
Long-Term Carbon Sequestration in Pine Forests under Different Silvicultural and Climatic Regimes in Spain

 ,
,  ,
,  , ,
, ,  , ,
, ,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
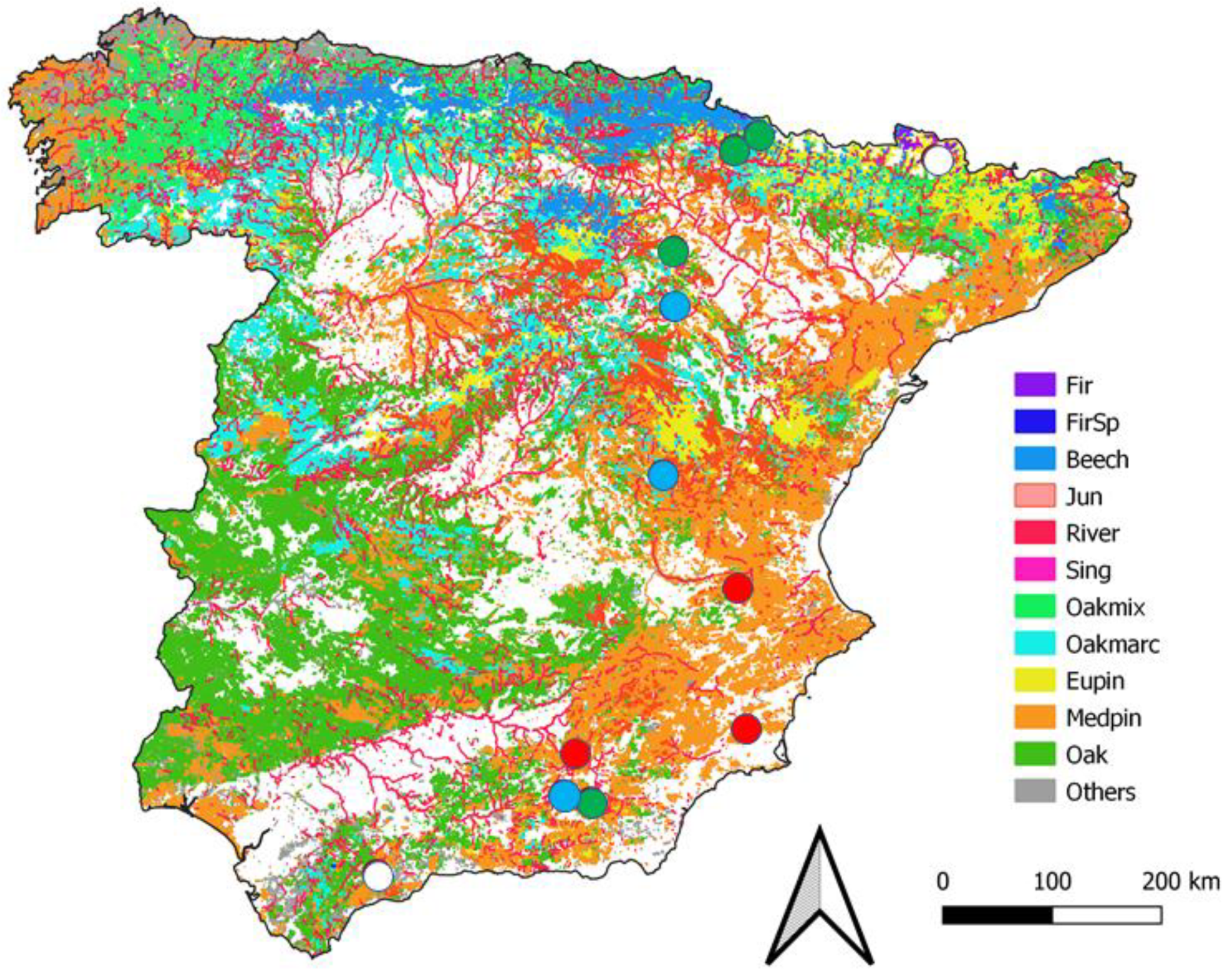
2.1. Study Area
2.2. Experimental Design and Field Data
2.3. Growth and Biomass Estimation
2.4. Soils Samples and SOC Estimation
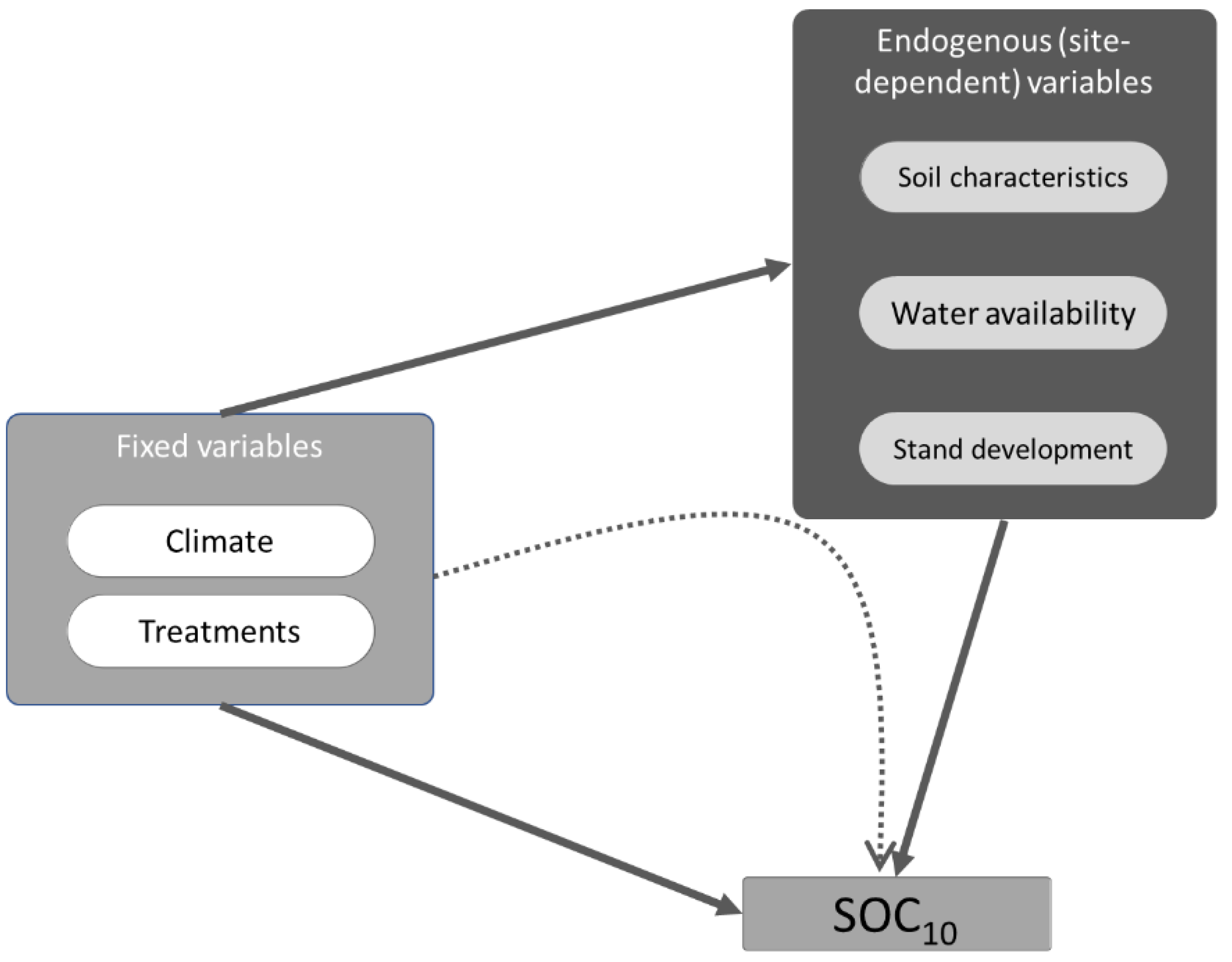
2.5. Environmental Variables
2.6. Statistical Analysis
3. Results
3.1. Biomass and Radial Growth Responses to Thinning
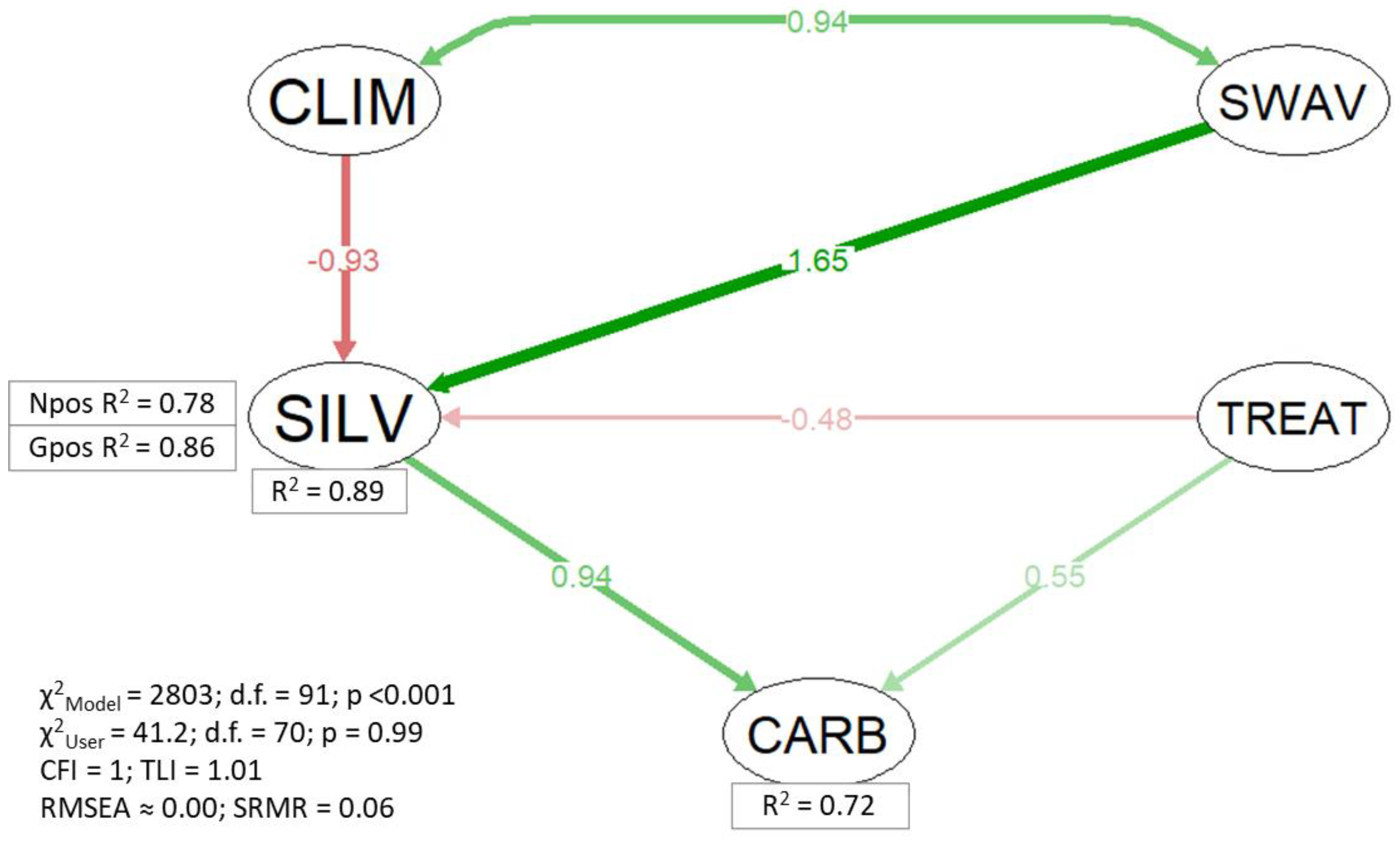
3.2. Structural Equations Model
4. Discussion
4.1. Effects of Thinning on Radial Growth
4.2. Thinning as a Tool to Optimize C Sequestration
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC—International Panel of Climate Change. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. In Climate Change; IPCC—International Panel of Climate Change, Pachauri, R.K., Meyer, L.A., Eds.; Synthesis Report; IPCC: Geneva, Switzerland, 2014; p. 151. [Google Scholar]
- Olivier, J.G.J.; Peters, J.A.H.W. Trends in Global CO2 and Total Greenhouse Gas Emissions: Report 2019; PBL Netherlands Environmental Assessment Agency: The Hague, The Netherlands, 2020; p. 70. [Google Scholar]
- Six, J.; Callewaert, P.; Lenders, S.; De Gryze, S.; Morris, S.J.; Gregorich, E.G.; Paustian, K. Measuring and understanding carbon storage in afforested soils by physical fractionation. Soil Sci. Soc. Am. J. 2002, 66, 1981–1987. [Google Scholar] [CrossRef] [Green Version]
- Alemu, B. The role of forest and soil carbon sequestrations on climate change mitigation. Res. J. Agric. Environ. Manag. 2014, 3, 492–505. [Google Scholar]
- Pan, L.; Oldenburg, C.M.; Pruess, K.; Wu, Y.S. Transient CO2 leakage and injection in wellbore-reservoir systems for geologic carbon sequestration. Greenh. Gases Sci. Technol. 2011, 1, 335–350. [Google Scholar] [CrossRef]
- Domke, G.M.; Oswalt, S.N.; Walters, B.F.; Morin, R.S. Tree planting has the potential to increase carbon sequestration capacity of forests in the United States. Proc. Natl. Acad. Sci. USA 2020, 117, 24649–24651. [Google Scholar] [CrossRef] [PubMed]
- Bárcena, T.G.; Gundersen, P.; Vesterdal, L. Afforestation effects on SOC in former cropland: Oak and spruce chronosequences resampled after 13 years. Glob. Change Biol. 2014, 20, 2938–2952. [Google Scholar] [CrossRef]
