
Assimilation Efficiencies and Gas Exchange Responses of Four Salix Species in Elevated CO2 under Soil Moisture Stress and Fertilization Treatments

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material, Growing Conditions, and Treatment Delivery
2.2. Assimilation Efficiencies and Gas Exchange Experiment: 2017
2.3. CO2 × Fertilizer Experiment: 2018
2.4. Statistical Analysis
3. Results
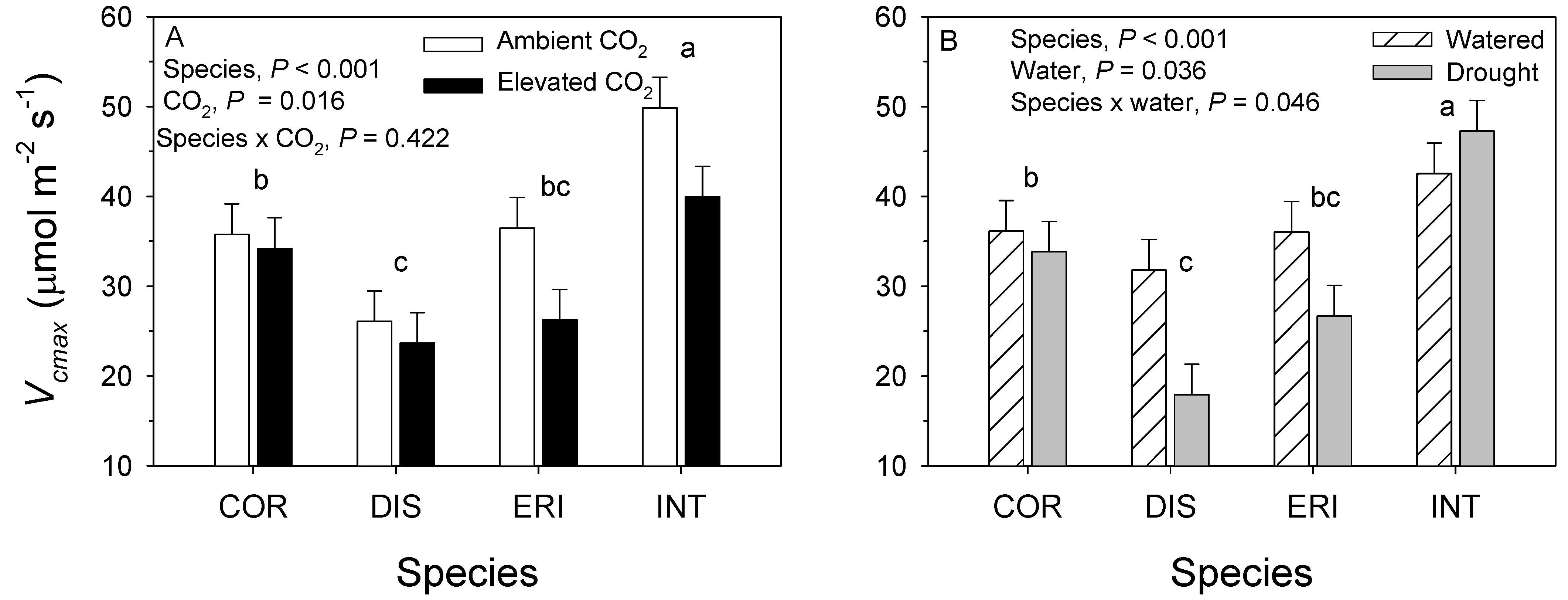
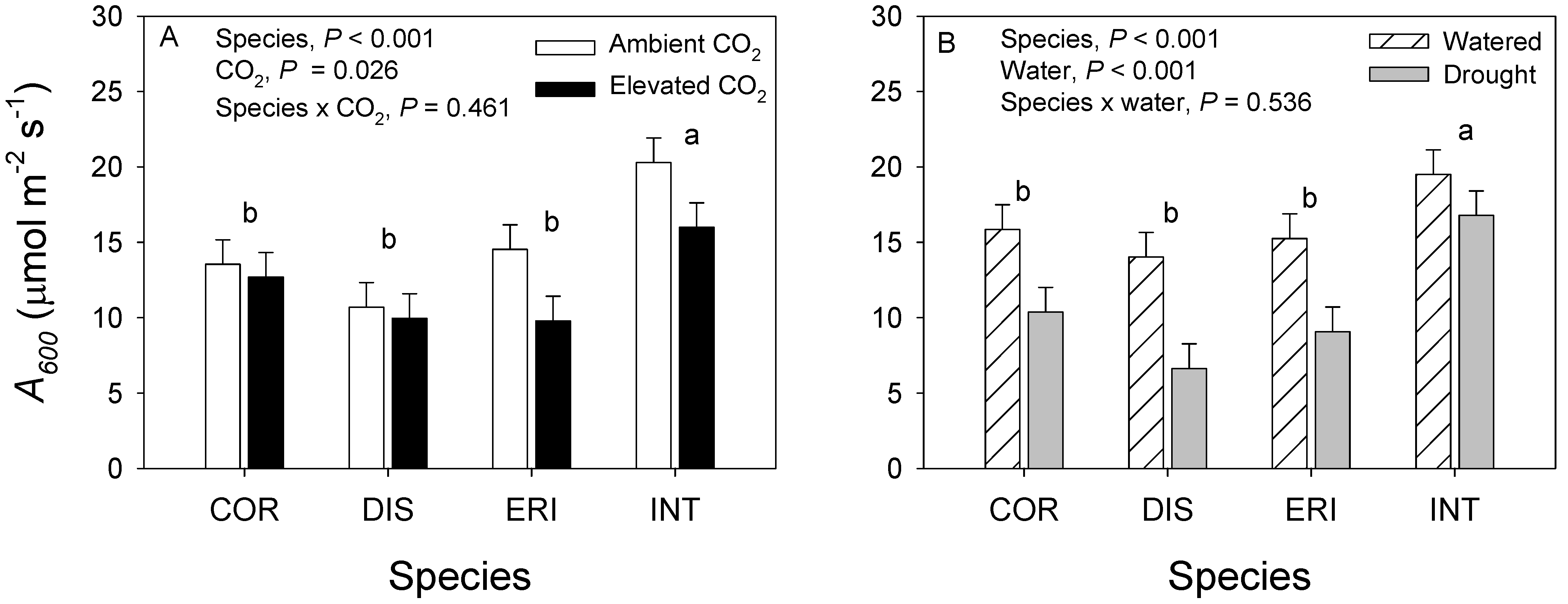
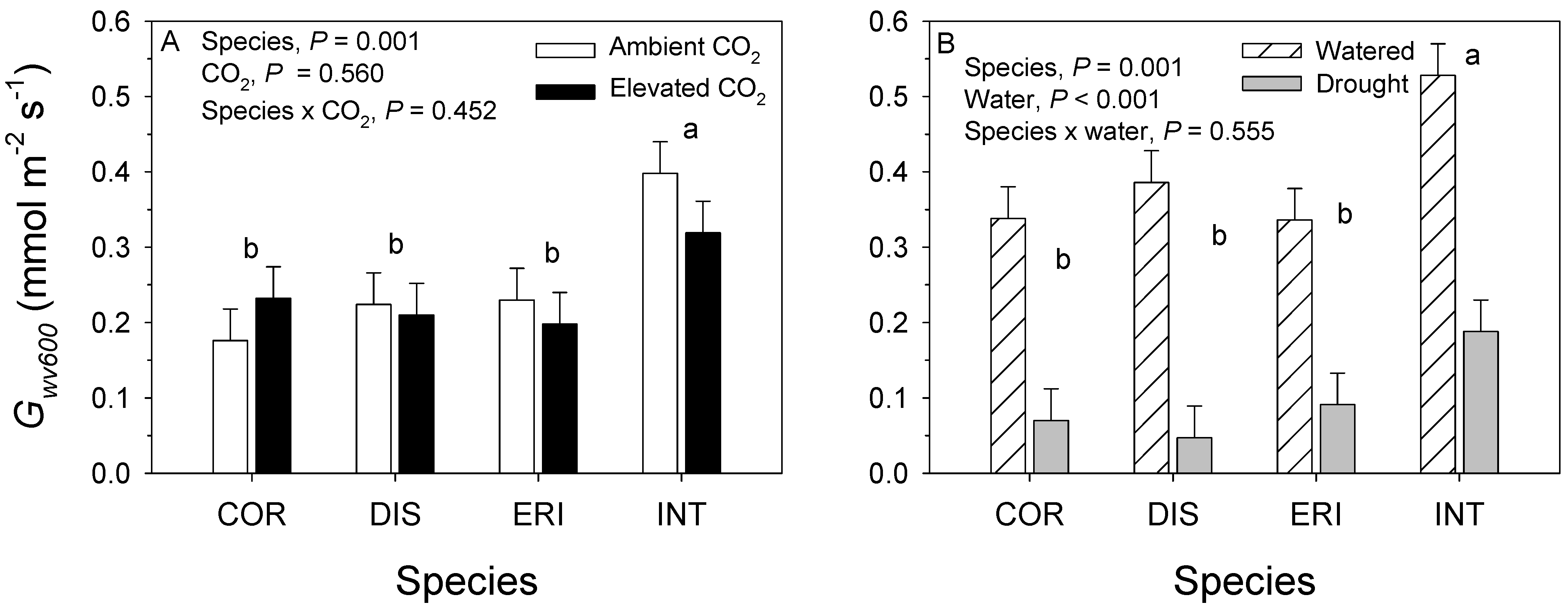
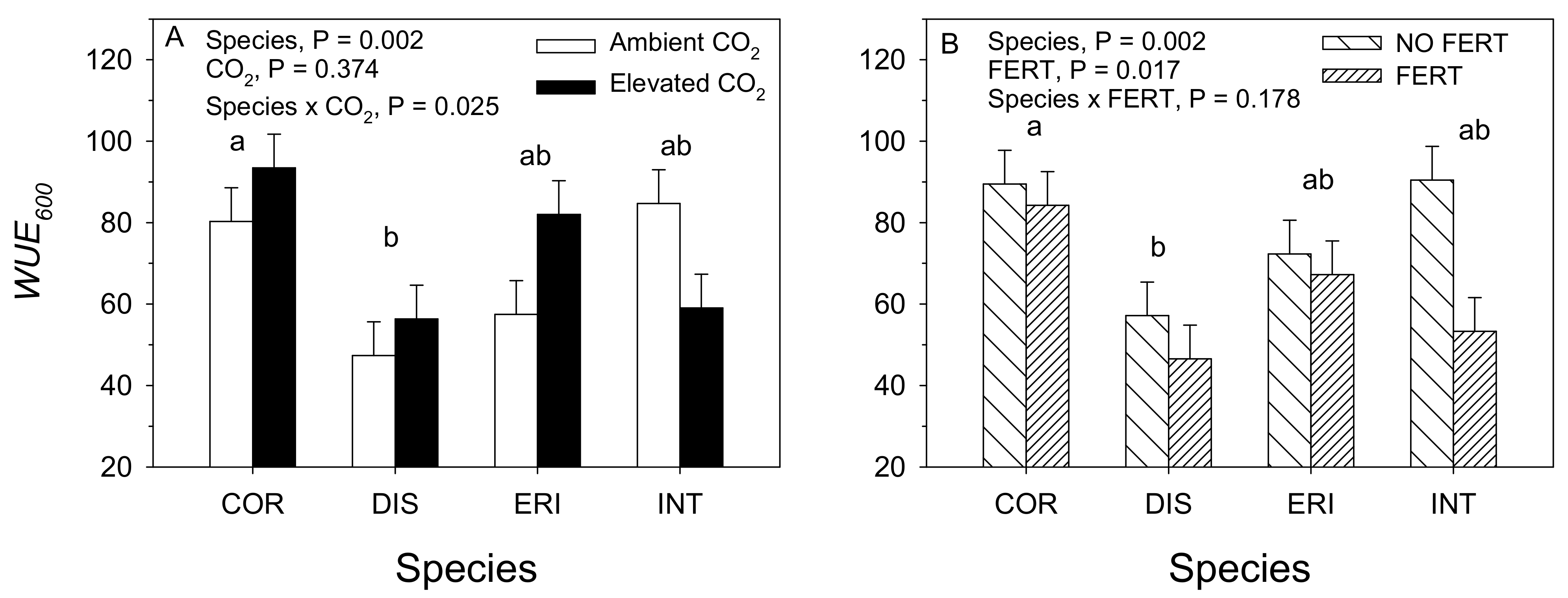
3.1. 2017. CO2 × Water Experiment
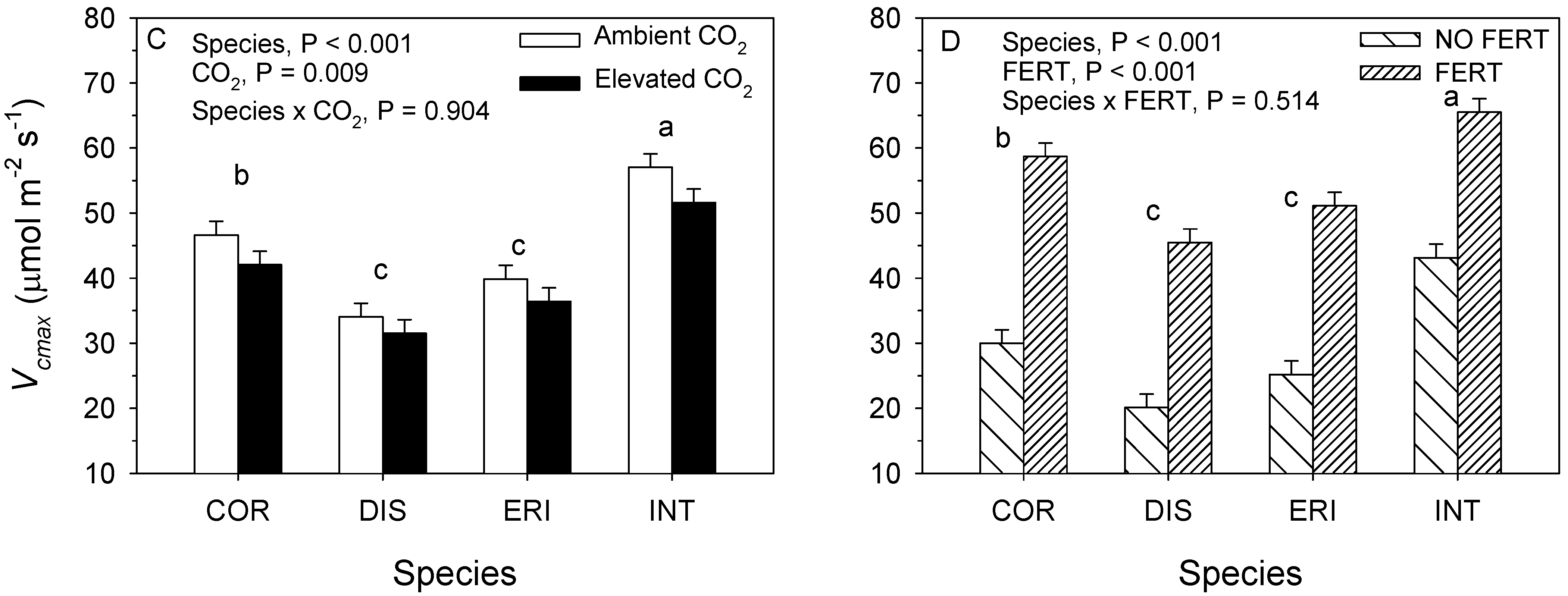
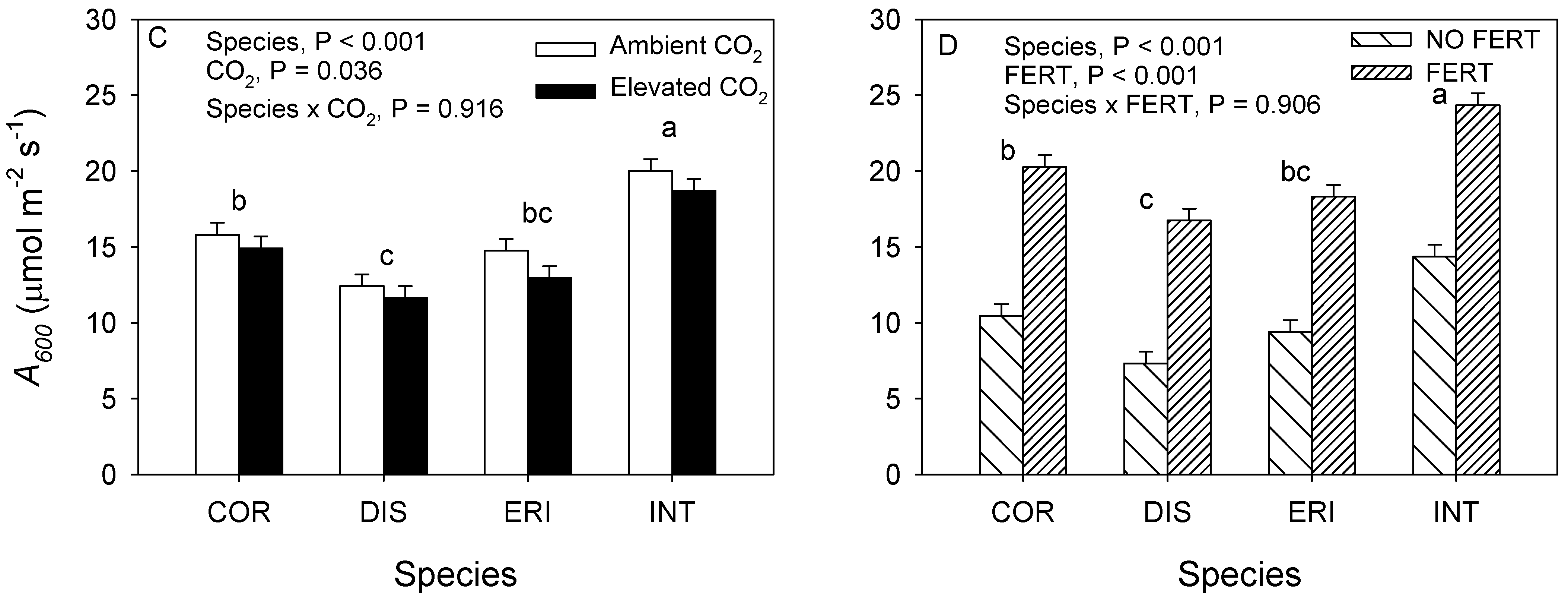
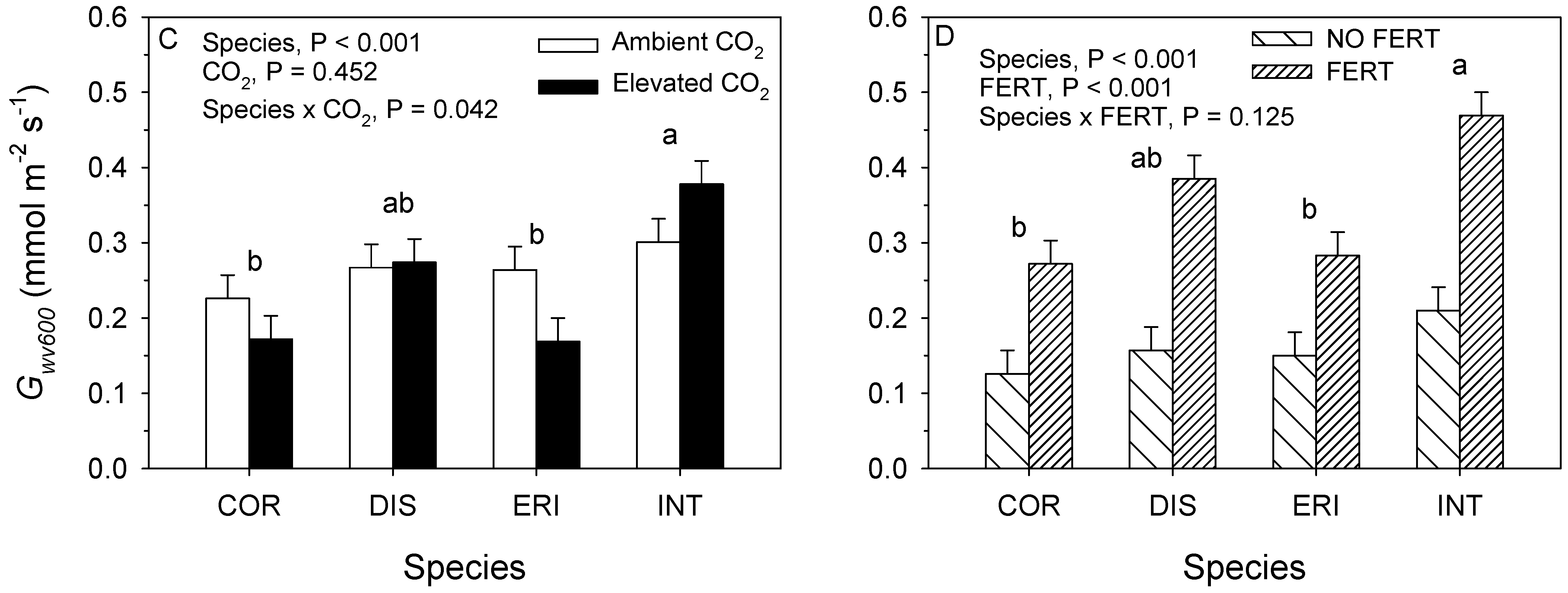
3.2. 2018. CO2 × Fertilizer Experiment
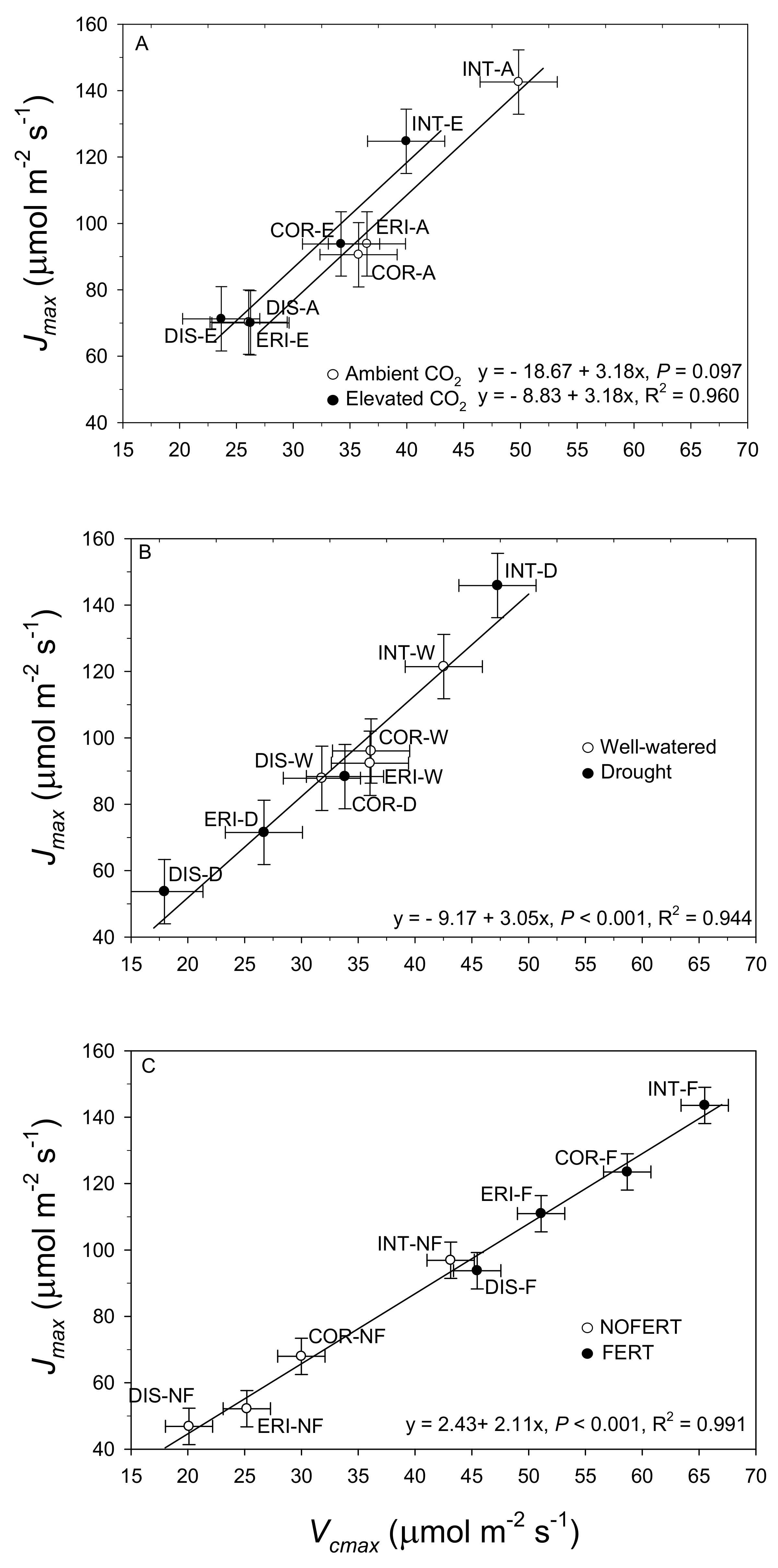
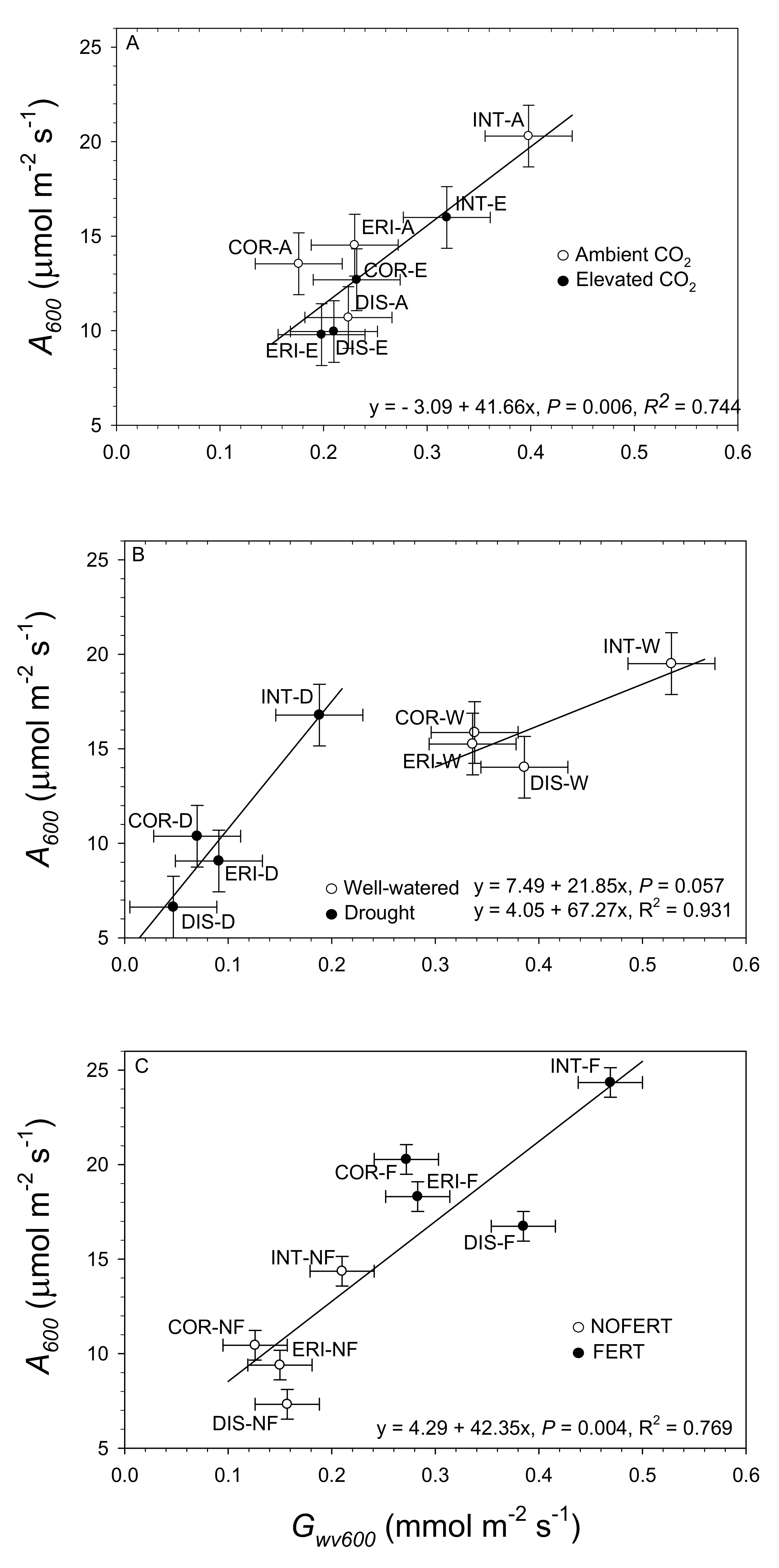
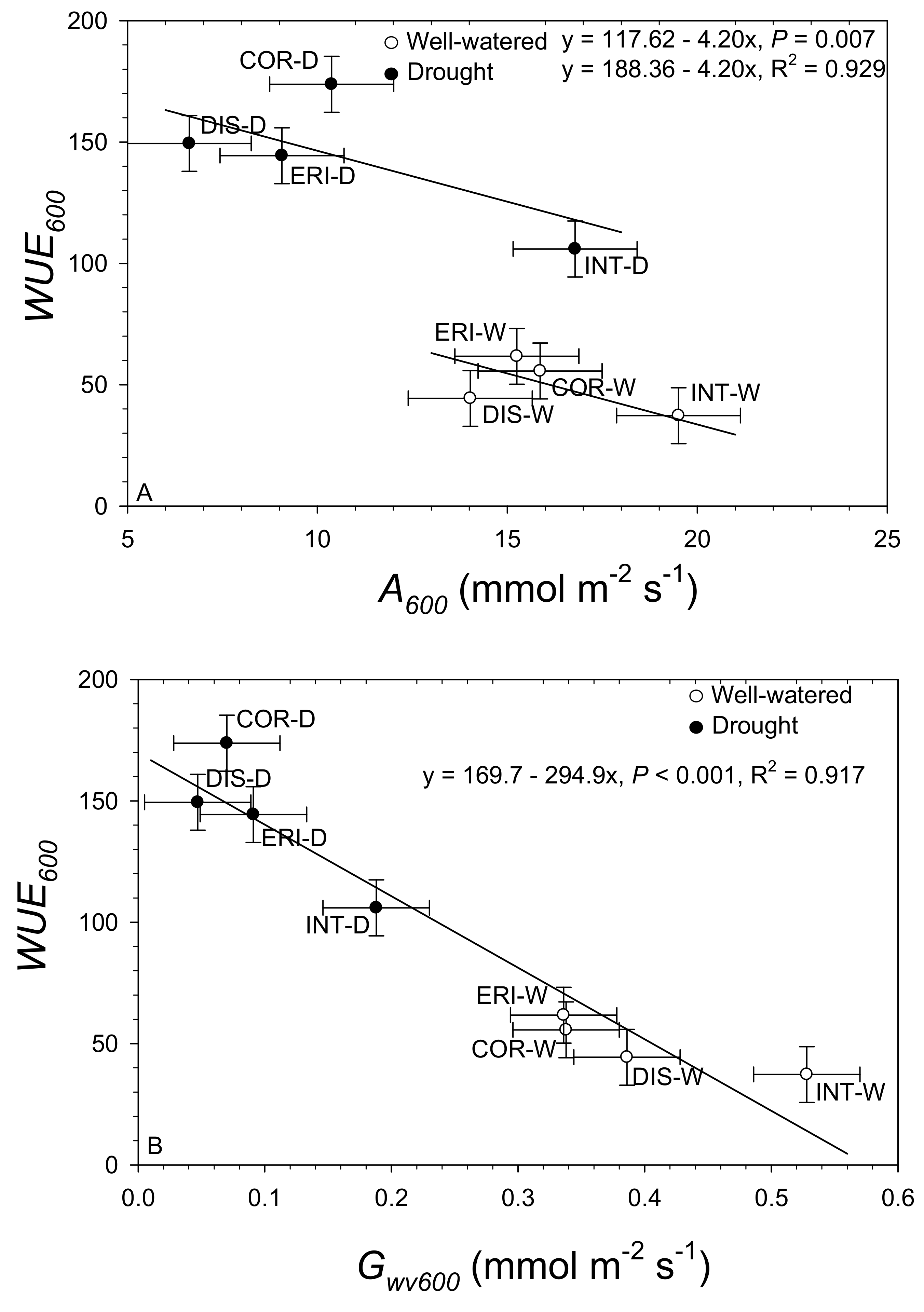
3.3. Covariate Relationships among Willow Species and Environmental Change
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bauer, G.A.; Berntson, G.M.; Bazzaz, F.A. Regenerating temperate forests under elevated CO2 and nitrogen deposition: Comparing biochemical and stomatal limitations of photosynthesis. New Phytol. 2001, 152, 249–266. [Google Scholar] [CrossRef]
- Zhou, Y.M.; Wang, C.G.; Han, S.J.; Cheng, X.B.; Li, M.H.; Fan, A.N.; Wang, X.X. Species-specific and needle age-related responses of photosynthesis in two Pinus species to long-term exposure to elevated CO2 concentration. Trees 2011, 25, 163–173. [Google Scholar] [CrossRef] [Green Version]
- Greenup, H.; Turnbull, M.H.; Whitehead, D. Response of photosynthesis in second-generation Pinus radiata trees to long-term exposure to elevated carbon dioxide partial pressure. Tree Physiol. 2003, 23, 569–576. [Google Scholar] [CrossRef] [PubMed]
- Ellsworth, D.S.; Reich, P.B.; Nauburg, E.S.; Koch, G.W.; Kubiske, M.E.; Smith, S.D. Photosynthesis, carboxylation and leaf nitrogen responses of 16 species to elevated pCO2 across four free-air CO2 enrichment experiments in forest, grassland and desert. Glob. Chg. Biol. 2004, 10, 2121–2138. [Google Scholar] [CrossRef] [Green Version]
- Crous, K.Y.; Walters, M.B.; Ellsworth, D.S. Elevated CO2 concentration affects leaf photosynthesis—nitrogen relationships in Pinus teada over nine years in FACE. Tree Physiol. 2008, 28, 607–614. [Google Scholar] [CrossRef]
