
Inherent Water-Use Efficiency of Different Forest Ecosystems and Its Relations to Climatic Variables


Abstract
:1. Introduction
2. Materials and Methods
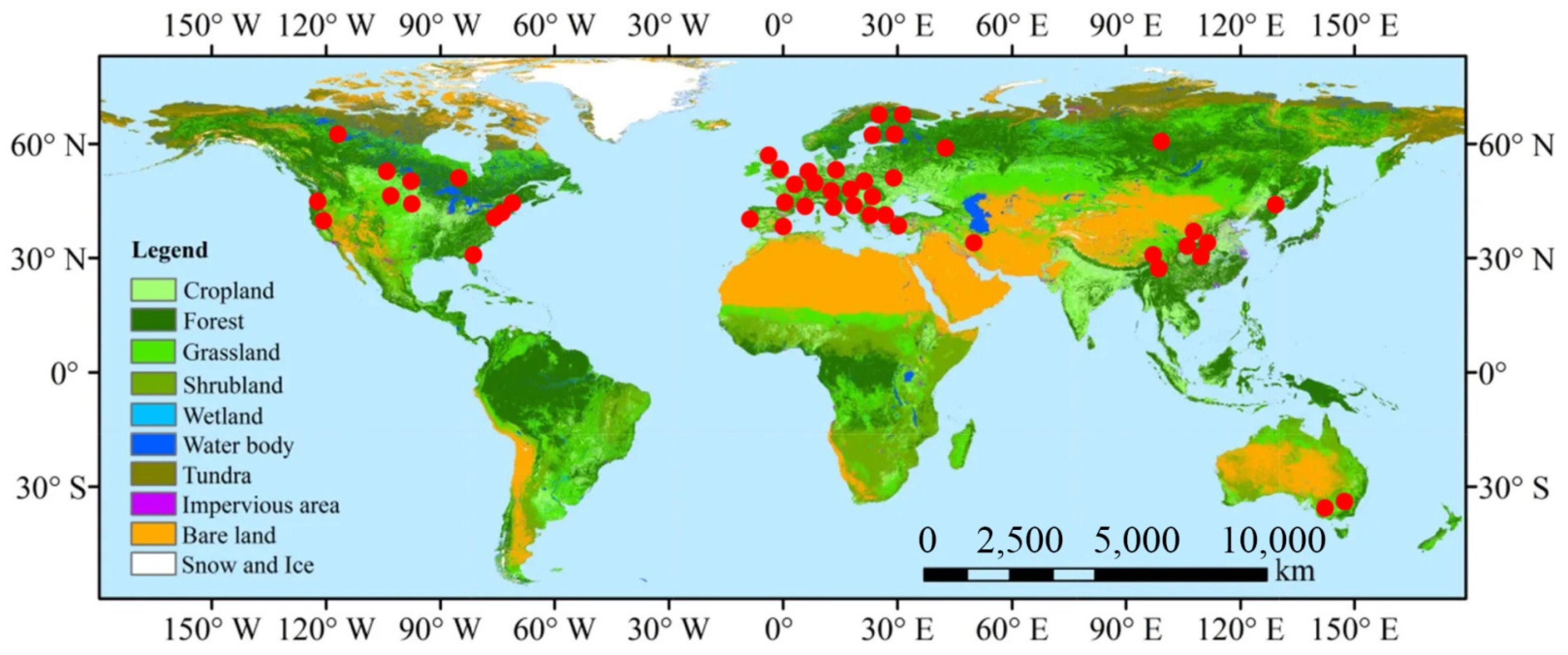
2.1. Data Sources and Processing
2.2. The IWUE of Forest Ecosystems
2.3. Statistical Analysis
3. Results
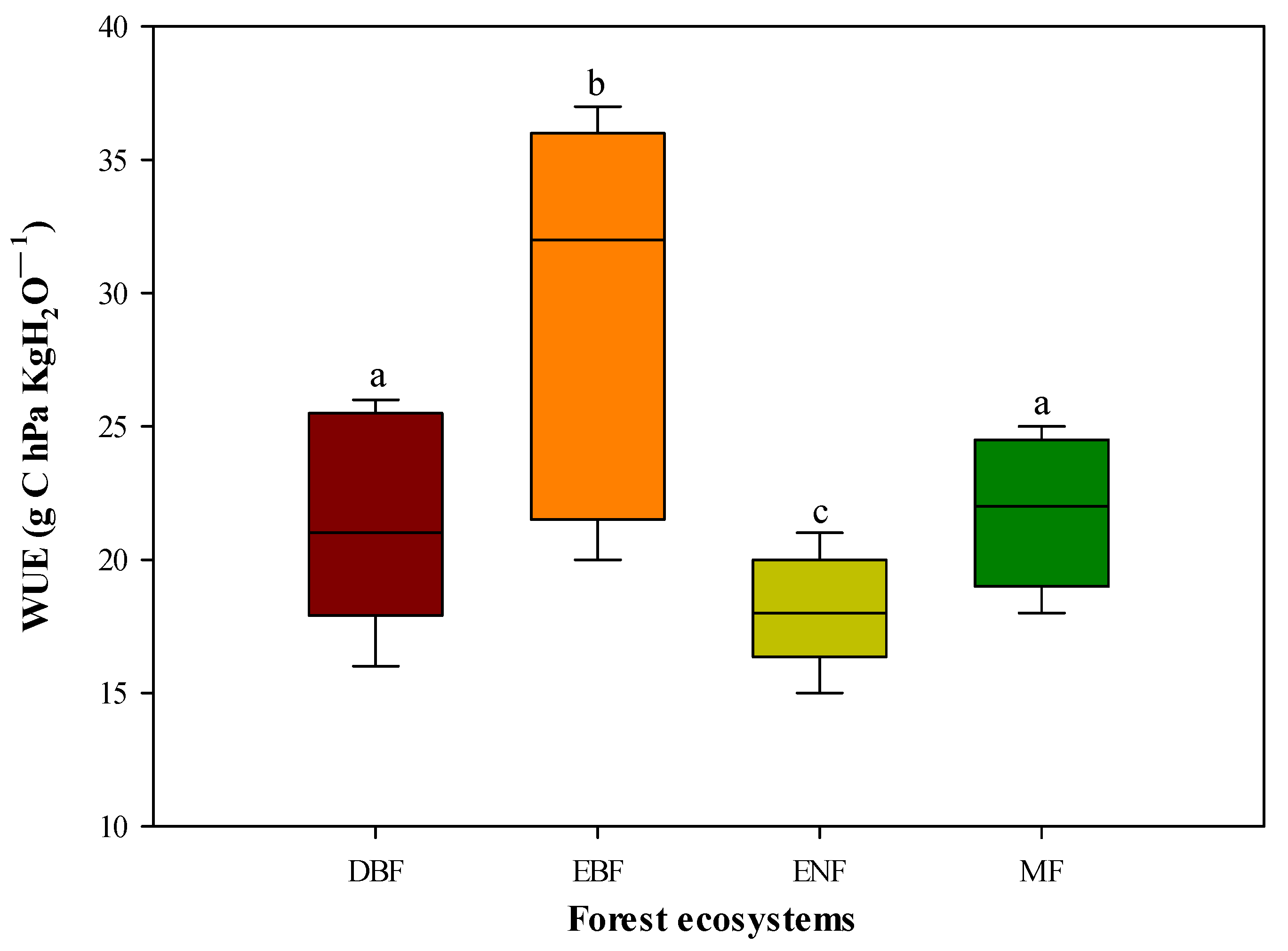
3.1. Differences in the IWUE of Forest Ecosystems
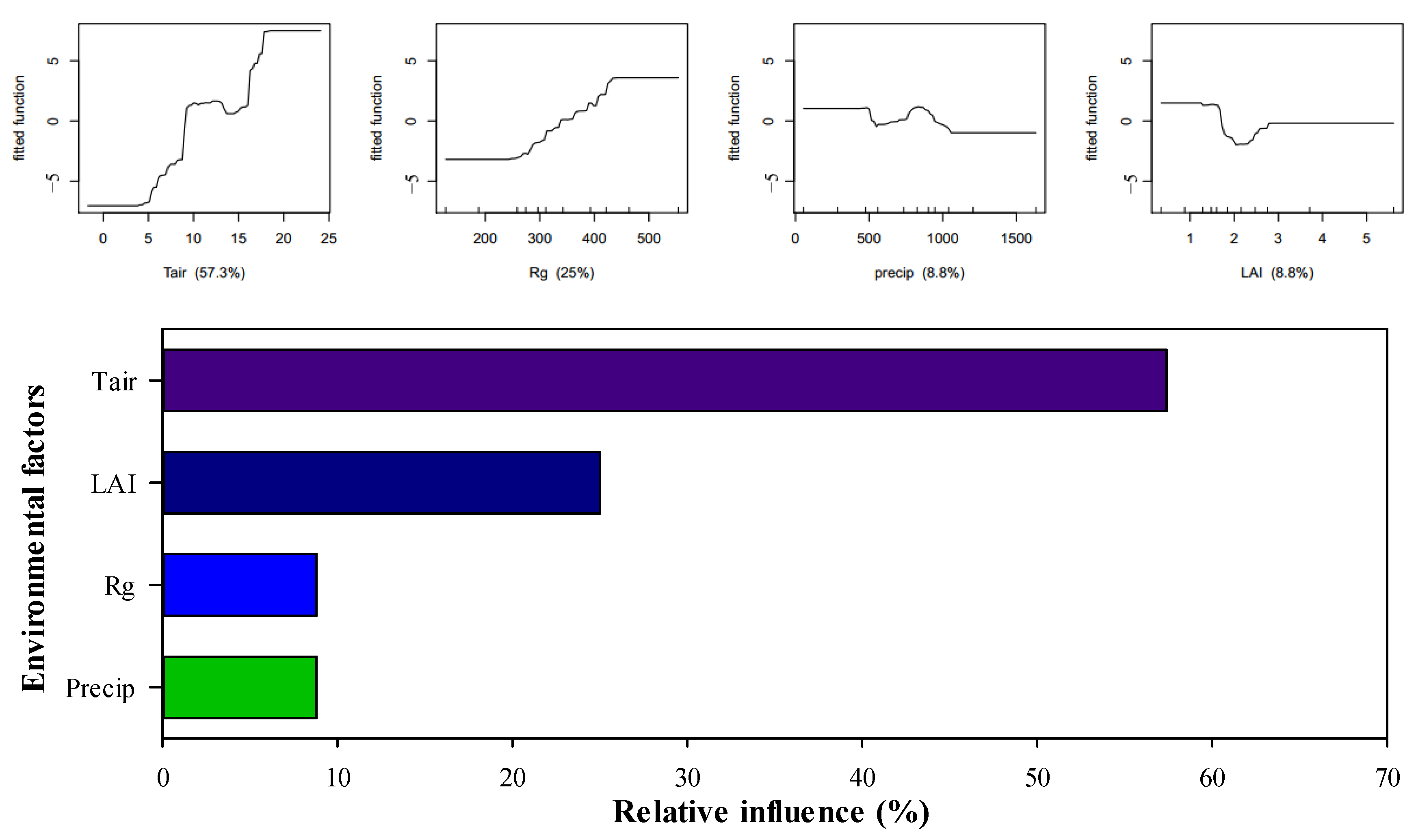
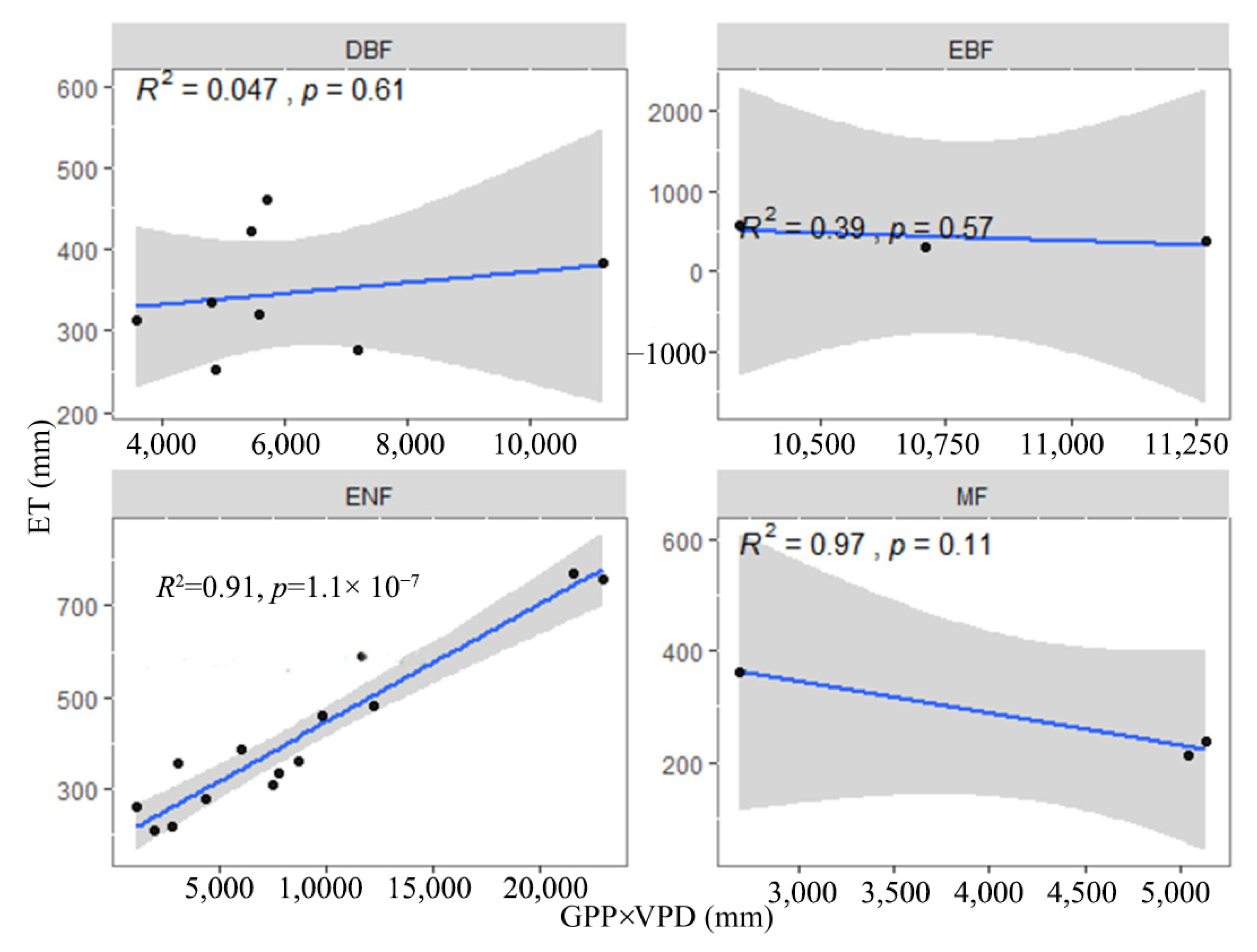
3.2. Effects of Environmental Factors on the IWUE of Different Ecosystems
3.3. Influence Weight of Environmental Factors on the IWUE
3.4. Factors Influencing IWUE Interannual Changes
4. Discussion
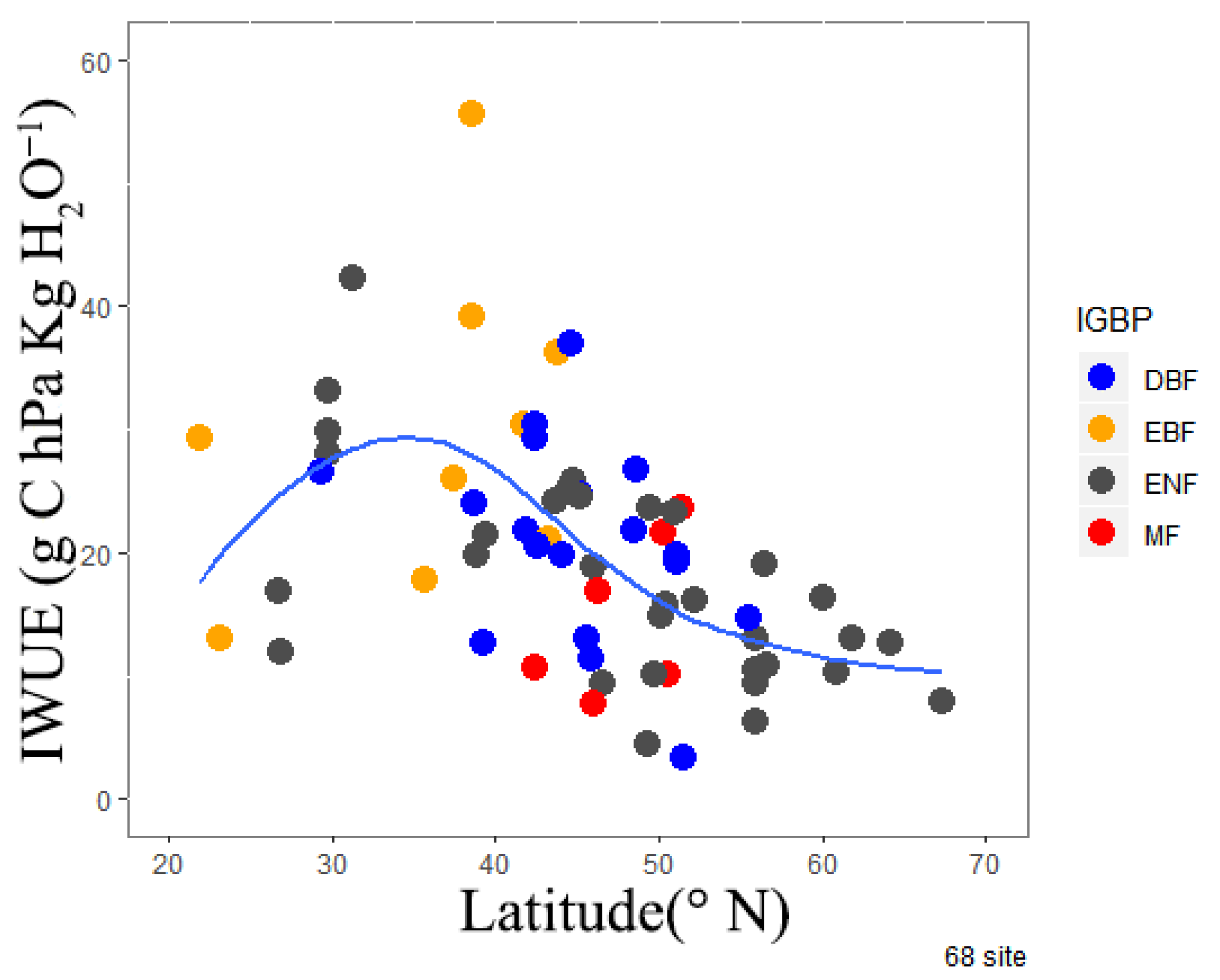
4.1. Spatial Distribution of the IWUE of Forest Ecosystems
4.2. Effects of Environmental Factors on the IWUE in Different Ecosystems
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vye, R. Cultivation of plants in space: Their contribution to stabilizing atmospheric composition in closed ecological systems. Adv. Space Res. 1996, 18, 165–176. [Google Scholar]
- Hu, Z.; Yu, G.; Fu, Y.; Sun, X.; Li, Y.; Shi, P.; Zheng, Z. Effects of vegetation control on ecosystem water use efficiency within and among four grassland ecosystems in China. Glob. Change Biol. 2008, 14, 1609–1619. [Google Scholar] [CrossRef]