- Palmero-Iniesta, M.; Pino, J.; Pesquer, L.; Espelta, J.M. Recent forest area increases in Europe: Expanding and regenerating forests differ in their regional patterns, drivers and productivity trends. Eur. J. For. Res. 2021, 140, 793–805. [Google Scholar] [CrossRef]
- Paquette, A.; Messier, C. The role of plantations in managing world’s forests in the Anthropocene. Front. Ecol. Environ. 2010, 8, 27–34. [Google Scholar] [CrossRef] [Green Version]
- Ciais, P.; Schelhaas, M.J.; Zaehle, S.; Piao, S.L.; Cescatti, A.; Liski, J.; Nabuurs, G.J. Carbon accumulation in European forests. Nat. Geosci. 2008, 1, 425–429. [Google Scholar] [CrossRef]
- Agyei, T.; Juráň, S.; Ofori-Amanfo, K.K.; Šigut, L.; Urban, O.; Marek, M.V. The impact of drought on total ozone flux in a mountain Norway spruce forest. J. For. Sci. 2020, 66, 278–280. [Google Scholar] [CrossRef]
- Solberg, S.; Dobbertin, M.; Reinds, G.J.; Lange, H.; Andreassen, K.; Fernandez, P.G.; de Vries, W. Analyses of the impact of changes in atmospheric deposition and climate on forest growth in European monitoring plots: A stand growth approach. For. Ecol. Manag. 2009, 258, 1735–1750. [Google Scholar] [CrossRef]
- Devi, N.M.; Kukarskih, V.V.; Galimova, A.A.; Mazepa, V.S.; Grigoriev, A.A. Climate change evidence in tree growth and stand productivity at the upper treeline ecotone in the Polar Ural Mountains. For. Ecosyst. 2020, 7, 7. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y. Integrate carbon dynamic models in analyzing carbon sequestration impact of forest biomass harvest. Sci. Total Environ. 2018, 615, 581–587. [Google Scholar] [CrossRef]
- Valade, A.; Bellassen, V.; Magand, C.; Luyssaert, S. Sustaining the sequestration efficiency of the European forest sector. For. Ecol. Manag. 2017, 405, 44–55. [Google Scholar] [CrossRef]
- Jandl, R.; Lindner, M.; Vesterdal, L.; Bauwens, B.; Baritz, R.; Hagedorn, F.; Johnson, D.; Minkkinen, K.; Byrne, K. How strongly can forest management influence soil carbon sequestration? Geoderma. 2007, 15, 253–268. [Google Scholar] [CrossRef]
- Blanco, J.A.; Imbert, J.B.; Castillo, F.J. Adaptación al Cambio Climático en Pinares Pirenaicos: Controlando la Densidad del Rodal Según el Tipo de Clima. In Los Bosques y la Biodiversidad Frente al Cambio Climático: Impactos, Vulnerabilidad y Adaptación en España; Ministerio de Medio Ambiente: Madrid, Spain, 2015; pp. 565–572. [Google Scholar]