- Major, J.E.; Mosseler, A.; Malcolm, J.W. Genetic variation among pines and spruces in assimilation efficiencies and photosynthetic regulation under elevated CO2. Trees Struct. Funct. 2018, 32, 215–229. [Google Scholar] [CrossRef]
- Luomala, E.M.; Laitinen, K.; Kellomaki, S.; Vapavuori, E. Variable photosynthetic acclimation in consecutive cohorts of Scots pine needles during 3 years of growth at elevated CO2 and elevated temperature. Plant Cell Environ. 2003, 26, 645–660. [Google Scholar] [CrossRef]
- Trenberth, K.E.; Dai, A.; Van der Schreir, G.; Jones, P.D.; Barichivich, J.; Briffa, K.R.; Sheffield, J. Global warming and changes in drought. Nature Clim. Chang. 2014, 4, 17–22. [Google Scholar] [CrossRef]
- Allen, C.D.; Breshears, D.D.; McDowell, N.G. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 2015, 6, 1–55. [Google Scholar] [CrossRef]
- Attia, Z.; Domec, J.C.; Oren, R.; Way, D.A.; Moshelion, M. Growth and physiological responses of isohydric and anisohydric poplars to drought. J. Expt. Bot. 2015, 66, 4373–4381. [Google Scholar] [CrossRef] [Green Version]
- Teskey, R.O.; Wertin, T.; Bauweraerts, I.; Ameye, M.; McGuire, M.A.; Steppe, K. Responses of tree species to heat waves and extreme heat events. Plant Cell Environ. 2015, 38, 1699–1712. [Google Scholar] [CrossRef] [PubMed]
- Bhusal, N.; Lee, M.; Lee, H.; Adhikari, A.; Hane, A.R.; Han, A.; Kim, H.S. Evaluation of morphological, physiological, and biochemical traits for assessing drought resistance in eleven tree species. Sci. Total Environ. 2021, 779, 146466. [Google Scholar] [CrossRef] [PubMed]
- Duan, H.; O’Grady, P.O.; Duursma, R.A.; Choat, B.; Huang, G.; Smith, R.A.; Jiang, Y.; Tissue, D.T. Drought response of two gymnosperm species with contrasting stomatal regulation strategies under elevated CO2 and temperature. Tree Physiol. 2015, 35, 756–770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosseler, A.; Zsuffa, L.; Stoehr, M.U.; Kenney, W.A. Variation in biomass production, moisture content, and specific gravity in some North American willows (Salix L.). Can. J. For. Res. 1988, 18, 1535–1540. [Google Scholar] [CrossRef]
- Major, J.E.; Mosseler, A.; Malcolm, J.W. Salix species variation in gas exchange, leaf sodium and nutrient parameters at three levels of salinity. Can. J. For. Res. 2017, 47, 1045–1055. [Google Scholar] [CrossRef]
- Major, J.E.; Mosseler, A.; Malcolm, J.W.; Heartz, S. Salinity tolerance of three Salix species: Survival, biomass yield and allocation, and biochemical efficiencies. Biomass Bioenergy 2017, 105, 10–22. [Google Scholar] [CrossRef]