- Belén, A.M.; Valladares, F.; Irene, M.F. Both mature patches and expanding areas of Juniperus thurifera forests are vulnerable to climate change but for different reasons. Forests 2020, 11, 960. [Google Scholar]
- Giampiero, T.; Riccardo, V. Severe drought effects on ecosystem CO2 and H2O fluxes at three Mediterranean evergreen sites: Revision of current hypotheses. Glob. Change Biol. 2010, 8, 999–1017. [Google Scholar]
- Beer, C.; Ciais, P.; Reichstein, M.; Baldocchi, D.; Law, B.E.; Papale, D.; Wohlfahrt, G. Temporal and among-site variability of inherent water use efficiency at the ecosystem level. Glob. Biogeochem. Cycles 2009, 23, 1–13. [Google Scholar] [CrossRef]
- Yu, G.R.; Song, X.; Wang, Q.F.; Liu, Y.F.; Guan, D.X.; Yan, J.H.; Sun, X.M.; Zhang, L.M.; Wen, X.F. Water-use efficiency of forest ecosystems in eastern China and its relations to climatic variables. New Phytol. 2010, 177, 927–937. [Google Scholar] [CrossRef]
- Liu, Z.Q.; Liu, Q.Q.; Wei, Z.J.; Yu, X.X.; Jia, G.D.; Jiang, J. Partitioning tree water usage into storage and transpiration in a mixed forest. For. Ecosyst. 2021, 8, 13. [Google Scholar] [CrossRef]
- Yang, S.; Zhang, J.; Han, J. Evaluating global ecosystem water use efficiency response to drought based on multi-model analysis. Sci. Total Environ. 2021, 778, 146356. [Google Scholar] [CrossRef]