- Lado-Monserrat, L.; Lidón, A.; Bautista, I. Erratum to: Litterfall, litter decomposition and associated nutrient fluxes in Pinus halepensis: Influence of tree removal intensity in a Mediterranean forest. Eur. J. For. Res. 2016, 135, 203–214. [Google Scholar] [CrossRef] [Green Version]
- Fares, S.; Scarasia Mugnozza, G.; Corona, P.; Palahí, M. Sustainability: Five steps for managing Europe’s forests. Nat. News 2015, 519, 407. [Google Scholar] [CrossRef] [PubMed]
- Bombín, R.V. El mapa forestal de España escala 1: 50.000 (MFE50) como base del tercer inventario forestal nacional. Cuad. Soc. Española Cien. For. 2005, 19, 205–210. [Google Scholar]
- Vadell, E.; de-Miguel, S.; Pemán, J. Large-scale reforestation and afforestation policy in Spain: A historical review of its underlying ecological, socioeconomic and political dynamics. Land Use Policy 2016, 55, 37–48. [Google Scholar] [CrossRef]
- Alfaro-Sánchez, R.; Camarero, J.J.; López-Serrano, F.R.; Sánchez-Salguero, R.; Moya, D.; De Las Heras, J. Positive coupling between growth and reproduction in young post-fire Aleppo pines depends on climate and site conditions. Int. J. Wildland Fire 2015, 24, 507–517. [Google Scholar] [CrossRef] [Green Version]
- Navarro-Cerrillo, R.M.; Sánchez-Salguero, R.; Rodríguez, C.; Lazo, J.; Moreno-Rojas, J.; Palacios-Rodríguez, G.; Camarero, J.J. Is thinning an alternative when trees could die in response to drought? The case of planted Pinus nigra and P. sylvestris stands in southern Spain. For. Ecol. Manag. 2019, 433, 313–324. [Google Scholar] [CrossRef]
- Manrique-Alba, À.; Beguería, S.; Molina, A.J.; González-Sanchis, M.; Tomàs-Burguera, M.; Del Campo, A.D.; Camarero, J.J. Long-term thinning effects on tree growth, drought response and water use efficiency at two Aleppo pine plantations in Spain. Sci. Total Environ. 2020, 728, 138536. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Peinado, R.; Bravo-Oviedo, A.; López-Senespleda, E.; Montero, G.; Río, M. Do thinning influence biomass and soil carbon stocks in Mediterranean maritime pinewoods? Eur. J. For. Res. 2013, 132, 253–262. [Google Scholar] [CrossRef]
- Ruiz-Peinado, R.; Bravo-Oviedo, A.; Montero, G.; Del Río, M. Carbon stocks in a Scots pine afforestation under different thinning intensities management. Mitig. Adapt. Strateg. Glob. Change 2016, 21, 1059–1072. [Google Scholar] [CrossRef]