- Johnson, J.D.; Tognetti, R.; Paris, P. Water relations and gas exchange in poplar and willow under water stress and elevated CO2. Physiol. Plant 2002, 115, 93–100. [Google Scholar] [CrossRef]
- Fabio, E.S.; Leary, C.J.; Smart, L.B. Tolerance of novel inter-specific shrub willow hybrids to water stress. Trees 2019, 33, 1015–1026. [Google Scholar] [CrossRef]
- Li, S.; Pezeshki, S.R.; Goodwin, S.; Shields, F.D., Jr. Physiological responses of black willow (Salix nigra) cuttings to a range of soil moisture regimes. Photosynthetica 2004, 42, 585–590. [Google Scholar] [CrossRef]
- Savage, J.A.; Cavender-Bares, J.; Verhoven, A. Willow species (genus: Salix) with contrasting habitat affinities differ in their photoprotective responses to water stress. Funct. Plant Biol. 2009, 36, 300–309. [Google Scholar] [CrossRef] [Green Version]
- Parent, S.; Messier, C. A simple and efficient method to estimate microsite light availability under a forest canopy. Can. J. For. Res. 1996, 26, 151–154. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Von, C.S.; Berry, J.A. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef] [Green Version]
- Harley, P.C.; Thomas, R.B.; Reynolds, J.F.; Strain, B.R. Modelling photosynthesis of cotton grown in elevated CO2. Plant Cell Environ. 1992, 15, 271–282. [Google Scholar] [CrossRef]
- Mosseler, A.; Major, J.E. Phytoremediation efficacy of Salix discolor and S. eriocephela on adjacent acidic clay and shale overburden on a former mine site: Growth, soil and foliage traits. Forests 2017, 8, 475. [Google Scholar] [CrossRef] [Green Version]
- Hicks, C.R. Fundamental Concepts in the Design of Experiments, 3rd ed.; Holt, Reinhart, Winston—Saunders College Publishing: New York, NY, USA, 1982. [Google Scholar]
- Major, J.E.; Mosseler, A.; Malcolm, J.W. Growth and biomass allocation responses of four Salix species in elevated CO2, under soil moisture stress and fertilization treatments. 2022; Unpublished (in preparation to be submitted). [Google Scholar]
- Kucukahmetler, O. The effects of salinity on yield and quality of ornamental plants and cut flowers. Acta Hort 2002, 573, 407–414. [Google Scholar] [CrossRef]
- Dorn, R.D. A synopsis of American Salix. Can. J. Bot. 1976, 54, 2769–2789. [Google Scholar] [CrossRef]
- Brunsfeld, S.J.; Soltis, D.E.; Soltis, P.S. Patterns of genetic variation in Salix section Longifoliae (Salicaceae). Am. J. Bot. 1991, 78, 855–869. [Google Scholar] [CrossRef]
