
Heterogeneous Responses of Alpine Treelines to Climate Warming across the Tibetan Plateau




Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Region and Climate
2.2. Tree Species and Treeline Ecotone
2.3. Data Collection
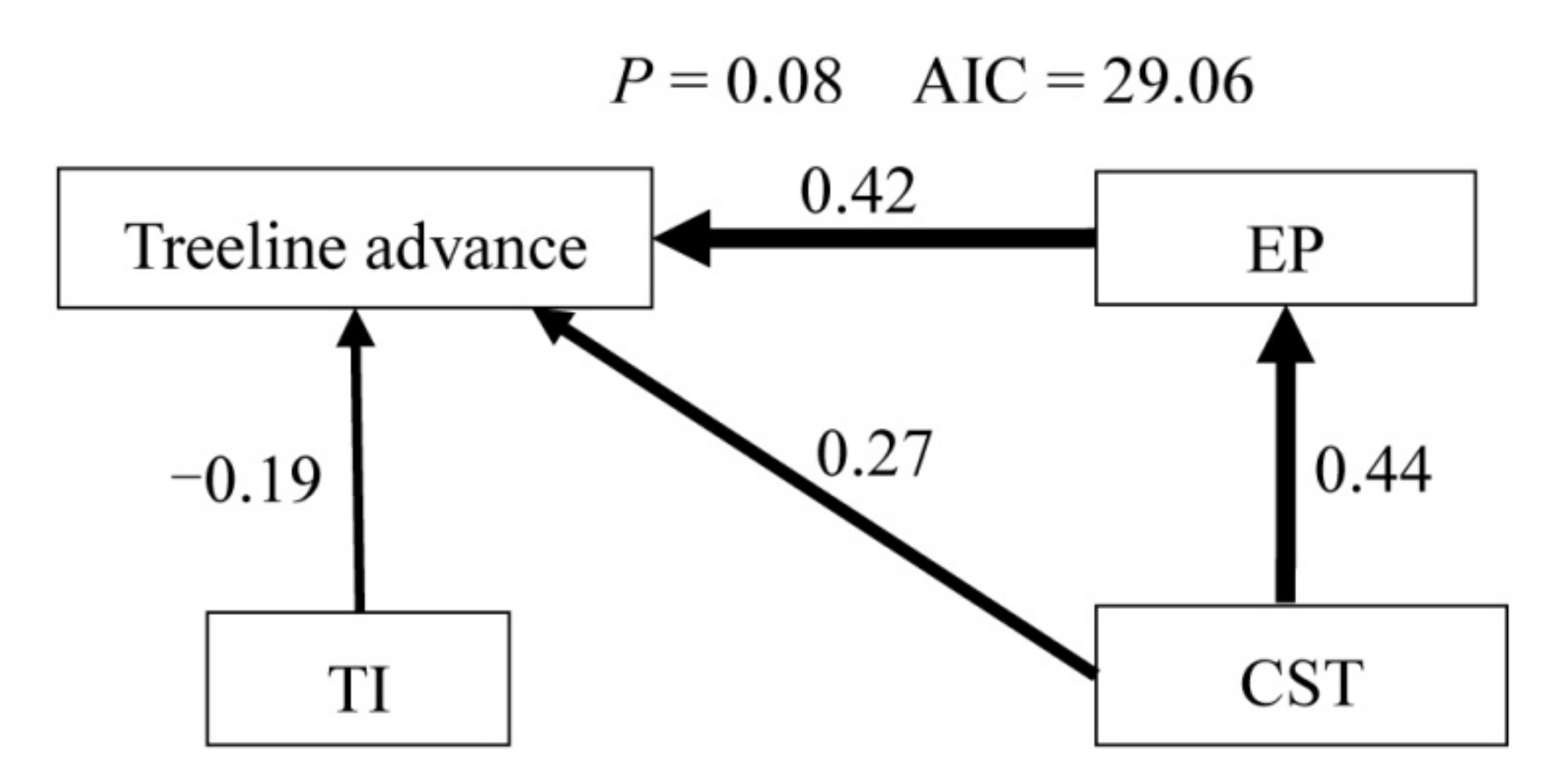
2.4. Data Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Fadrique, B.; Báez, S.; Duque, Á.; Malizia, A.; Blundo, C.; Carilla, J.; Osinaga-Acosta, O.; Malizia, L.; Silman, M.; Farfán-Ríos, W.; et al. Widespread but heterogeneous responses of Andean forests to climate change. Nature 2018, 564, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Liang, E.; Liu, R.; Babst, F.; Camarero, J.J.; Fu, Y.; Pioa, S.; Rossi, S.; Shen, M.; Wang, T.; et al. An earlier start of the thermal growing season enhances tree growth in cold humid areas but not in dry areas. Nat. Ecol. Evol. 2022, 6, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Beloiu, M.; Poursanidis, D.; Tsakirakis, A.; Chrysoulakis, N.; Hoffmann, S.; Lymberakis, P.; Barnias, A.; Kienle, D.; Beierkuhnlein, C. No treeline shift despite climate change over the last 70 Years. For. Ecosyst. 2022, 9, 100002. [Google Scholar] [CrossRef]
- Holtmeier, F.K. Mountain Timberlines: Ecology, Patchiness and Dynamics; Springer: Berlin, Germany, 2009. [Google Scholar]
- Liang, E.; Wang, Y.; Piao, S.; Lu, X.; Camarero, J.J.; Zhu, H.; Zhu, L.; Ellison, A.M.; Ciais, P.; Peñuelas, J. Species interactions slow warming-induced upward shifts of treelines on the Tibetan Plateau. Proc. Natl. Acad. Sci. USA 2016, 113, 4380–4385. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Liang, E.; Wang, Y.; Babst, F.; Camarero, J.J. Mountain treelines climb slowly despite rapid climate warming. Glob. Ecol. Biogeogr. 2021, 30, 305–315. [Google Scholar] [CrossRef]
- Du, H.; Liu, J.; Li, M.H.; Büntgen, U.; Yang, Y.; Wang, L.; Wu, Z.; He, H.S. Warming-induced upward migration of the alpine treeline in the Changbai Mountains, Northeast China. Glob. Change Biol. 2018, 24, 1256–1266. [Google Scholar] [CrossRef]
- Elliott, G.P. Influences of 20th-century warming at the upper tree line contingent on local-scale interactions: Evidence from a latitudinal gradient in the Rocky Mountains, USA. Glob. Change Biol. 2011, 20, 46–57. [Google Scholar] [CrossRef]
- Greenwood, S.; Chen, J.C.; Chen, C.T.; Jump, A.S. Strong topographic sheltering effects lead to spatially complex treeline advance and increased forest density in a subtropical mountain region. Glob. Change Biol. 2014, 20, 3756–3766. [Google Scholar] [CrossRef]
- Sigdel, S.R.; Wang, Y.; Camarero, J.J.; Zhu, H.; Liang, E.; Peñuelas, J. Moisture-mediated responsiveness of treeline shifts to global warming in the Himalayas. Glob. Change Biol. 2018, 24, 5549–5559. [Google Scholar] [CrossRef] [Green Version]
- Körner, C.; Paulsen, J. A world-wide study of high altitude treeline temperatures. J. Biogeogr. 2004, 31, 713–732. [Google Scholar] [CrossRef]
