. Catena 2015, 124, 68–76. [Google Scholar] [CrossRef]
- Wang, X.; Kent, M.; Fang, X. Evergreen broad-leaved forest in Eastern China: Its ecology and conservation and the importance of resprouting in forest restoration. Forest Ecol. Manag. 2007, 245, 76–87. [Google Scholar] [CrossRef]
- Seitz, S.; Goebes, P.; Song, Z.; Bruelheide, H.; Härdtle, W.; Kühn, P.; Li, Y.; Scholten, T. Tree species and functional traits but not species richness affect interrill erosion processes in young subtropical forests. Soil 2016, 2, 49–61. [Google Scholar] [CrossRef] [Green Version]
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon, and organic matter. In Methods Soil Analysis; American Society of Agronomy and Soil Science Society of America: Madison, WI, USA, 1983; pp. 539–579. [Google Scholar]
- Olsen, S.R. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate; Miscellaneous Paper Institute for Agricultural Research Samaru, US Department of Agriculture: Washington, DC, USA, 1954. [Google Scholar]
- May, P.B.; Douglas, L.A. Assay for soil urease activity. Plant Soil 1976, 45, 301–305. [Google Scholar] [CrossRef]
- Gerritse, R.G.; Van Dijk, H. Determination of phosphatase activities of soils and animal wastes. Soil Biol. Biochem. 1978, 10, 545–551. [Google Scholar] [CrossRef]
- Guan, S. Soil Enzymes and Their Research Methodology; Agriculture: Beijing, China, 1986; pp. 274–338. [Google Scholar]
- Yan, G.; Han, S.; Wang, Q.; Wang, X.; Hu, C.; Xing, Y. Variations of the effects of reduced precipitation and N addition on microbial diversity among different seasons in a temperate forest. Appl. Soil Ecol. 2021, 166, 103995. [Google Scholar] [CrossRef]
- Chamizo, S.; Rodríguez-Caballero, E.; Román, J.R.; Cantón, Y. Effects of biocrust on soil erosion and organic carbon losses under natural rainfall. Catena 2017, 148, 117–125. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Morillas, L.; Maestre, F.T.; Gallardo, A. Biocrusts control the nitrogen dynamics and microbial functional diversity of semi-arid soils in response to nutrient additions. Plant Soil 2013, 372, 643–654. [Google Scholar] [CrossRef]
- Büdel, B.; Colesie, C.; Green, T.G.A.; Grube, M.; Suau, R.L.; Loewen-Schneider, K.; Maier, S.; Peer, T.; Pintado, A.; Raggio, J.; et al. Improved appreciation of the functioning and importance of biological soil crusts in Europe: The Soil Crust International Project (SCIN). Biodivers. Conserv. 2014, 23, 1639–1658. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Zhang, X. Microbial diversity analysis of different biological soil crusts in Tengger desert. Ecol. Sci. 2017, 36, 36–42. [Google Scholar]
- Zhang, Y.; Zhang, S.; Zhang, S.; Liu, Y.; Zhao, J.; Li, J. Effect of moss crust on sandy soil properties and bacterial community in Mu Us sand land. Acta Pedol. Sin. 2021, 58, 1585–1597. [Google Scholar]
- Zhang, T.; Jia, R.L.; Yu, L.Y. Diversity and distribution of soil fungal communities associated with biological soil crusts in the southeastern Tengger Desert (China) as revealed by 454 pyrosequencing. Fungal Ecol. 2016, 23, 156–163. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, Z.; Zhao, L.; Wang, X.; Liu, L.; Hui, R.; Zhang, W.; Zhang, P.; Song, G.; Sun, J. Differences in bacterial community structure between three types of biological soil crusts and soil below crusts from the Gurbantunggut Desert, China. Eur. J. Soil Sci. 2018, 70, 1–14. [Google Scholar] [CrossRef]