- Brunsfeld, S.J.; Soltis, D.E.; Soltis, P.S. Evolutionary patterns and processes in Salixsect. Longifoliae: Evidence from chloroplast DNA. Syst. Bot. 1992, 17, 239–256. [Google Scholar]
- Briggs, M. Riparian Ecosystem Recovery in Arid Lands: Strategies and References; University of Arizona Press: Tucson, AZ, USA, 1996. [Google Scholar]
- Bhusal, N.; Han, S.-G.; Yoon, T.-M. Impact of drought stress on photosynthetic response, leaf water potential, and stem sap flow in two cultivars of bi-leader apple trees (Malus × domestica Borkh.). Sci. Hort. 2019, 246, 535–543. [Google Scholar] [CrossRef]
- Major, J.E.; Johnsen, K.H. Shoot water relations of mature black spruce families displaying a genotype × environment interaction in growth rate. II. Temporal trends and response to varying soil water conditions. Tree Physiol. 1999, 19, 375–382. [Google Scholar] [CrossRef] [Green Version]
- Limousin, J.M.; Roussel, A.; Rodríguez-Calcerrada, J.; Torres-Ruiz, J.M.; Moreno, M.; de Jalon, L.G.; Ourcival, J.-M.; Simioni, G.; Cochard, H.H.; Martin-St Paul, N. Drought acclimation of Quercus ilex leaves improves tolerance to moderate drought but not resistance to severe water stress. Plant Cell Environ. 2022. [Google Scholar] [CrossRef] [PubMed]
- Crous, K.Y.; Ellsworth, D.S. Canopy position affects photosynthetic adjustments to long-term elevated CO2 concentration (FACE) in aging needles in a mature Pinus taeda forest. Tree Physiol. 2004, 24, 961–970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ainsworth, E.A.; Long, S.P. What have we learned from 15 years of free-air CO2 enrichment (FACE)? A meta-analytic review of the responses of photosynthesis, canopy properties and plant production to rising CO2. New Phytol. 2005, 165, 351–372. [Google Scholar] [CrossRef] [PubMed]
- Tissue, D.T.; Griffin, K.L.; Ball, T. Photosynthetic adjustment in field-grown ponderosa pine trees after six years of exposure to elevated CO2. Tree Physiol. 1999, 19, 221–228. [Google Scholar] [CrossRef]
- Urban, O.; Marek, M.V. Seasonal changes of selected parameters of CO2 fixation biochemistry of Norway spruce under the long-term impact of elevated CO2. Photosynthetica 1999, 36, 533–545. [Google Scholar] [CrossRef]
- Tjoelker, M.G.; Oleksyn, J.; Reich, P.B. Seedlings of five boreal tree species differ in acclimation of net photosynthesis to elevated CO2 and temperature. Tree Physiol. 1998, 18, 715–726. [Google Scholar] [CrossRef] [Green Version]