- Mk, A.; Nr, A.; Tl, A. Carbon sequestration and water yield tradeoffs following restoration of abandoned agricultural lands in Mediterranean mountains. Environ. Res. 2021, 207, 112203. [Google Scholar]
- Tong, X.; Wu, P.; Liu, X. A global meta-analysis of fruit tree yield and water use efficiency under deficit irrigation. Agric. Water Manag. 2022, 260, 107321. [Google Scholar] [CrossRef]
- Hutley, L.B.; Beringer, J.; Fatichi, S. Gross primary productivity and water use efficiency are increasing in a high rainfall tropical savanna. Glob. Change Biol. 2022, 28, 2360–2380. [Google Scholar] [CrossRef]
- Baldocchi, D.; Chu, H.; Reichstein, M. Inter-annual variability of net and gross ecosystem carbon fluxes: A review. Agric. For. Meteorol. 2018, 249, 520–533. [Google Scholar] [CrossRef] [Green Version]
- Ponton, S.; Flanagan, L.B.; Alstad, K.P.; Johnson, B.G.; Morgenstern, K.A.I.; Kljun, N.; Barr, A.G. Comparison of ecosystem water-use efficiency among Douglas-fir forest, aspen forest and grassland using eddy covariance and carbon isotope techniques. Glob. Change Biol. 2006, 12, 294–310. [Google Scholar] [CrossRef]
- Saurer, M.; Spahni, R.; Frank, D.C.; Joos, F.; Leuenberger, M.; Loader, N.J.; Young, G.H. Spatial variability and temporal trends in water-use efficiency of European forests. Glob. Change Biol. 2014, 20, 3700–3712. [Google Scholar] [CrossRef]