- Li, J.Y.; Jin, X.Y.; Zhang, X.C.; Chen, L.; Liu, J.L.; Zhang, H.M.; Zhang, X.; Zhang, Y.F.; Zhao, J.H.; Ma, Z.S.; et al. Comparative metagenomics of two distinct biological soil crusts in the Tengger Desert, China. Soil Biol. Biochem. 2020, 140, 107637. [Google Scholar] [CrossRef]
- Malam Issa, O.; Trichet, J.; Défarge, C.; Couté, A.; Valentin, C. Morphology and microstructure of microbiotic soil crusts on a tiger bush sequence (Niger, Sahel). Catena 1999, 37, 175–196. [Google Scholar] [CrossRef] [Green Version]
- Malam Issa, O.; Défarge, C.; Le Bissonnais, Y.; Marin, B.; Duval, O.; Bruand, A.; D’Acqui, L.P.; Nordenberg, S.; Annerman, M. Effects of the inoculation of cyanobacteria on the microstructure and the structural stability of a tropical soil. Plant Soil 2007, 290, 209–219. [Google Scholar] [CrossRef] [Green Version]
- Colesie, C.; Felde, V.J.M.; Büdel, B. Composition and macrostructure of biological soil crusts. In Biological Soil Crust: An Organizing Principle in Drylands; Springer: Cham, Switzerland, 2016; pp. 159–172. [Google Scholar]
- Seppelt, R.D.; Downing, A.J.; Deane-Coe, K.K.; Zhang, Y.; Zhang, J. Bryophytes within biological soil crusts. In Biological Soil Crust: An Organizing Principle in Drylands; Springer: Cham, Switzerland, 2016; pp. 101–120. [Google Scholar]
- Xiao, B.; Veste, M. Moss-dominated biocrusts increase soil microbial abundance and community diversity and improve soil fertility in semi-arid climates on the Loess Plateau of China. Appl. Soil Ecol. 2017, 117–118, 165–177. [Google Scholar] [CrossRef]
- He, M.; Hu, R.; Jia, R. Biological soil crusts enhance the recovery of nutrient levels of surface dune soil in arid desert regions. Ecol. Indic. 2019, 106, 105497. [Google Scholar] [CrossRef]
- Zhang, Y.; Gao, M.; Yu, C.; Zhang, H.; Yan, N.; Wu, Q.; Song, Y.; Li, X. Soil nutrients, enzyme activities, and microbial communities differ among biocrust types and soil layers in a degraded karst ecosystem. Catena 2022, 212, 106057. [Google Scholar] [CrossRef]
- Jin, X.; Zhang, X.; Jin, D.; Chen, Y.; Li, J. Diversity and seasonal dynamics of bacteria among different biological soil crusts in the southeast Tengger Desert. Biodiversity 2020, 28, 718–726. [Google Scholar] [CrossRef]
- Xu, H.J.; Wang, X.H.; Li, H.; Yao, H.Y.; Su, J.Q.; Zhu, Y.G. Biochar Impacts Soil Microbial Community Composition and Nitrogen Cycling in an Acidic Soil Planted with Rape. Environ. Sci. Technol. 2014, 48, 9391–9399. [Google Scholar] [CrossRef]
- Ding, Y.; Liu, Y.; Liu, S.; Li, Z.; Tan, X.; Huang, X.; Zeng, G.; Zhou, L.; Zheng, B. Biochar to improve soil fertility. A review. Agron. Sustain. Dev. 2016, 36, 36. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Liu, J.; Shen, J.; Chen, D.; Li, Y.; Jiang, B.; Wu, J. Effects of biochar amendment on net greenhouse gas emissions and soil fertility in a double rice cropping system: A 4-year field experiment. Agric. Ecosyst. Environ. 2018, 262, 83–96. [Google Scholar] [CrossRef]
- Liang, J.; Tang, S.; Gong, J.; Zeng, G.; Tang, W.; Song, B.; Zhang, P.; Yang, Z.; Luo, Y. Responses of enzymatic activity and microbial communities to biochar/compost amendment in sulfamethoxazole polluted wetland soil. J. Hazard. Mater. 2020, 385, 121533. [Google Scholar] [CrossRef]
- Zhang, L.; Jing, Y.; Xiang, Y.; Zhang, R.; Lu, H. Responses of soil microbial community structure changes and activities to biochar addition: A meta-analysis. Sci. Total Environ. 2018, 643, 926–935. [Google Scholar] [CrossRef]
- Taketani, R.G.; Tsai, S.M. The influence of different land uses on the structure of archaeal communities in Amazonian anthrosols based on 16S rRNA and amoA genes. Microb. Ecol. 2010, 59, 734–743. [Google Scholar] [CrossRef]