- Centritto, M.; Jarvis, P.G. Long-term effects of elevated carbon dioxide concentration and provenance on four clones of Sitka spruce (Picea sitchensis). II. Photosynthetic capacity and nitrogen use efficiency. Tree Physiol. 1999, 19, 807–814. [Google Scholar] [CrossRef] [Green Version]
- Griffin, K.L.; Tissue, D.T.; Turnbull, M.H.; Whitehead, D. The onset of photosynthetic acclimation to elevated CO2 partial pressure in field-grown Pinus radiata D. Don. after 4 years. Plant Cell Environ. 2000, 23, 1089–1098. [Google Scholar] [CrossRef]
- Oren, R.; Ellsworth, D.S.; Johnsen, K.H.; Phillips, N.; Ewers, B.E.; Maier, C.; Katul, G.G.; Schäfer, K.V.R.; McCarthy, H.; Hendrey, G.; et al. Soil fertility limits carbon sequestration by forest ecosystems in a CO2-enriched atmosphere. Nature 2001, 411, 469–472. [Google Scholar] [CrossRef]
- Doty, S.L.; Oakley, B.; Xin, G.; Kang, J.W.; Singleton, G.; Khan, Z.; Vajzovic, A.; Staley, J.T. Diazotrophic endophytes of native black cottonwood and willow. Symbiosis 2009, 47, 23–33. [Google Scholar] [CrossRef]
- Kandel, S.L.; Firrincieli, A.; Joubert, P.M.; Okubara, P.A.; Leston, N.D.; McGeorge, K.M.; Mugnozza, G.S.; Harfouche, A.; Kim, S.-H.; Doty, S.L. An In vitro Study of Bio-Control and Plant Growth Promotion Potential of Salicaceae Endophytes. Front. Microbiol. 2017, 8, 386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wullschleger, S.D. Biochemical limitations to carbon assimilation in C3 plants—a retrospective analysis of A/Ci curves from 109 species. J. Exp. Bot. 1993, 44, 907–920. [Google Scholar] [CrossRef]
- Le, T.D.; Dixon, M. Acclimation of photosynthesis in Norway spruce and red oak grown in open-top chambers and subjected to natural drought and elevated CO2. Can. J. For. Res. 1996, 26, 87–94. [Google Scholar]
- Wertin, T.M.; McGuire, M.A.; Teskey, R.O. The influence of elevated temperature, elevated atmospheric CO2 concentration and water stress on net photosynthesis of loblolly pine (Pinus taeda L.) at northern, central and southern sites in its native range. Glob. Chg. Biol. 2010, 16, 2089–2103. [Google Scholar] [CrossRef]
- Hiroke, H.; Kinoshita, T.; Kume, A.; Hanba, Y.T. Responses of leaf photosynthetic traits, water use efficiency, and water relations in five urban shrub tree species under drought stress and recovery. Trees 2021. [Google Scholar] [CrossRef]