- Huxman, T.E.; Smith, M.D.; Fay, P.A.; Knapp, A.K.; Shaw, M.R.; Loik, M.E.; Smith, S.D. Convergence across biomes to a common rain-use efficienc. Nature 2004, 429, 649–651. [Google Scholar] [CrossRef]
- Huang, M.; Piao, S.; Sun, Y.; Ciais, P.; Cheng, L.; Mao, J.; Wang, Y. Change in terrestrial ecosystem water-use efficiency over the last three decades. Glob. Change Biol. 2015, 21, 2366–2378. [Google Scholar] [CrossRef]
- Lu, K.; Chen, N.; Zhang, X.; Wang, J.; Wang, M.; Khan, S.; Zhao, C. Increased drought and atmospheric CO2 positively impact intrinsic water use efficiency but do not promote tree growth in semi-arid areas of northwestern China. Trees 2019, 33, 669–679. [Google Scholar] [CrossRef]
- Topbjerg, H.B.; Kaminski, K.P.; Markussen, B.; Kørup, K.; Nielsen, K.L.; Kirk, H.G.; Liu, F. Physiological factors affecting intrinsic water use efficiency of potato clones within a dihaploid mapping population under well-watered and drought-stressed conditions. Sci. Hortic. 2014, 178, 61–69. [Google Scholar] [CrossRef]
- Frank, D.C.; Poulter, B.; Saurer, M.; Esper, J.; Huntingford, C.; Helle, G.; Weigl, M. Water-use efficiency and transpiration across European forests during the Anthropocene. Nat. Clim. Change 2015, 5, 579–583. [Google Scholar] [CrossRef] [Green Version]