- Chen, J.; Liu, X.; Zheng, J.; Zhang, B.; Lu, H.; Chi, Z.; Pan, G.; Li, L.; Zheng, J.; Zhang, X. Biochar soil amendment increased bacterial but decreased fungal gene abundance with shifts in community structure in a slight acid rice paddy form Southwest China. Appl. Soil Ecol. 2013, 71, 33–34. [Google Scholar] [CrossRef]
- Killham, K.; Amato, M.; Ladd, J.N. Effect of substrate location in soil of and soil pore-water regime on carbon turnover. Soil Biol. Biochem. 1993, 25, 57–62. [Google Scholar] [CrossRef]
- Wright, D.A.; Killham, K.; Glover, L.A.; Prosser, J.I. Role of pore size location in determining bacteria activity during predation by protozoa in soil. Appl. Environ. Microbiol. 1995, 61, 3537–3543. [Google Scholar] [CrossRef] [Green Version]
- Young, I.M.; Ritz, K. The habitat of soil microbes. In Biological Diversity and Function in Soils; Cambridge University Press: Cambridge, UK, 2005; pp. 31–34. [Google Scholar]
- Davinic, M.; Fultz, L.M.; Acosta-Martinez, V.; Calderón, F.J.; Cox, S.B.; Dowd, S.E.; Allen, V.G.; Zak, J.C.; Moore-Kucera, J. Pyrosequencing and mid-infrared spectroscopy reveal distinct aggregate stratification of soil bacterial communities and organic matter composition. Soil Biol. Biochem. 2012, 46, 63–72. [Google Scholar] [CrossRef]
- Carly, S.; Cecilia, D.; Cassandre, G.; Edu, D.; Nancy, D.; David, G.; Albert, B.; Didier, A.; Roland, B.; David, F.; et al. Changes in species composition of European acid grasslands observed along a gradient of nitrogen deposition. J. Veg. Sci. 2011, 22, 207–215. [Google Scholar]
- Bandick, A.K.; Dick, R.P. Field management effffects on soil enzyme activities. Soil Biol. Biochem. 1999, 31, 1471–1479. [Google Scholar] [CrossRef]
- Qin, S.; Dong, W.; Hu, C. Tillage affffecting the turnover rate of soil urease: Implications for enzyme assays and ecological modeling. Fresenius Environ. Bull. 2010, 19, 717–720. [Google Scholar]
- Waldrop, M.P.; Zak, D.R.; Sinsabaugh, R.L.; Gallo, M.; Lauber, C. Nitrogen deposition modififies soil carbon storage through changes in microbial enzymatic activity. Ecol. Appl. 2004, 14, 1172–1177. [Google Scholar] [CrossRef]
- Ge, C.; Xue, D.; Yao, H. Microbial biomass, community diversity, and enzyme activities in response to urea application in tea orchard soils. Commun. Soil Sci. Plant Anal. 2010, 41, 797–810. [Google Scholar] [CrossRef]
- Bowles, T.M.; Acosta-Martínez, V.; Calderón, F.; Jackson, L. Soil enzyme activities, microbial communities, and carbon and nitrogen availability in organic agroecosystems across an intensively-managed agricultural landscape. Soil Biol. Biochem. 2014, 68, 252–262. [Google Scholar] [CrossRef]
- Elfstrand, S.; Hedlund, K.; Mårtensson, A. Soil enzyme activities, microbial community composition and function after 47 years of continuous green manuring. Appl. Soil Ecol. 2007, 35, 610–621. [Google Scholar] [CrossRef]
- Liu, Z.; Rong, Q.; Zhou, W.; Liang, G. Effects of inorganic and organic amendment on soil chemical properties, enzyme activities, microbial community and soil quality in yellow clayey soil. PLoS ONE 2017, 12, e0172767. [Google Scholar] [CrossRef] [PubMed]
- DeBruyn, J.M.; Nixon, L.T.; Fawaz, M.N.; Johnson, A.M.; Radosevich, M. Global biogeography and quantitative seasonal dynamics of Gemmatimonadetes in soil. Appl. Environ. Microbiol. 2011, 77, 6295–6300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, J.D.; Johnson, R.; Lehmann, J.; Olk, D.; Neves, E.; Thompson, M.; Schmidt-Rohr, K. Abundant and stable char residues in soils: Implications for soil fertility and carbon sequestration. Environ. Sci. Technol. 2012, 46, 9571–9576. [Google Scholar] [CrossRef]