- Mosseler, A.; Major, J.E. Biomass and root stem production of a colony-forming willow (Salix interior) on highly disturbed, low fertility sites. Biomass Bioenergy 2015, 74, 202–212. [Google Scholar] [CrossRef]
- Mosseler, A.; Major, J.E. Clonal variation in coppiced and uncoppiced growth, root-sprout stem formation, and biomass partitioning in Salix interior on two highly disturbed site types. Can. J. For. Res. 2022, 52, 148–157. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Clone ID | Provenance | Lat-Long | Measured | |
|---|---|---|---|---|---|
| 2017 | 2018 | ||||
| S. cordata | BIG-C4m | Big Sandy Bay, ON | 44°06′ N–77°43′ W | X | |
| BIG-C5f | Big Sandy Bay, ON | 44°06′ N–77°43′ W | |||
| NOR-C4f | North Beach Park, ON | 43°56′ N–77°31′ W | X | X | |
| OUT-C1m | Outlet Beach, ON | 43°53′ N–77°13′ W | X | ||
| S. discolor | HAW-D4m | Hawkesbury, ON | 45°36′ N–74°36′ W | X | |
| HAW-D5f | Hawkesbury, ON | 45°36′ N–74°36′ W | X | ||
| MON-D1f | Montmagny, QC | 46°58′ N–70°33′ W | X | X | |
| RIC-D2m | Richmond Fen, On | 45°07′ N–75°42′ W | |||
| S. eriocephala | ALL-E2m | Allumette Island, QC | 45°54′ N–77°06′ W | X | |
| FRE-E1f | Fredericton, NB | 45°57′ N–66°38′ W | X | ||
| GRE-E1f | Green River, NB | 47°34′ N–68°19′ W | X | X | |
| WAI-E1m | Wainfleet, ON | 42°55′ N–79°20′ W | |||
| S. interior | LAF-I2f | Ottawa, ON | 45°25′ N–75°41′ W | X | X |
| LAF-I12m | Ottawa, ON | 45°25′ N–75°41′ W | |||
| WAI-I1m | Wainfleet, ON | 42°55′ N–79°20′ W | X | ||
| WAI-I2f | Wainfleet, ON | 42°55′ N–79°20′ W | X | ||
| Source of Variation | df | Vcmax (µmol·m−2·s−1) | Jmax (µmol·m−2·s−1) | TPU (µmol·m−2·s−1) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| MS | VC (%) | p-Value | MS | VC (%) | p-Value | MS | VC (%) | p-Value | ||
| Block | 1 | 97.1 | 1.0 | 0.310 | 483.0 | 0.6 | 0.427 | 6.80 | 2.5 | 0.094 |
| Species | 3 | 1119.7 | 33.3 | <0.001 | 12059.5 | 42.2 | <0.001 | 30.54 | 33.4 | <0.001 |
| CO2 | 1 | 580.6 | 5.8 | 0.016 | 1397.9 | 1.6 | 0.179 | 5.62 | 2.0 | 0.127 |
| Water | 1 | 431.2 | 4.3 | 0.036 | 1461.2 | 1.7 | 0.170 | 29.22 | 10.6 | 0.001 |
| CO2 × species | 3 | 88.0 | 2.6 | 0.422 | 729.1 | 2.6 | 0.414 | 4.26 | 4.6 | 0.155 |
| Water × species | 3 | 265.3 | 7.9 | 0.046 | 2519.2 | 8.8 | 0.027 | 4.43 | 4.8 | 0.142 |
| Water × CO2 | 1 | <0.1 | <0.1 | 0.982 | 159.3 | 0.2 | 0.647 | 0.03 | <0.1 | 0.909 |
| Water × CO2 × species | 3 | 79.4 | 2.4 | 0.468 | 361.0 | 1.3 | 0.697 | 1.90 | 2.1 | 0.491 |