- Pregitzer, K.S.; Euskirchen, E.S. Carbon cycling and storage in world forests: Biome patterns related to forest age. Glob. Change Biol. 2004, 10, 2052–2077. [Google Scholar] [CrossRef]
- Zhang, X.; Guan, D.; Li, W.; Sun, D.; Jin, C.; Yuan, F.; Wu, J. The effects of forest thinning on soil carbon stocks and dynamics: A meta-analysis. For. Ecol. Manag. 2018, 429, 36–43. [Google Scholar] [CrossRef]
- Lal, R.; Smith, P.; Jungkunst, H.F.; Mitsch, W.J.; Lehmann, J.; Nair, P.R.; Ravindranath, N.H. The carbon sequestration potential of terrestrial ecosystems. J. Soil Water Conserv. 2018, 73, 145A–152A. [Google Scholar] [CrossRef] [Green Version]
- Krug, J.H. How can forest management increase biomass accumulation and CO2 sequestration? A case study on beech forests in Hesse, Germany. Carbon Balance Manag. 2019, 14, 17. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.W.; Curtis, P.S. Effects of forest management on soil C and N storage: Meta-analysis. For. Ecol. Manag. 2001, 140, 227–238. [Google Scholar] [CrossRef]
- Lal, R. Forest soils and carbon sequestration. For. Ecol. Manag. 2005, 220, 242–258. [Google Scholar] [CrossRef]
- Lal, R. Managing soils and ecosystems for mitigating anthropogenic carbon emissions and advancing global food security. BioScience 2010, 60, 108–721. [Google Scholar] [CrossRef]
- Prescott, C.E. Litter decomposition: What controls it and how can we alter it to sequester more carbon in forest soils? Biogeochemistry 2010, 101, 133–149. [Google Scholar] [CrossRef]
- Nave, L.; Vance, E.; Swanston, C.; Curtis, P. Harvest impacts on soil carbon storage in temperate forests. For. Ecol. Manag. 2010, 259, 857–866. [Google Scholar] [CrossRef]
- Blanco, J.A. Managing Forest soils for carbon sequestration: Insights from modeling forests around the globe. In Soil Management and Climate Change; Muñoz, M.Á., Zornoza, R., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2018; pp. 237–252. [Google Scholar]
- Noormets, A.; Epron, D.; Domec, J.C.; McNulty, S.G.; Fox, T.; Sun, G.; King, J.S. Effects of forest management on productivity and carbon sequestration: A review and hypothesis. For. Ecol. Manag. 2015, 355, 124–140. [Google Scholar] [CrossRef] [Green Version]
- Ontl, T.A.; Janowiak, M.K.; Swanston, C.W.; Daley, J.; Handler, S.; Cornett, M.; Patch, N. Forest management for carbon sequestration and climate adaptation. J. For. 2020, 118, 86–101. [Google Scholar] [CrossRef] [Green Version]