- Cheng, Y.; Liu, Z.; Shen, Q.; Yang, X.; Xiao, X.; Zhang, T. The impact of straw biochar on corn rhizospheric and non-rhizospheric soil microbial community structure. Ecol. Environ. Sci. 2018, 27, 1870–1877. [Google Scholar]
- Liu, L.; Liu, Y.; Hui, R.; Xie, M. Recovery of microbial community structure of biological soil crusts in successional stage of Shapotou desert revegetation, northwest China. Soil Biol. Biochem. 2017, 107, 125–128. [Google Scholar] [CrossRef]
- Liu, L.; Liu, Y.; Zhang, P.; Song, G.; Hui, R.; Wang, Z.; Wang, J. Development of bacterial communities in biological soil crusts along a revegetation chronosequence in the Tengger Desert, northwest China. Biogeosciences 2017, 14, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Maier, S.; Schmidt, T.S.B.; Zheng, L.; Peer, T.; Wagner, V.; Grube, M. Analyses of dryland biological soil crusts highlight lichens as an important regulator of microbial communities. Biodivers. Conserv. 2014, 23, 1735–1755. [Google Scholar] [CrossRef]
- Wang, J.; Bao, J.; Su, J.; Li, X.; Chen, G.; Ma, X. Impact of inorganic nitrogen additions on microbes in biological soil crusts. Soil Biol. Biochem. 2015, 88, 303–313. [Google Scholar] [CrossRef]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef] [PubMed]
- Xun, W.; Xiong, W.; Huang, T.; Ran, W.; Li, D.; Shen, Q.; Li, Q.; Zhang, R. Swine manure and quicklime have different impacts on chemical properties and composition of bacterial communities of an acidic soil. Appl. Soil Ecol. 2016, 100, 38–44. [Google Scholar] [CrossRef]
- Häni, H.; Siegenthaler, A.; Candinas, T. Soil effects due to sewage sludge application in agriculture. Fertil. Res. 1995, 43, 149–156. [Google Scholar] [CrossRef]
- Gao, L.; Wang, R.; Shen, G.; Zhang, J.; Meng, G.; Zhang, J. Effects of biochar on nutrients and the microbial community structure of tobacco-planting soils. J. Soil Sci. Plant Nut. 2017, 17, 884–896. [Google Scholar] [CrossRef] [Green Version]
- Lopes, A.R.; Manaia, C.M.; Nunes, O.C. Bacterial community variations in an alfalfa-rice rotation system revealed by 16S rRNA gene 454-pyrosequencing. FEMS Microbiol. Ecol. 2014, 87, 650–663. [Google Scholar] [CrossRef] [Green Version]
- Rousk, J.; Brookes, P.C.; Bååth, E. Fungal and bacterial growth responses to N fertilization and pH in the 150-year ‘Park Grass’ UK grassland experiment. FEMS Microbiol. Ecol. 2011, 76, 89–99. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Liu, W.; Zhu, C.; Luo, G.; Kong, Y.; Ling, N.; Wang, M.; Dai, J.; Shen, Q.; Guo, S. Bacterial rather than fungal community composition is associated with microbial activities and nutrient-use efficiencies in a paddy soil with short-term organic amendments. Plant Soil 2018, 424, 335–349. [Google Scholar] [CrossRef]
- Zhu, M.; Liu, X.; Wang, J.; Liu, Z.; Pan, G. Effects of biochar application on soil microbial diversity in soil aggregates from paddy soil. Acta Ecol. Sin. 2020, 40, 1505–1516. [Google Scholar]
- Li, F.; Li, M.; Liu, J.; Hu, Y.; Zhang, Q.; Zhao, H. Effect of biochar on fungal abundance of rhizosphere soil and cucumber root growth in greenhouse. Trans. Chin. Soc. Agric. Mach. 2017, 48, 265–270. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | Treatment | pH | OM (g·kg−1) | TN (g·kg−1) | TP (g·kg−1) | TK (g·kg−1) | UE (U·g−1) | ACP (U·g−1) | SC (U·g−1) | PPO (U·g−1) |
|---|---|---|---|---|---|---|---|---|---|---|