- Macias-Fauria, M.; Johnson, E.A. Warming-induced upslope advance of subalpine forest is severely limited by geomorphic processes. Proc. Natl. Acad. Sci. USA 2013, 110, 8117–8122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camarero, J.J.; Gazol, A.; Sánchez-Salguero, R.; Fajardo, A.; McIntire, E.J.B.; Gutiérrez, E.; Batllori, E.; Boudreau, S.; Carrer, M.; Diez, J.; et al. Global fading of the temperature-growth coupling at alpine and polar treelines. Glob. Change Biol. 2021, 27, 1879–1889. [Google Scholar] [CrossRef] [PubMed]
- Harsch, M.A.; Hulme, P.E.; McGlone, M.S.; Duncan, R.P. Are treelines advancing? a global meta-analysis of treeline response to climate warming. Ecol. Lett. 2009, 12, 1040–1049. [Google Scholar] [CrossRef] [PubMed]
- Liang, E.; Wang, Y.; Eckstein, D.; Luo, T. Little change in the fir tree-line position on the southeastern Tibetan Plateau after 200 years of warming. New Phytol. 2011, 190, 760–769. [Google Scholar] [CrossRef]
- Lett, S.; Dorrepaal, E. Global drivers of tree seedling establishment at alpine treelines in a changing Climate. Funct. Ecol. 2018, 32, 1666–1680. [Google Scholar] [CrossRef] [Green Version]
- Lyu, L.; Zhang, Q.-B.; Pellatt, M.G.; Büntgen, U.; Li, M.H.; Cherubini, P. Drought limitation on tree growth at the northern hemisphere’s highest tree line. Dendrochronologia 2019, 53, 40–47. [Google Scholar] [CrossRef]
- Miehe, G.; Miehe, S.; Vogel, J.; Co, S.; Duo, L. Highest Treeline in the northern hemisphere found in southern Tibet. Mt. Res. Dev. 2007, 27, 169–173. [Google Scholar] [CrossRef] [Green Version]
- Cui, J.; Qin, J.; Sun, H. Population spatial dynamics of Larix potaninii in alpine treeline ecotone in the eastern margin of the Tibetan Plateau, China. Forests 2017, 8, 356. [Google Scholar] [CrossRef]
- Gou, X.; Zhang, F.; Deng, Y.; Ettl, G.J.; Yang, M.; Gao, L.; Fang, K. Patterns and dynamics of tree-line response to climate change in the Eastern Qilian Mountains, Northwestern China. Dendrochronologia 2012, 30, 121–126. [Google Scholar] [CrossRef]
- Lyu, L.; Zhang, Q.-B.; Deng, X.; Mäkinen, H. Fine-scale distribution of treeline trees and the nurse plant facilitation on the eastern Tibetan Plateau. Ecol. Indic. 2016, 66, 251–258. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Mao, Q.; Ren, P.; Sigdel, S.R. Opposite tree-tree interactions jointly drive the natural fir treeline population on the southeastern Tibetan Plateau. Forests 2021, 12, 1417. [Google Scholar] [CrossRef]
- Zhang, F.; Chen, Q.; Gou, X.; Du, M.; Wang, F.; Zhang, J. Climatic control on the growth and regeneration of Juniperus Przewalskii at alpine treeline in the eastern Qilian Mountains, northwest China. Trees 2021, 35, 1085–1097. [Google Scholar] [CrossRef]