- Tong, X.; Brandt, M.; Yue, Y.; Ciais, P.; Jepsen, M.R.; Penuelas, J.; Fensholt, R. Forest management in southern China generates short term extensive carbon sequestration. Nat. Commun. 2020, 11, 129. [Google Scholar] [CrossRef] [PubMed]
- Körner, C. A matter of tree longevity. Science 2017, 355, 130–131. [Google Scholar] [CrossRef] [PubMed]
- Serrada, R. (Ed.) Compendio de Selvicultura Aplicada en España; Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria, Ministerio de Educación y Ciencia: Madrid, Spain, 2008.
- Molina, A.J.; Navarro-Cerrillo, R.M.; Pérez-Romero, J.; Alejano, R.; Bellot, J.F.; Blanco, J.A.; Camarero, J.J.; Carrara, A.; Castillo, V.M.; Cervera, T.; et al. SilvAdapt. Net: A Site-Based Network of Adaptive Forest Management Related to Climate Change in Spain. Forests 2021, 12, 1807. [Google Scholar] [CrossRef]
- Collalti, A.; Trotta, C.; Keenan, T.F.; Ibrom, A.; Bond-Lamberty, B.; Grote, R.; Vicca, S.; Reyer, C.P.O.; Migliavacca, M.; Veroustraete, F.; et al. Thinning can reduce losses in carbon use efficiency and carbon stocks in managed forests under warmer climate. J. Adv. Model. Earth Syst. 2018, 10, 2427–2452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Peinado, R.; Oviedo, J.A.B.; Senespleda, E.L.; Oviedo, F.B.; del Río Gaztelurrutia, M. Forest management and carbon sequestration in the Mediterranean region: A review. For. Syst. 2017, 26, 10. [Google Scholar] [CrossRef]
- Ruiz-Peinado, R.; del Rio, M. New models for estimating the carbon sink capacity of Spanish softwood species. For. Syst. 2011, 20, 176–188. [Google Scholar] [CrossRef] [Green Version]
- Nelson, D.W.; Sommers, L.E. Total Carbon, Organic Carbon, and Organic Matter. Methods Soil Anal. Part 3 Chem. Methods 1996, 3, 961–1010. [Google Scholar]
- Post, W.M.; Kwon, K.C. Soil Carbon Sequestration and Land-Use Change: Processes and Potential. Glob. Change Boil. 2000, 6, 317–327. [Google Scholar] [CrossRef] [Green Version]
- Farina, R.; Marchetti, A.; Francaviglia, R.; Napoli, R.; di Bene, C. Modeling Regional Soil C Stocks and CO2 Emissions under Mediterranean Cropping Systems and Soil Types. Agric. Ecosyst. Environ. 2017, 238, 128–141. [Google Scholar] [CrossRef]
- Felicísimo, A.M. Impactos, Vulnerabilidad y Adaptación al Cambio Climático de la Biodiversidad Española. 1. Flora y Vegetación; Oficina Española de Cambio Climático, Ministerio de Medio Ambiente y Medio Rural y Marino: Madrid, Spain, 2011.