| Error | 47 | 92.2 | 42.9 | 750.9 | 41.1 | 2.33 | 39.9 | |||
| R2 | 0.571 | 0.589 | 0.601 | |||||||
| Source of Variation | df | A600 (µmol·m−2·s−1) | Gwv600 (mmol·m−2·s−1) | WUE600 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| MS | VC (%) | p-Value | MS | VC (%) | p-Value | MS | VC (%) | p-Value | ||
| Block | 1 | 46.0 | 2.0 | 0.148 | 0.10 | 6.8 | 0.001 | 2570.9 | 1.1 | 0.126 |
| Species | 3 | 179.1 | 23.3 | <0.001 | 0.087 | 9.8 | 0.001 | 5307.6 | 7.1 | 0.004 |
| CO2 | 1 | 113.0 | 4.9 | 0.026 | 0.005 | 0.2 | 0.560 | 126.5 | 0.1 | 0.731 |
| Water | 1 | 474.8 | 20.6 | <0.001 | 1.422 | 53.5 | <0.001 | 140223.2 | 62.3 | <0.001 |
| CO2 × species | 3 | 18.6 | 2.4 | 0.461 | 0.012 | 1.4 | 0.452 | 1148.3 | 1.5 | 0.365 |
| Water × species | 3 | 15.7 | 2.0 | 0.536 | 0.010 | 1.1 | 0.555 | 1964.8 | 2.6 | 0.151 |
| Water × CO2 | 1 | 4.6 | 0.2 | 0.644 | 0.041 | 1.5 | 0.094 | 3310.5 | 1.5 | 0.084 |
| Water × CO2 × species | 3 | 8.6 | 1.1 | 0.751 | 0.009 | 1.0 | 0.572 | 1209.5 | 1.6 | 0.342 |
| Error | 47 | 21.3 | 43.4 | 0.014 | 24.7 | 1060.3 | 22.2 | |||
| R2 | 0.566 | 0.753 | 0.778 | |||||||
| Treatment | Organic Matter (%) | Carbon (%) | Nitrogen (%) | Potassium (meq/100 g) | Calcium (meq/100 g) | Magnesium (meq/100 g) | Phosphorus (ppm) |
|---|---|---|---|---|---|---|---|
| NOFERT | 0.67 ± 0.40 a | 0.39± 0.02 a | 0.085 ± 0.009 a | 0.04 ± 0.01 b | 1.21 ± 0.08 a | 0.050 ± 0.004 b | 6.50 ± 0.50 b |
| FERT | 0.63 ± 0.40 a | 0.37 ± 0.02 a | 0.090 ± 0.009 a | 0.16 ± 0.01 a | 1.08 ± 0.08 a | 0.068 ± 0.004 a | 9.50 ± 0.50 a |
| Site | Sodium (%) | Sulfur (%) | pH | C:N Ratio |
|---|---|---|---|---|
| NOFERT | 0.11 ± 0.01 a | 0.020 ± 0.007 a | 6.8 ± 0.1 a | 4.7 ± 0.2 a |
| FERT | 0.11 ± 0.01 a | 0.013 ± 0.007 a | 6.2 ± 0.1 b | 3.9 ± 0.2 a |
| Source of Variation | df | Vcmax (µmol·m−2·s−1) | Jmax (µmol·m−2·s−1) | TPU (µmol·m−2·s−1) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| MS | VC (%) | p-Value | MS | VC (%) | p-Value | MS | VC (%) | p-Value | ||
| Block | 1 | 70.5 | 0.4 | 0.160 | 156.0 | 0.2 | 0.424 | 0.80 | 0.1 | 0.746 |
| Species | 3 | 1366.7 | 24.1 | <0.001 | 7415.7 | 28.1 | <0.001 | 19.58 | 8.7 | 0.063 |
| CO2 | 1 | 255.5 | 1.5 | 0.009 | 1.0 | <0.1 | 0.948 | 2.04 | 0.3 | 0.605 |
| FERT | 1 | 10471.1 | 61.5 | <0.001 | 43202.1 | 54.5 | <0.001 | 102.87 | 15.2 | 0.001 |
| CO2 × species | 3 | 6.5 | 0.1 | 0.904 | 377.1 | 1.4 | 0.208 | 21.36 | 9.4 | 0.048 |