- Kwon, H.; Law, B.E.; Thomas, C.K.; Johnson, B.G. The influence of hydrological variability on inherent water use efficiency in forests of contrasting composition, age, and precipitation regimes in the Pacific Northwest. Agric. For. Meteorol. 2017, 249, 488–500. [Google Scholar] [CrossRef]
- Jiang, S.Z.; Zhao, L.; Liang, C.; Hu, X.T.; Wang, Y.S.; Gong, D.Z.; Zheng, S.S.; Huang, Y.W.; He, Q.Y.; Cui, N.B. Leaf-and ecosystem-scale water use efficiency and their controlling factors of a kiwifruit orchard in the humid region of Southwest China. Agric. Water Manag. 2022, 260, 107329. [Google Scholar] [CrossRef]
- Zhao, T.T.; Zhang, Y.J.; Zhang, T.; Xu, M.J.; Zhu, J.T.; He, Y.T.; Yu, G.R. Drought occurrence and time dominated variations in water use efficiency in an alpine meadow on the Tibetan Plateau. Ecohydrology 2022, 5, 2360. [Google Scholar] [CrossRef]
- Zhang, Q.F.; Lv, X.Z.; Yu, X.X.; Ni, Y.X.; Ma, L.; Liu, Z.Q. Species and spatial differences of vegetation rainfall interception capacity a synthesis and meta-analysis in China. CATENA 2022, 213, 106223. [Google Scholar] [CrossRef]
- Liu, Z.Q.; Zhuang, H.; Jia, G.D.; Yu, X.X.; Jiang, J. Evidence of foliar water uptake in a conifer species. Agric. Water Manag. 2021, 255, 106993. [Google Scholar] [CrossRef]