| In BSCs | CK | 3.73 c | 13.16 b | 1.53 b | 0.317 a | 6.26 bc | 499.04 a | 47660.07 a | 31.31 b | 40.76 a |
| Biochar | 4.27 a | 26.29 a | 2.33 a | 0.207 b | 7.51 a | 607.77 a | 46500.84 a | 64.20 a | 39.51 a | |
| Beneath BSCs | CK | 3.97 b | 6.48 d | 0.74 c | 0.287 ab | 5.80 c | 178.82 b | 49020.77 a | 17.88 b | 35.90 ab |
| Biochar | 4.23 a | 9.08 c | 1.29 b | 0.280 ab | 6.70 b | 193.03 b | 41211.91 b | 22.26 b | 24.06 b |
| Microbes | Position | Treatment | Kingdom | Phylum | Class | Order | Family | Genus | Species |
|---|---|---|---|---|---|---|---|---|---|
| Bacteria | In BSCs | CK | 1 | 17 | 40 | 88 | 127 | 190 | 204 |
| Biochar | 1 | 17 | 42 | 94 | 135 | 194 | 205 | ||
| Beneath BSCs | CK | 1 | 17 | 43 | 93 | 137 | 201 | 217 | |
| Biochar | 1 | 18 | 44 | 97 | 139 | 212 | 226 | ||
| Fungi | In BSCs | CK | 1 | 7 | 21 | 52 | 78 | 107 | 85 |
| Biochar | 1 | 7 | 20 | 53 | 75 | 102 | 80 | ||
| Beneath BSCs | CK | 1 | 6 | 20 | 45 | 64 | 82 | 65 | |
| Biochar | 1 | 6 | 16 | 40 | 54 | 59 | 44 |
| Microbes | Position | Treatment | OTU Number | ACE | Chao1 | Simpson | Shannon |
|---|---|---|---|---|---|---|---|
| Bacteria | In BSCs | CK | 677 ± 78 a | 751.2 ± 58.9 a | 752.0 ± 63.3 a | 0.985 ± 0.002 a | 7.471 ± 0.163 a |
| Biochar | 657 ± 114 a | 753.6 ± 78.7 a | 764.8 ± 85.1 a | 0.984 ± 0.003 a | 7.432 ± 0.144 a | ||
| Beneath BSCs | CK | 343 ± 25 b | 414.2 ± 27 b | 409.9 ± 24.8 b | 0.976 ± 0.005 a | 6.675 ± 0.209 a | |
| Biochar | 512 ± 163 a | 547.7 ± 149 ab | 560.0 ± 146.7 ab | 0.976 ± 0.008 a | 6.913 ± 0.438 a | ||
| Fungi | In BSCs | CK | 409 ± 56 a | 453.0 ± 47.7 a | 463.3 ± 52.4 a | 0.834 ± 0.043 a | 4.541 ± 0.520 a |
| Biochar | 385 ± 24 a | 441.0 ± 14.8 a | 442.0 ± 22.1 ab | 0.756 ± 0.041 a | 4.074 ± 0.222 a | ||
| Beneath BSCs | CK | 230 ± 38 b | 346.2 ± 43.2 ab | 323.8 ± 38.3 bc | 0.899 ± 0.030 a | 4.856 ± 0.347 a | |
| Biochar | 143 ± 7 b | 275.6 ± 44.4 b | 216.4 ± 23.9 c | 0.788 ± 0.111 a | 4.072 ± 0.806 a |
| Microbes | Name | Explains % | Contribution % | pseudo-F | P |
|---|---|---|---|---|---|
| Bacteria | UE | 38.8 | 78.7 | 6.3 | 0.006 |
| TN | 3.5 | 7.2 | 0.6 | 0.602 | |
| OM | 6.9 | 14.1 | 1.1 | 0.314 | |
| Fungi | UE | 50.2 | 76.8 | 10.1 | 0.008 |
| pH | 11.4 | 17.4 | 2.7 | 0.102 | |
| OM | 2.1 | 3.2 | 0.5 | 0.686 | |
| SC | 1.8 | 2.7 | 0.4 | 0.734 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Huang, R.; Zhu, L.; Guan, H.; Lin, L.; Fang, H.; Yang, M.; Ji, S.; Zou, X.; Li, X. The Effects of Biochar on Microbial Community Composition in and Beneath Biological Soil Crusts in a Pinus massoniana Lamb. Plantation. Forests 2022, 13, 1141. https://doi.org/10.3390/f13071141
Wang J, Huang R, Zhu L, Guan H, Lin L, Fang H, Yang M, Ji S, Zou X, Li X. The Effects of Biochar on Microbial Community Composition in and Beneath Biological Soil Crusts in a Pinus massoniana Lamb. Plantation. Forests. 2022; 13(7):1141. https://doi.org/10.3390/f13071141
Chicago/Turabian StyleWang, Jinping, Rongzhen Huang, Liqin Zhu, Hongzhi Guan, Lijing Lin, Huanying Fang, Mengjia Yang, Shaohui Ji, Xianhua Zou, and Xin Li. 2022. "The Effects of Biochar on Microbial Community Composition in and Beneath Biological Soil Crusts in a Pinus massoniana Lamb. Plantation" Forests 13, no. 7: 1141. https://doi.org/10.3390/f13071141
APA StyleWang, J., Huang, R., Zhu, L., Guan, H., Lin, L., Fang, H., Yang, M., Ji, S., Zou, X., & Li, X. (2022). The Effects of Biochar on Microbial Community Composition in and Beneath Biological Soil Crusts in a Pinus massoniana Lamb. Plantation. Forests, 13(7), 1141. https://doi.org/10.3390/f13071141