1. Introduction
Xanthoceras sorbifolium Bunge, commonly known as yellow horn, is an oil-rich seed species that belongs to the Sapindaceae family. The oil concentration is about 55–66% in the seed kernel. Historically,
X. sorbifolium seed oil was used for illuminating oil and cooking oil [
1]. Recently, it has been used to produce renewable and environmentally friendly biodiesel and was considered as one of the eight species listed by the national authorities for the production of biodiesel. Moreover, so far, the planted area of yellow horn has increased to 5 × 10
5 ha. [
2]. As an alternative to petrodiesel, the production of
X. sorbifolium has received much attention [
3,
4,
5,
6,
7]. It was reported that yellow horn has experienced extensive management and a ratio of female to male flowers over the past several decades, which has led to low seed yields, i.e., the average yield of mature trees was 670 kg/ha, while 2000 kg/ha was expected under the elite germplasm choice and optimal cultivation and management [
8,
9]. The performance can be improved if the seed from a broad genetic base or from provenance selection is used to match specific site conditions.
Genetic variation within and between populations and seed sources are essential to exploit the improvement potential. Provenance variation, with respect to seed and seedling morphological, anatomical, physiological, and biochemical traits, has been well studied for a number of tree species [
10,
11,
12]. Therefore, a screening test is necessary to take advantage of the naturally available genetic variation so to select the ideal planting material for better productivity.
X. sorbifolium has large range of distribution in North China, extending from 28°34′–47°20′ N, 73°20′–120°25′ E, and at a mean annual rainfall of between 43 and 969 mm [
8]. Since the environmental conditions vary extensively within the natural range of the species, genetic variation among
X. sorbifolium provenances would be expected in some traits. However, to date, little work has been done on the relationship between seed traits and seedling growth performance. In this study, seeds of four provenances from across the natural range of
X. sorbifolium were examined for differences in both seed morphochemical traits and seedling growth and architectural traits in the nursery. The specific aims of this study were to: (1) determine the variation in seed and seedling traits among four different
X. sorbifolium provenances; (2) evaluate the broad-sense heritability and correlations among traits in
X. sorbifolium; (3) assess the correlations among seed and seedling traits and correlations between these traits and environmental conditions; (4) evaluate the four provenances based on these traits. The results will illustrate the relationship between the morphological and chemical traits of seeds from four provenances and help to choose seeds with desired chemical components based on that relationship. Furthermore, this study will offer guidance on how to predict the seedling growth performance according to the seed morphochemical traits.
2. Materials and Methods
2.1. Plant Materials
X. sorbifolium seeds were harvested from four provenances in China, arrayed from east to west: Ar Horqin Banner, Inner Mongolia Autonomous Region (AB); Ongniud Banner, Inner Mongolia Autonomous Region (OB); Shanxian, Henan Province (SX); Mulei, Xinjiang Uygur Autonomous Region (ML). The four provenances span the natural distribution range of the species in China, with the AB, ML, and SX provenances representing the northeastern, northwestern, and southern ranges of distribution of the species, respectively [
13]. The geographic locations and climatic conditions of the different provenances are presented in
Table 1 and
Figure 1. These trees were established by farmers from locally collected seeds from wild sources. In each provenance, to ensure the maximum genetic variation within the population, 2 kg of seeds were collected from all directions of the outer crown of 30 dominant candidate trees. These trees were spaced 30–40 m apart and the seeds were extracted from fruits collected at the peak of the fruit season (July–August) in 2014. All seeds from the same provenance were pooled. Seeds were placed in partially sealed polyethylene bags (100 μm thick, permeable to carbon dioxide and oxygen, yet largely impermeable to moisture), and stored at 2 °C.
2.2. Determination of Seed Characteristics
Determination of seed characteristics was made on each of the 40 seed replicates, drawn at random from the pooled samples from each provenance. Measurement of each trait was performed on the specified number of samples. Five samples of 40 seeds from each provenance were used to measure seed size. Seed length (SL) and width (SW) were measured using a digital vernier caliper (Deli LLC, Ningbo, China). The 1000-seed weight (1000 SW) was determined by measuring the fresh weight (digital balance, UX820H, Shanghai Xiangfan Instruments Ltd., Shanghai, China) of eight samples of 100 seeds, and the 1000-seed weight was calculated. After measuring the morphological parameters, the seeds were oven dried at 80 °C for 4–5 h to determine individual seed dry weight (SDW).