- Medlyn, B.E.; Duursma, R.A.; Eamus, D.; Ellsworth, D.S.; Prentice, I.C.; Barton, C.V.M.; Crous, K.Y.; De Angelis, P.; Freeman, M.; Wingate, L. Reconciling the opticaland empirical approaches to modelling stomatal conductance. Glob. Change Biol. 2017, 2134–2144. [Google Scholar]
- Xiao, J.; Sun, G.; Chen, J.; Chen, H.; Chen, S.; Dong, G.; Zhou, J. Carbon fluxes, evapotranspiration, and water use efficiency of terrestrial ecosystems in China. Agric. For. Meteorol. 2013, 182–183, 76–90. [Google Scholar] [CrossRef]
- Liu, Z.Q.; Jia, G.D.; Yu, X.X. Variation of water uptake in degradation agroforestry shelterbelts on the North China Plain. Ag-Riculture Ecosyst. Environ. 2020, 287, 106697. [Google Scholar] [CrossRef]
- Gao, Y.; Markkanen, T.; Aurela, M. Response of water use efficiency to summer drought in a boreal Scots pine forest in Finland. Biogeosciences 2017, 14, 4409–4422. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, M.E.; Zwieniecki, M.A.; Holbrook, N.M. Independent variation in photosynthetic capacity and stomatal conductance leads to differences in intrinsic water use efficiency in 11 soybean genotypes before and during mild drought. J. Exp. Bot. 2011, 62, 2875–2887. [Google Scholar] [CrossRef]
- Ji, Y.D.; Li, Y.; Yao, N.; Biswas, A.; Zou, Y.F.; Meng, Q.T.; Liu, F.G. The lagged effect and impact of soil moisture drought on terrestrial ecosystem water use efficiency. Ecol. Indic. 2021, 133, 108349. [Google Scholar] [CrossRef]