- Wang, Y.; Sylvester, S.P.; Lu, X.; Dawadi, B.; Sigdel, S.R.; Liang, E.; Julio Camarero, J. The stability of spruce treelines on the eastern Tibetan Plateau over the last century is explained by pastoral disturbance. For. Ecol. Manag. 2019, 442, 34–45. [Google Scholar] [CrossRef]
- Wang, Y.; Liang, E.; Sigdel, S.R.; Liu, B.; Camarero, J.J. The coupling of treeline elevation and temperature is mediated by non-thermal factors on the Tibetan Plateau. Forests 2017, 8, 109. [Google Scholar] [CrossRef] [Green Version]
- Sigdel, S.R.; Liang, E.; Wang, Y.; Dawadi, B.; Camarero, J.J. Tree-to-tree interactions slow down Himalayan treeline shifts as inferred from tree spatial patterns. J. Biogeogr. 2020, 47, 1816–1826. [Google Scholar] [CrossRef]
- Naudiyal, N.; Wang, J.; Ning, W.; Gaire, N.P.; Peili, S.; Yanqiang, W.; Jiali, H.; Ning, S. Potential distribution of Abies, Picea, and Juniperus species in the sub-alpine forest of Minjiang headwater region under current and future climate scenarios and its implications on ecosystem services supply. Ecol. Indic. 2021, 121, 107131. [Google Scholar] [CrossRef]
- Yao, T.; Thompson, L.; Yang, W.; Yu, W.; Gao, Y.; Guo, X.; Yang, X.; Duan, K.; Zhao, H.; Xu, B.; et al. Different glacier status with atmospheric circulations in Tibetan Plateau and surroundings. Nat. Clim. Change 2012, 2, 663–667. [Google Scholar] [CrossRef]
- Zheng, D. A Study on the Ecogeographic Regional System of China; Chinese Business Press: Beijing, China, 2008. [Google Scholar]
- Fu, T.; Liang, E.; Lu, X.; Gao, S.; Zhang, L.; Zhu, H.; Rossi, S.; Julio Camarero, J.J. Tree growth responses and resilience after the 1950-Zayu-Medog earthquake, southeast Tibetan Plateau. Dendrochronologia 2020, 62, 125724. [Google Scholar] [CrossRef]
- Liu, B.; Liang, E.; Zhu, L. Microclimatic conditions for Juniperus saltuaria treeline in the Sygera Mountains, southeastern Tibetan Plateau. Mt. Res. Dev. 2011, 31, 45–53. [Google Scholar] [CrossRef]
- Dubey, B.; Yadav, R.R.; Singh, J.; Chaturvedi, R. Upward shift of Himalayan pine in Western Himalaya, India. Curr. Sci. 2003, 85, 1135–1136. [Google Scholar]
- Baker, B.B.; Moseley, R.K. Advancing treeline and retreating glaciers: Implications for conservation in Yunnan, China. Arct. Antarct. Alp. Res. 2007, 39, 200–209. [Google Scholar] [CrossRef] [Green Version]
- Miao, N.; Liu, S.; Yu, H.; Shi, Z.; Moermond, T.; Liu, Y. Spatial analysis of remnant tree effects in a secondary Abies-Betula forest on the eastern edge of the Qinghai-Tibetan Plateau, China. For. Ecol. Manag. 2014, 313, 104–111. [Google Scholar] [CrossRef]