Dried seeds were then crushed into powder, and about 0.2 g of each sample was sieved through a 0.25 mm screen. Determination was carried out after four technical replications of sieving. The samples were wet-digested using the H
2SO
4–H
2O
2 method [
14]. Total nitrogen concentration was determined by the micro-Kjeldehl method (UDK-152, Velp Scientifica, Deer Park, NY, USA) [
15]. Seed crude protein concentration (SCPC) was then estimated by the multiplication of factor 6.25 with N concentration [
16]. Seed soluble sugar concentration (SSSC) was determined on five replicates using an anthrone colorimetry method [
17].
To determine seed oil concentration (SOC), each 100-seed sample was dried to a constant weight at 80 °C, and then pulverized with ball mills. The organic solvent was petroleum ether (boiling point, 60 °C). The kernel oil components were extracted using Soxhlet extraction (Soxtec 8000, FOSS, Hillerod, Denmark) through the following process: boil pulverized seeds at 120 °C for 5 min, leach oils for 1 h, and recover oils for 25 min. The kernel oil weight of five replicates was calculated according to the weight difference between the extraction products and the samples [
3].
Seed mineral concentrations (N, P, K, Ca, Mg, Cu, Zn, Fe, and Mn) of five replicates were determined using atomic-absorption spectrophotometry (Varian SpectrAA 220FS, VARIAN Technology Co., Ltd., Palo Alto, CA, USA). Samples were prepared according to the standard method as described in AOAC [
18].
2.3. Assessment of Seedling Traits
For assessment of the variability in seedling growth parameters, a nursery experiment was performed in a greenhouse at the Beijing Forestry University Forest Science Co., Ltd. (Beijing, China, 40°00′ N, 116°34′ E). Four replicates of 60 seeds per provenance were randomly sampled and arranged in a completely randomized design. Seeds were sown on 2 April 2014 (one seed per container) at a depth of 1–2 cm in a 1050 mL plastic cuboid-shaped container (8 cm diameter × 20 cm deep) filled with a peat: vermiculite 3:1 ratio by volume. Trays were placed on rolling benches under natural light in the greenhouse and rotated weekly to minimize microsite effects. The seedlings were irrigated as needed and grown in temperatures of 24:21 °C (day:night).
Eight seedlings were selected randomly from each replicate, resulting in a total of 32 seedlings per provenance. Thirty days after sowing, seedling height (H) and diameter (D) at ground level were recorded on selected seedlings every 30 days until growth cessation (150 days after sowing) [
19,
20]. In late October, a final destructive sampling was performed. The eight seedlings (per replicate) were excavated and washed gently until free of growing medium and separated into stems and roots. Each plant tissue type was oven-dried at 70 °C for 48 h and then weighed using precision weighing balance to determine dry mass. Total biomass (TB) was calculated as the sum of root and stem dry mass (SB).
Prior to measuring the root dry mass (RB), the roots were scanned on a professional scanner (Epson Expression 1680 Pro, Seiko Epson Corporation, Nagano, Japan) and then analyzed by an image analysis system (WinRHIZO, Regent Instruments Inc., Quebec City, QC, Canada) to determine root length, surface area, and volume (RL, RA, and RB).
2.4. Statistical Analysis
All results were reported as the mean ± standard error (SE). Statistical analysis was performed using SPSS 24.0 (Chicago, IL, USA). The explore function was used to examine data for normality and homogeneity prior to analysis. Analysis of variance (ANOVA) was conducted to test the effects of provenance on the measured parameters. When the ANOVA analysis found significant differences among provenances, Duncan’s test was conducted to detect differences among provenance means (α = 0.05). The Pearson’s correlation coefficient was applied to assess correlations among the different seed and seedling traits. The Vp, representing total phenotypic variance in the provenances, consists of the Vg (genotypic variance) and Ve (environmental variance). The broad sense heritability (H
2) and coefficient of variance (CV) were calculated by the following equations [
21,
22,
23]:
where Vg and Vp are the estimates of the genetic (σ
2g) and residual (σ
2e) variances derived from the expected mean squares of the variance analysis, B is the number of replicates, S is the standard deviation, and
is the mean of the sample data.