- De Cáceres, M.; Martin, N.; Granda, V.; Cabon, A. Meteoland: Landscape Meteorology Tools. R package Version 0.7.1. 2018. Available online: https://cran.r-project.org/web/packages/meteoland/meteoland.pdf (accessed on 21 September 2021).
- Esri ArcGIS Resources Online. Geoprocessing Tool Reference 2011. Available online: https://desktop.arcgis.com/es/arcmap/latest/tools/main/a-quick-tour-of-geoprocessing-tool-references.htm (accessed on 21 September 2021).
- Zuur, A.; Ieno, E.; Walker, N.; Saveliev, A.; Smith, G. Mixed Effects Models and Extensions in Ecology with R; Springer: New York, NY, USA, 2009; pp. 246–250. [Google Scholar]
- Sokal, R.R.; Rohlf, F.J. Biometry: The Principles and Practices of Statistics in Biological Research; W. H. Freeman and Company: New York, NY, USA, 1995. [Google Scholar]
- Mueller, R.O.; Hancock, G.R. Structural Equation Modeling; Routledge: New York, NY, USA; Taylor & Francis Group: Abingdon, UK, 2019. [Google Scholar]
- Finch, W.H.; French, B.F. Latent Variable Modeling with R; Routledge: New York, NY, USA, 2015. [Google Scholar]
- Rosseel, Y. Lavaan: An R package for structural equation modeling and more. Version 0.5–12 (BETA). J. Stat. Softw. 2012, 48, 1–36. [Google Scholar] [CrossRef] [Green Version]
- Epskamp, S.; Epskamp, M.S. MplusAutomation, Package ‘semPlot’. R Package Version, 1. 2019. Available online: https://www.scirp.org/(S(i43dyn45teexjx455qlt3d2q))/reference/referencespapers.aspx?referenceid=3012616 (accessed on 21 September 2021).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Martín-Benito, D.; Del Río, M.; Heinrich, I.; Helle, G.; Cañellas, I. Response of climate-growth relationships and water use efficiency to thinning in a Pinus nigra afforestation. For. Ecol. Manag. 2010, 259, 967–975. [Google Scholar] [CrossRef]
- Pérez-de-Lis, G.; García-González, I.; Rozas, V.; Arévalo, J.R. Effects of thinning intensity on radial growth patterns and temperature sensitivity in Pinus canariensis afforestations on Tenerife Island, Spain. Ann. For. Sci. 2011, 68, 1093–1104. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Gutiérrez, C.; Dawson, T.E.; Nicolás, E.; Querejeta, J.I. Isotopes reveal contrasting water use strategies among coexisting plant species in a Mediterranean ecosystem. New Phytol. 2012, 196, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Cerrillo, R.M.; Duque-Lazo, J.; Rodríguez-Vallejo, C.; Varo-Martínez, M.; Palacios-Rodríguez, G. Airborne laser scanning cartography of on-site carbon stocks as a basis for the silviculture of Pinus halepensis plantations. Remote Sens. 2018, 10, 1660. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, T.J.; Del Campo, A.D.; Herrera, R.; Molina, A.J. Simultaneous assessment, through sap flow and stable isotopes, of water use efficiency (WUE) in thinned pines shows improvement in growth, tree-climate sensitivity and WUE, but not in WUEi. For. Ecol. Manag. 2016, 361, 298–308. [Google Scholar] [CrossRef]
- del Río, M.; Barbeito, I.; Bravo-Oviedo, A.; Calama, R.; Cañellas, I.; Herrero, C.; Bravo, F. Mediterranean pine forests: Management effects on carbon stocks. In Managing Forest Ecosystems: The Challenge of Climate Change; Springer: Cham, Switzerland, 2017; pp. 301–327. [Google Scholar]
- Jiménez, M.N.; Navarro, F.B.; Sánchez-Miranda, A.; Ripoll, M.A. Using stem diameter variations to detect and quantify growth and relationships with climatic variables on a gradient of thinned Aleppo pines. For. Ecol. Manag. 2019, 442, 53–62. [Google Scholar] [CrossRef]
- Alvarez, S.; Ortiz, C.; Díaz-Pinés, E.; Rubio, A. Influence of tree species composition, thinning intensity and climate change on carbon sequestration in Mediterranean mountain forests: A case study using the CO2Fix model. Mitig. Adapt. Strateg. Glob. Change 2016, 21, 1045–1058. [Google Scholar] [CrossRef]
- Mäkinen, H.; Isomäki, A. Thinning intensity and growth of Norway spruce stands in Finland. Forestry 2004, 77, 349–364. [Google Scholar] [CrossRef] [Green Version]