- Ran, F.; Liang, Y.; Yang, Y.; Yang, Y.; Wang, X. Spatial-temporal dynamics of an Abies fabri population near the alpine treeline in the Yajiageng area of Gongga Mountains, China. Acta Ecol. Sin. 2014, 34, 6872–6878. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Case, B.; Lu, X.; Ellison, A.M.; Peñuelas, J.; Zhu, H.; Liang, E.; Camarero, J.J. Fire facilitates warming-induced upward shifts of alpine treelines by altering interspecific interactions. Trees 2019, 33, 1051–1061. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Zhang, Y.; Luo, P.; Wang, Q.; Wu, N. Ecological characteristics of a Sabina saltuaria population at timberline on the south facing slope of Baima Snow mountain, southwest China. Chinese. J. Plant Ecol. 2007, 31, 857–864. [Google Scholar] [CrossRef] [Green Version]
- Liang, E.; Shao, X.; Qin, N. Tree-ring based summer temperature reconstruction for the source region of the Yangtze River on the Tibetan Plateau. Glob. Planet. Change 2008, 61, 313–320. [Google Scholar] [CrossRef]
- Zhu, H.; Shao, X.; Yin, Z.; Xu, P.; Xu, Y.; Tian, H. August temperature variability in the southeastern Tibetan Plateau since AD 1385 inferred from tree rings. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2011, 305, 84–92. [Google Scholar] [CrossRef]
- Liu, X.; Zeng, X.; Leavitt, S.W.; Wang, W.; An, W.; Xu, G.; Sun, W.; Wang, Y.; Qin, D.; Ren, J. A 400-year tree-ring δ18O chronology for the southeastern Tibetan Plateau: Implications for inferring variations of the regional hydroclimate. Glob. Planet. Change 2013, 104, 23–33. [Google Scholar] [CrossRef]
- Cheng, W.; Luo, P.; Wu, N. Ecological characteristics of Minjiang fir (Abies faxoniana Rehd et Wild) population near timberline on upper Min River. Chin. J. Appl. Environ. Biol. 2005, 11, 300–303. [Google Scholar] [CrossRef]
- Shao, X.; Huang, L.; Liu, H.; Liang, E.; Fang, X.; Wang, L. Precipitation changes recorded in tree ring over the last 1000 years in the Delingha region of Qinghai province. Sci. China Ser. D Earth Sci. 2004, 34, 145–153. [Google Scholar] [CrossRef]
- Zhu, H.; Zheng, Y.; Shao, X.; Liu, X.; Xu, Y.; Liang, E. Millennial temperature reconstruction based on tree-ring widths of Qilian juniper from Wulan, Qinghai Province. China Sci. Bull. 2008, 53, 3914–3920. [Google Scholar] [CrossRef] [Green Version]
- Fang, K.; Gou, X.; Chen, F.; Peng, J.; D’Arrigo, R.; Wright, W.; Li, M.H. Response of regional tree-line forests to climate change: Evidence from the northeastern Tibetan Plateau. Trees 2009, 23, 1321–1329. [Google Scholar] [CrossRef]
- Shi, C.; Shen, M.; Wu, X.; Cheng, X.; Li, X.; Fan, T.; Li, Z.; Zhang, Y.; Fan, Z.; Shi, F.; et al. Growth response of alpine treeline forests to a warmer and drier climate on the southeastern Tibetan Plateau. Agric. For. Meteorol. 2019, 264, 73–79. [Google Scholar] [CrossRef]
- Harris, I.; Jones, P.D.; Osborn, T.J.; Lister, D.H. Updated high-resolution grids of monthly climatic observations–the CRU TS3. 10 Dataset. Int. J. Climatol. 2014, 34, 623–642. [Google Scholar] [CrossRef] [Green Version]
- Hofgaard, A.; Dalen, L.; Hytteborn, H. Tree recruitment above the treeline and potential for climate-driven treeline change. J. Veg. Sci. 2009, 20, 1133–1144. [Google Scholar] [CrossRef]