- Haynes, A.F. Presence of n fixing neighbors increases 1eaf n and d13c in casti11eja app1egatei a root hemiparasite. Plant Ecol. 2021, 223, 213–228. [Google Scholar] [CrossRef]
- Jassal, R.S.; Black, T.A.; Spittlehouse, D.L.; Brümmer, C.; Nesic, Z. Evapotranspiration and water use efficiency in different-aged Pacific Northwest Douglas-fir stands. Agric. For. Meteorol. 2009, 149, 1168–1178. [Google Scholar] [CrossRef]
- Krishnan, P.; Black, T.A.; Grant, N.J. Impact of changing soil moisture distribution on net ecosystem productivity of a boreal aspen forest during and following drought. Agric. For. Meteorol. 2006, 139, 208–223. [Google Scholar] [CrossRef]
- Nie, C.; Huang, Y.F.; Zhang, S.; Yang, Y.T.; Zhou, S.; Lin, C.J.; Wang, G.Q. Effects of soil water content on forest ecosystem water use efficiency through changes in transpiration/evapotranspiration ratio. Agric. For. Meteorol. 2021, 308, 108605. [Google Scholar] [CrossRef]
- Heilman, K.A.; Trouet, V.M.; Belmecheri, S. Increased water use efficiency leads to decreased precipitation sensitivity of tree growth, but is offset by high temperatures. Oecologia 2021, 197, 1095–1110. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Symbol |
|---|---|
| water use efficiency | WUE |