- Del Campo, A.D.; González-Sanchis, M.; Molina, A.J.; García-Prats, A.; Ceacero, C.J.; Bautista, I. Effectiveness of water-oriented thinning in two semiarid forests: The redistribution of increased net rainfall into soil water, drainage and runoff. For. Ecol. Manag. 2019, 438, 163–175. [Google Scholar] [CrossRef]
- Sohn, J.A.; Saha, S.; Bauhus, J. Potential of forest thinning to mitigate drought stress: A meta-analysis. For. Ecol. Manag. 2016, 380, 261–273. [Google Scholar] [CrossRef]
- González de Andrés, E.; Camarero, J.J.; Blanco, J.A.; Imbert, J.B.; Lo, Y.H.; Sangüesa-Barreda, G.; Castillo, F.J. Tree-to-tree competition in mixed European beech-Scots pine forests has different impacts on growth and water-use efficiency depending on site condition. J. Ecol. 2018, 106, 59–75. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Salguero, R.; Navarro-Cerrillo, R.M.; Swetnam, T.W.; Zavala, M.A. Is drought the main decline factor at the rear edge of Europe? The case of southern Iberian pine plantations. For. Ecol. Manag. 2012, 271, 158–169. [Google Scholar] [CrossRef]
- Kaarakka, L.; Cornett, M.; Domke, G.; Ontl, T.; Dee, L.E. Improved forest management as a natural climate solution: A review. Ecol. Solut. Evid. 2021, 2, e12090. [Google Scholar] [CrossRef]
- Navarro-Cerrillo, R.M.; Rodriguez-Vallejo, C.; Silveiro, E.; Hortal, A.; Palacios-Rodríguez, G.; Duque-Lazo, J.; Camarero, J.J. Cumulative drought stress leads to a loss of growth resilience and explains higher mortality in planted than in naturally regenerated Pinus pinaster stands. Forests 2018, 9, 358. [Google Scholar] [CrossRef] [Green Version]
- Bradley, R.L.; Kimmins, J.P.; Martin, W.L. Post-clearcutting chronosequences in the B.C. Coastal Western Hemlock Zone: II. Tracking the assart flush. J. Sustain. For. 2002, 14, 23–43. [Google Scholar] [CrossRef]
- Matsushima, M.; Chang, S.X. Effects of understory removal, N fertilization, and litter layer removal on soil N cycling in a 13-year-old white spruce plantation infested with Canada bluejoint grass. Plant Soil 2007, 292, 243–258. [Google Scholar] [CrossRef]
- Hedo de Santiago, J.; Lucas-Borja, M.E.; Wic-Baena, C.; Andrés-Abellán, M.; de las Heras, J. Effects of thinning and induced drought on microbiological soil properties and plant species diversity at dry and semiarid locations. Land Degrad. Dev. 2016, 27, 1151–1162. [Google Scholar] [CrossRef]
- Seibt, U.; Rajabi, A.; Griffiths, H.; Berry, J.A. Carbon isotopes and water use efficiency: Sense and sensitivity. Oecologia 2008, 155, 441–454. [Google Scholar] [CrossRef] [PubMed]
- Vayreda, J.; Martinez-Vilalta, J.; Gracia, M.; Retana, J. Recent climate changes interact with stand structure and management to determine changes in tree carbon stocks in Spanish forests. Glob. Change Biol. 2012, 18, 1028–1041. [Google Scholar] [CrossRef]
- Fernandez-de-Una, L.; Cañellas, I.; Gea-Izquierdo, G. Stand competition determines how different tree species will cope with a warming climate. PLoS ONE 2015, 10, e0122255. [Google Scholar] [CrossRef] [Green Version]
- del Castillo, J.; Comas, C.; Voltas, J.; Ferrio, J.P. Dynamics of competition over water in a mixed oak-pine Mediterranean forest: Spatio-temporal and physiological components. For. Ecol. Manag. 2016, 382, 214–224. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Specie | Thinning | D | Dbh | G | Wa | Wt | SOC10 | SOC20 | SOC10–20 | BAI20 | BAI |
|---|---|---|---|---|---|---|---|---|---|---|---|
| P. halepensis | CT | 1477.30(102.59) | 14.44(1.63) | 24.90(3.92) a | 68.45(8.52) a | 94.54(22.86) | 11.79(1.02) c | 12.72(0.71) c | 24.52(1.33) c | 2.10(0.59) | 2.40(0.88) |
| LT | 754.30(40.63) | 16.63(1.90) | 16.07(2.80) ab | 46.21(9.98) b | 64.11(16.82) | 17.86(1.16) b | 18.10(1.88) b | 35.96(2.86) b | 3.60(0.92) | 3.73(0.98) | |
| HT | 557.30(47.66) | 17.44(2.08) | 12.03(1.94) b | 35.95(9.69) b | 49.99(12.52) | 27.30(3.63) a | 23.42(4.28) a | 50.72(7.85) a | 3.75(1.03) | 4.20(1.06) | |