- Körner, C. Alpine Treelines: Functional Ecology of the Global High Elevation Tree Limits; Springer: Basel, Switzerland, 2012. [Google Scholar]
- Camarero, J.J.; Gutiérrez, E. Pace and pattern of recent treeline dynamics: Response of ecotones to climatic variability in the Spanish Pyrenees. Clim. Change 2004, 63, 181–200. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: http://www.r-project.org (accessed on 15 April 2022).
- Sadeghi, K.; Marchetti, G.M. Graphical markov models with mixed graphs. R J. 2012, 4, 65–73. [Google Scholar] [CrossRef] [Green Version]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; Heisterkamp, S.; Willigen, B.V.; Ranke, J. nlme: Linear and nonlinear mixed effects models. UTC 2022, 3, 25. [Google Scholar]
- Wang, Y.; Liang, E.; Lu, X.; Camarero, J.J.; Babst, F.; Shen, M.; Peñuelas, J. Warming-induced shrubline advance stalled by moisture limitation on the Tibetan Plateau. Ecography 2021, 44, 1631–1641. [Google Scholar] [CrossRef]
- Lefcheck, J.S. PiecewiseSEM: Piecewise structural equation modelling in R for ecology, evolution, and systematics. Methods Ecol. Evol. 2016, 7, 573–579. [Google Scholar] [CrossRef]
- Shi, H.; Zhou, Q.; He, R.; Zhang, Q.; Dang, H. Climate warming will widen the lagging gap of global treeline shift relative to densification. Agri. For. Meteorol. 2022, 313, 108917. [Google Scholar] [CrossRef]
- Batllori, E.; Camarero, J.J.; Ninot, J.M.; Gutiérrez, E. Seedling recruitment, survival and facilitation in alpine Pinus uncinata tree line ecotones. Implications and potential responses to climate warming. Glob. Ecol. Biogeogr. 2009, 18, 460–472. [Google Scholar] [CrossRef]
- Camarero, J.J.; Gutiérrez, E.; Fortin, M.J. Spatial patterns of plant richness across treeline ecotones in the Pyrenees reveal different locations for richness and tree cover boundaries. Glob. Ecol. Biogeogr. 2006, 15, 182–191. [Google Scholar] [CrossRef]
- Resler, L.M. Geomorphic controls of spatial pattern and process at alpine treeline. Prof. Geogr. 2006, 58, 124–138. [Google Scholar] [CrossRef]
- Shi, H.; Zhou, Q.; Xie, F.; He, N.; He, R.; Zhang, K.; Zhang, Q.; Dang, H. Disparity in elevational shifts of upper species limits in response to recent climate warming in the Qinling Mountains, North-central China. Sci. Total Environ. 2020, 706, 135718. [Google Scholar] [CrossRef]
- Gazol, A.; Moiseev, P.; Camarero, J.J. Changes in plant taxonomic and functional diversity patterns following treeline advances in the South Urals. Plant Ecol. Divers. 2017, 10, 283–292. [Google Scholar] [CrossRef]
- Kullman, L. A review and analysis of factual change on the max rise of the Swedish Scandes treeline, in relation to climate change over the past 100 years. J. Ecol. Nat. Res. 2018, 2, 8. [Google Scholar] [CrossRef]