| FERT × species | 3 | 26.9 | 0.5 | 0.514 | 150.8 | 0.6 | 0.599 | 21.10 | 9.3 | 0.050 |
| CO2 × FERT | 1 | 274.0 | 1.6 | 0.007 | 202.6 | 0.3 | 0.362 | 0.45 | 0.1 | 0.809 |
| CO2 × FERT × species | 3 | 41.5 | 0.7 | 0.321 | 195.1 | 0.7 | 0.492 | 10.71 | 4.7 | 0.248 |
| Error | 47 | 34.7 | 9.6 | 239.5 | 14.2 | 7.52 | 52.2 | |||
| R2 | 0.904 | 0.858 | 0.478 | |||||||
| Source of Variation | df | A600 (µmol·m−2·s−1) | Gwv600 (mmol·m−2·s−1) | WUE600 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| MS | VC (%) | p-Value | MS | VC (%) | p-Value | MS | VC (%) | p-Value | ||
| Block | 1 | 3.39 | 0.2 | 0.411 | 0.001 | 0.1 | 0.743 | 321.3 | 0.6 | 0.448 |
| Species | 3 | 155.35 | 20.8 | <0.001 | 0.064 | 14.3 | <0.001 | 3292.5 | 19.3 | 0.002 |
| CO2 | 1 | 22.81 | 1.0 | 0.036 | 0.004 | 0.3 | 0.452 | 442.2 | 0.9 | 0.374 |
| FERT | 1 | 1454.81 | 65.0 | <0.001 | 0.588 | 43.5 | <0.001 | 3373.2 | 6.6 | 0.017 |
| CO2 × species | 3 | 0.84 | 0.1 | 0.916 | 0.023 | 5.0 | 0.042 | 1871.2 | 11.0 | 0.025 |
| FERT × species | 3 | 0.91 | 0.1 | 0.906 | 0.015 | 3.4 | 0.125 | 937.4 | 5.5 | 0.178 |
| CO2 × FERT | 1 | 24.90 | 1.1 | 0.029 | 0.013 | 1.0 | 0.201 | 0.3 | <0.1 | 0.983 |
| CO2 × FERT × species | 3 | 9.71 | 1.3 | 0.131 | 0.027 | 5.9 | 0.022 | 998.7 | 5.8 | 0.156 |
| Error | 47 | 4.92 | 10.3 | 0.008 | 26.5 | 548.5 | 50.3 | |||
| R2 | 0.897 | 0.735 | 0.497 | |||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Major, J.E.; Mosseler, A.; Malcolm, J.W. Assimilation Efficiencies and Gas Exchange Responses of Four Salix Species in Elevated CO2 under Soil Moisture Stress and Fertilization Treatments. Forests 2022, 13, 776. https://doi.org/10.3390/f13050776
Major JE, Mosseler A, Malcolm JW. Assimilation Efficiencies and Gas Exchange Responses of Four Salix Species in Elevated CO2 under Soil Moisture Stress and Fertilization Treatments. Forests. 2022; 13(5):776. https://doi.org/10.3390/f13050776
Chicago/Turabian StyleMajor, John E., Alex Mosseler, and John W. Malcolm. 2022. "Assimilation Efficiencies and Gas Exchange Responses of Four Salix Species in Elevated CO2 under Soil Moisture Stress and Fertilization Treatments" Forests 13, no. 5: 776. https://doi.org/10.3390/f13050776
APA StyleMajor, J. E., Mosseler, A., & Malcolm, J. W. (2022). Assimilation Efficiencies and Gas Exchange Responses of Four Salix Species in Elevated CO2 under Soil Moisture Stress and Fertilization Treatments. Forests, 13(5), 776. https://doi.org/10.3390/f13050776