| P. nigra | CT | 698.50(77.25) | 15.16(0.83) | 18.26(5.24) | 74.65(24.32) | 87.10(26.78) | 6.40(0.90) | 5.97(0.70) | 12.69(1.78) | 3.42(0.43) | |
| LT | 475.75(38.58) | 15.28(1.05) | 13.66(4.18) | 48.51(13.47) | 56.98(14.90) | 5.38(0.66) | 6.79(1.01) | 12.41(1.80) | 4.35(0.96) | 4.95(1.03) | |
| HT | 401.00(20.74) | 16.78(0.56) | 8.66(0.16) | 47.39(16.02) | 56.12(17.43) | 5.51(1.46) | 6.10(0.42) | 11.62(1.78) | 4.93(1.22) | 5.60(0.97) | |
| P. sylvestris | CT | 2666.11(498.38) | 16.06(1.02) | 46.21(9.13) | 171.48(31.00) | 192.40(17.54) | 31.75(5.13) | 15.52(5.83) | 29.89(9.19) | 6.04(0.90) | |
| LT | 1474.33(244.18) | 18.09(1.01) | 34.84(6.72) | 143.79(26.07) | 160.43(20.92) | 33.28(6.38) | 7.59(1.86) | 16.17(3.99) | 6.83(0.86) | 7.07(0.83) | |
| HT | 1053.89(168.48) | 17.81(1.02) | 25.92(5.95) | 112.18(22.24) | 143.51(20.55) | 30.83(5.02) | 17.09(7.42) | 31.01(12.65) | 6.55(0.60) | 7.29(0.66) |
| Variable | Description | Units |
|---|---|---|
| Soil water availability (SWAV) | ||
| wn | mean annual soil moisture | mm |
| SM90 | max. annual soil volumetric water content calculated as the 90 quantile of daily | % vol. |
| SM50 | wiegthed mean soil volumetric water content calculated as the 50 quantile of daily | % vol. |
| theta_s | saturated soil volumetric water content | % vol. |
| theta_r | saturated soil volumetric water content | % vol. |
| doy_stress | total number of days under soil moisture stress | days |
| Climatic characteristics (CLIM) | ||
| ea_n | annual actual evapotranspiration | mm |
| tair | Mean annual temperature (air) | °C |
| deficit | Hydric deficit | mm |
| Soil chemical and physical characteristics (SCHR) | ||
| pH | Unitless | |
| Bulk_density | Bulk soil density | g cm−3 |
| CEC | ||
| Silvicultural characteristics (SILV) | ||
| Npos | Density of the plot 10 years after thinning? | Trees ha−1 |
| GPos | Basal area of the plot 10 years after thinning | m2 ha−1 |
| Thinning (TREAT) | ||
| TR | Thinning factor (intensity) | Unitless |
| Intensity | Percentage of trees | % |
| Selected Variables | # Comp. | Cum. Var. |
|---|---|---|
| SWAV =~ wn + SM90 + SM50 + theta_s + theta_r + doy_stress | 1 | 90.61% |
| CLIM =~ ea_n + tair + déficit | 1 | 79.07% |
| SCHR =~ pH + Bulk_density + CEC | 1 | 89.72% |
| SILV =~ Npos + Gpos | 1 | 91.31% |
| TREAT =~ TR + Intensity | … | N/A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Navarro-Cerrillo, R.M.; Ruiz-Gómez, F.J.; Camarero, J.J.; Castillo, V.; Barberá, G.G.; Palacios-Rodríguez, G.; Navarro, F.B.; Blanco, J.A.; Imbert, J.B.; Cachinero-Vivar, A.M.; et al. Long-Term Carbon Sequestration in Pine Forests under Different Silvicultural and Climatic Regimes in Spain. Forests 2022, 13, 450. https://doi.org/10.3390/f13030450
Navarro-Cerrillo RM, Ruiz-Gómez FJ, Camarero JJ, Castillo V, Barberá GG, Palacios-Rodríguez G, Navarro FB, Blanco JA, Imbert JB, Cachinero-Vivar AM, et al. Long-Term Carbon Sequestration in Pine Forests under Different Silvicultural and Climatic Regimes in Spain. Forests. 2022; 13(3):450. https://doi.org/10.3390/f13030450
Chicago/Turabian StyleNavarro-Cerrillo, Rafael M., Francisco J. Ruiz-Gómez, Jesús J. Camarero, Víctor Castillo, Gonzalo G. Barberá, Guillermo Palacios-Rodríguez, Francisco B. Navarro, Juan A. Blanco, Juan B. Imbert, Antonio M. Cachinero-Vivar, and et al. 2022. "Long-Term Carbon Sequestration in Pine Forests under Different Silvicultural and Climatic Regimes in Spain" Forests 13, no. 3: 450. https://doi.org/10.3390/f13030450
APA StyleNavarro-Cerrillo, R. M., Ruiz-Gómez, F. J., Camarero, J. J., Castillo, V., Barberá, G. G., Palacios-Rodríguez, G., Navarro, F. B., Blanco, J. A., Imbert, J. B., Cachinero-Vivar, A. M., Molina, A. J., & del Campo, A. D. (2022). Long-Term Carbon Sequestration in Pine Forests under Different Silvicultural and Climatic Regimes in Spain. Forests, 13(3), 450. https://doi.org/10.3390/f13030450