- Moret, P.; Muriel, P.; Jaramillo, R.; Dangles, O. Humboldt’s tableau physique revisited. Proc. Natl. Acad. Sci. USA 2019, 116, 12889–12894. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Liang, E.; Gričar, J.; Rossi, S.; Čufar, K.; Ellison, A.M. Critical minimum temperature limits xylogenesis and maintains treelines on the southeastern Tibetan Plateau. Sci. Bull. 2017, 62, 804–812. [Google Scholar] [CrossRef] [Green Version]
- Liang, E.; Wang, Y.; Xu, Y.; Liu, B.; Shao, X. Growth variation in Abies georgei var. smithii along altitudinal gradients in the Sygera Mountains, southeastern Tibetan Plateau. Trees 2010, 24, 363–373. [Google Scholar] [CrossRef]
- Wang, Y.; Pederson, N.; Ellison, A.M.; Buckley, H.L.; Case, B.S.; Liang, E.; Camarero, J.J. Increased stem density and competition may diminish the positive effects of warming at alpine treeline. Ecology 2016, 97, 1668–1679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, S.N.; Elliott, G.P.; Schliep, E.M. Seasonal temperature-moisture interactions limit seedling establishment at upper treeline in the Southern Rockies. Ecosphere 2021, 12, e03568. [Google Scholar] [CrossRef]
- Zheng, L.; Shi, P.; Zhou, T.; Hou, G.; Song, M.; Yu, F. Tree regeneration patterns on contrasting slopes at treeline ecotones in eastern Tibet. Forests 2021, 12, 1605. [Google Scholar] [CrossRef]
- Lu, X.; Camarero, J.J.; Liang, E. Threshold responses of juniper tree growth and regeneration to climate warming and drought stress at alpine treeline. Trees 2021, 35, 1081–1083. [Google Scholar] [CrossRef]
- Schickhoff, U.; Bobrowski, M.; Böhner, J.; Bürzle, B.; Chaudhary, R.P.; Gerlitz, L.; Heyken, H.; Lange, J.; Müller, M.; Scholten, T.; et al. Do Himalayan treelines respond to recent climate change? An evaluation of sensitivity indicators. Earth Syst. Dyn. 2015, 6, 245–265. [Google Scholar] [CrossRef]
- Holtmeier, F.K.; Broll, G. Treeline advance-driving processes and adverse factors. Landsc. Online 2007, 1, 1–33. [Google Scholar] [CrossRef]
- Shen, W.; Zhang, L.; Guo, Y.; Luo, T. Causes for treeline stability under climate warming: Evidence from seed and seedling transplant experiments in southeast Tibet. For. Ecol. Manag. 2018, 408, 45–53. [Google Scholar] [CrossRef]
- Körner, C. Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems; Springer: Berlin, Germany, 2003. [Google Scholar]
- Johnson, J.S.; Gaddis, K.D.; Cairns, D.M.; Krutovsky, K.V. Seed dispersal at alpine treeline: An assessment of seed movement within the alpine treeline ecotone. Ecosphere 2017, 8, e01649. [Google Scholar] [CrossRef]
- Zhang, J.; Li, X.; Ren, P.; Leavitt, S.W.; Rossi, S.; Liang, E. Terminal bud size, spring and summer temperatures regulate the timing of height-growth cessation of Smith fir on the southeastern Tibetan Plateau. Agric. For. Meteorol. 2022, 316, 108883. [Google Scholar] [CrossRef]
- Neuschulz, E.L.; Merges, D.; Bollmann, K.; Gugerli, F.; Böhning-Gaese, K. Biotic interactions and seed deposition rather than abiotic factors determine recruitment at elevational range limits of an alpine tree. J. Ecol. 2018, 106, 948–959. [Google Scholar] [CrossRef]