| intrinsic water use efficiency | IWUE |
| evapotranspiration | ET |
| gross primary productivity | GPP |
| vapor pressure deficit | VPD |
| temperature | T |
| precipitation | P |
| leaf area index | LAI |
| mean diurnal course | MDC |
| look-up table | LUT |
| deciduous broadleaf forests | DBF |
| evergreen broadleaf forests | EBF |
| evergreen needle-leaf forests | ENF |
| mixed forests | MF |
| inherent water-use efficiency | ΔWUE |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.; Ji, X.; Ye, L.; Jiang, J. Inherent Water-Use Efficiency of Different Forest Ecosystems and Its Relations to Climatic Variables. Forests 2022, 13, 775. https://doi.org/10.3390/f13050775
Liu Z, Ji X, Ye L, Jiang J. Inherent Water-Use Efficiency of Different Forest Ecosystems and Its Relations to Climatic Variables. Forests. 2022; 13(5):775. https://doi.org/10.3390/f13050775
Chicago/Turabian StyleLiu, Ziqiang, Xiaofang Ji, Limin Ye, and Jiang Jiang. 2022. "Inherent Water-Use Efficiency of Different Forest Ecosystems and Its Relations to Climatic Variables" Forests 13, no. 5: 775. https://doi.org/10.3390/f13050775
APA StyleLiu, Z., Ji, X., Ye, L., & Jiang, J. (2022). Inherent Water-Use Efficiency of Different Forest Ecosystems and Its Relations to Climatic Variables. Forests, 13(5), 775. https://doi.org/10.3390/f13050775