- Chapin, F.S.; Shaver, G.R.; Giblin, A.E.; Nadelhoffer, K.J.; Laundre, J.A. Responses of arctic tundra to experimental and observed changes in climate. Ecology 1995, 76, 694–711. [Google Scholar] [CrossRef]
- Greenwood, S.; Jump, A.S. Consequences of treline shifts for the diveristy and function of high altitude ecosystems. Arct. Antarct. Alp. Res. 2014, 46, 829–840. [Google Scholar] [CrossRef] [Green Version]
- Maher, C.T.; Millar, C.I.; Affleck, D.L.R.; Keane, R.E.; Sala, A.; Tobalske, C.; Larson, A.J.; Nelson, C.R. Alpine treeline ecotones are potential refugia for a montane pine species threatened by bark beetle outbreaks. Ecol. Appl. 2021, 31, e02274. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Correlation (r) |
|---|---|
| Recruitment | 0.39 ** |
| Height growth rate | 0.42 ** |
| CAT | 0.09 |
| CST | 0.49 *** |
| CWT | 0.14 |
| AP | −0.17 |
| SP | −0.12 |
| WP | 0.24 |
| Northness | 0.16 |
| Slope | 0.06 |
| Aspect | −0.11 |
| Vegetation TI | −0.34 ** |
| Disturbance regime | 0.13 |
| EP | 0.54 *** |
| Variables | Partial Correlations with Treeline Shifts | Controlled Variables |
|---|---|---|
| CST | 0.49 *** | CAT, CWT |
| CAT | −0.10 | CST, CWT |
| CWT | −0.08 | CAT, CST |
| AP | −0.19 | SP, WP |
| SP | 0.172 | AP, WP |
| WP | 0.29 | AP, SP |
| RE | 0.31 * | HGR, TI, EP |
| HGR | 0.08 | RE, TI, EP |
| TI | −0.41 ** | RE, HGR, EP |
| EP | 0.41 ** | RE, HGR, TI |
| Fixed Effectssignificance level | AIC | R2m (%) | R2c (%) |
|---|---|---|---|
| CST ** + EP * + TI ** + RE ** + HGR | 493.4 | 57.7 | 66.9 |
| CST ** + EP * + TI ** + RE ** | 503.8 | 56.6 | 63.0 |
| CST * + EP ** + TI | 521.0 | 39.7 | 40.1 |
| CST ** + EP ** + RE ** | 515.5 | 47.4 | 54.6 |
| CST ** + EP ** | 526.9 | 36.8 | 38.4 |
| CST *** + TI | 526.3 | 29.3 | 30.6 |
| CST *** + RE ** | 518.7 | 36.5 | 36.5 |
| CST *** + HGR *** | 512.8 | 37.4 | 37.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Li, D.; Ren, P.; Ram Sigdel, S.; Camarero, J.J. Heterogeneous Responses of Alpine Treelines to Climate Warming across the Tibetan Plateau. Forests 2022, 13, 788. https://doi.org/10.3390/f13050788
Wang Y, Li D, Ren P, Ram Sigdel S, Camarero JJ. Heterogeneous Responses of Alpine Treelines to Climate Warming across the Tibetan Plateau. Forests. 2022; 13(5):788. https://doi.org/10.3390/f13050788
Chicago/Turabian StyleWang, Yafeng, Daihan Li, Ping Ren, Shalik Ram Sigdel, and Jesús Julio Camarero. 2022. "Heterogeneous Responses of Alpine Treelines to Climate Warming across the Tibetan Plateau" Forests 13, no. 5: 788. https://doi.org/10.3390/f13050788
APA StyleWang, Y., Li, D., Ren, P., Ram Sigdel, S., & Camarero, J. J. (2022). Heterogeneous Responses of Alpine Treelines to Climate Warming across the Tibetan Plateau. Forests, 13(5), 788. https://doi.org/10.3390/f13050788