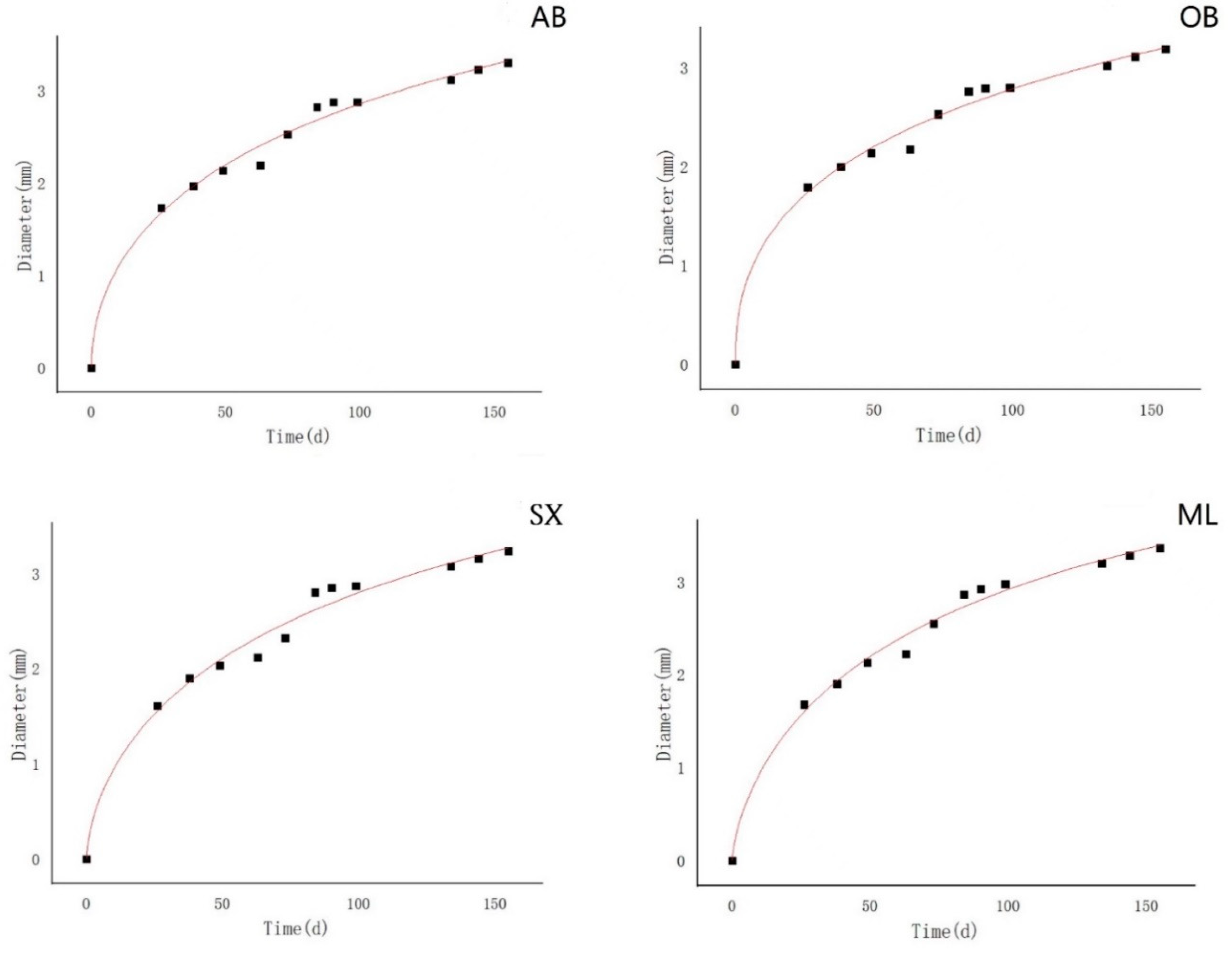
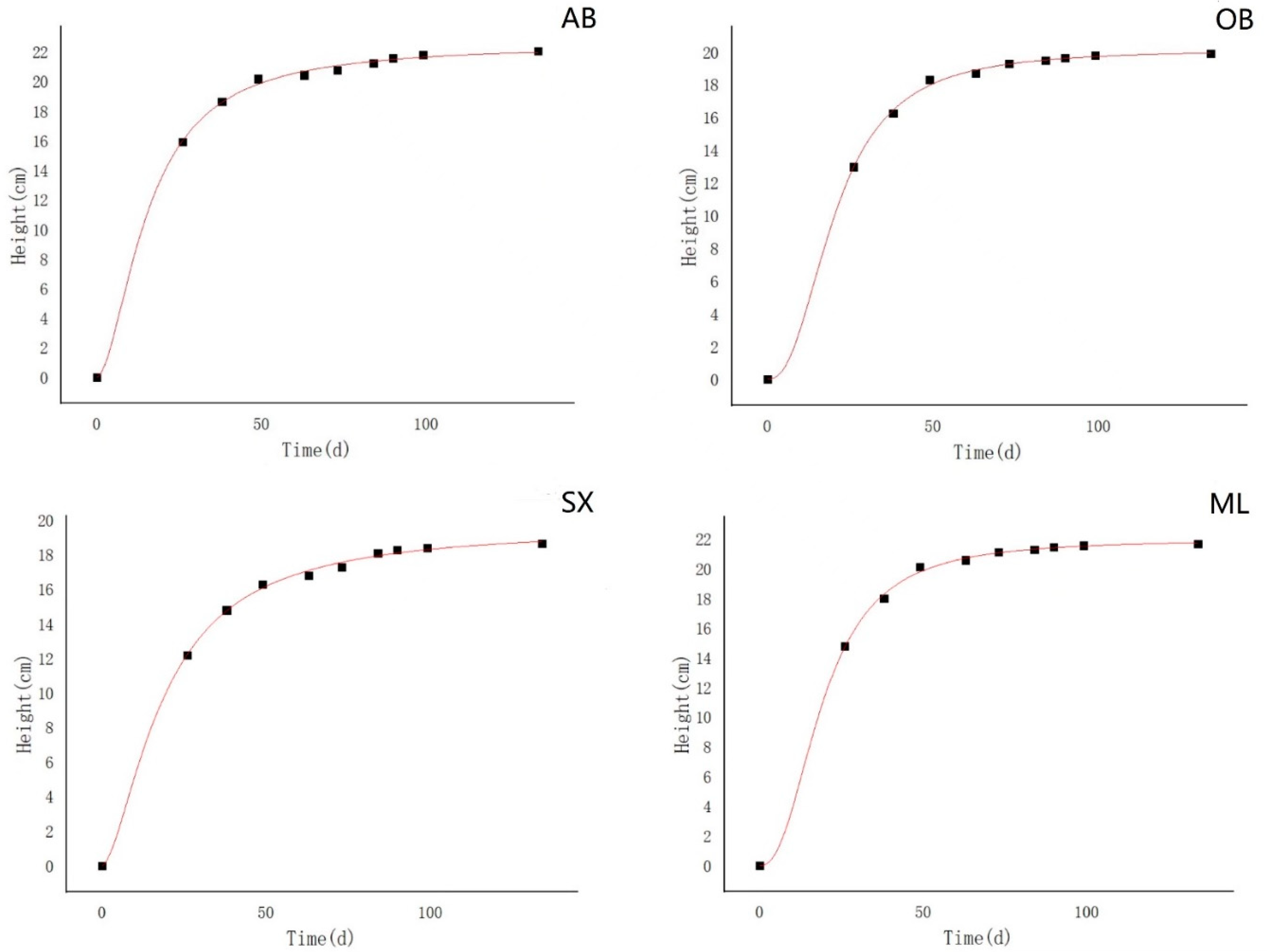
The logistic curve was used to fit the annual growth of the seedling height and ground diameter [
24]. The result was calculated by the following equation:
where y is the growth of the seedlings, t is the growth time (calculated from the sowing day, with the sowing day considered as 0), a and b are the undetermined coefficients, and k is the upper limit value of growth theoretical limitation.
4. Discussion
The distribution of
X. sorbifolium over a wide geographical and climatic range is expected to be reflected in the genetic constitution of its populations. The source of the variation in seed traits has been previously documented in
X. sorbifolium in different provenances [
25,
26,
27]. However, most of the reports neglected the relationship between the seed and the seedling traits. In the present study, a considerable variation in the seed and seedling traits was observed among the provenances. The variation suggested the combined effects of the genotype and the environment, such as geographical and climatic factors.
Seed morphological traits showed significant correlations among themselves (
Table 11), revealing that these traits are independent and genetically controlled. Similar findings were also reported for
Pinus wallichiana [
12],
Acacia catechu Willd. [
28], and
Tectona grandis L.f. [
29]. In addition, this was also supported by the result of the variability analysis. Most of the seed morphological traits were controlled strongly by genotypic factors, while environmental factors played a minor role in shaping these characters (
Table 3), which was in accordance with the trend of the H
2 values. The SW, SDW, and 1000SW were significantly negatively correlated with the mean annual rainfall (
Table 14), indicating that
X. sorbifolium is sensitive to waterlogging. Similar results were also found in
Haloxylon ammodendron [
30]. Both the RB and TB had stronger positive relationships with the mean temperature in January rather than in July. Wulff (1995) also suggested that low temperature played a more important role in increasing the seed mass during seed development and maturation [
31,
32]. Among the provenances, seeds from OB showed the maximum seed length and width (
Table 2). According to the previous reports [
33,
34], this could be attributed to the extremely harsh climate and the long periods of moisture stress that OB experienced. Thus, its seeds needed to store more nutrition for the seedlings to survive after germination.
Among seed chemical components (SCPC, SSSC, and SOC), the H
2 values ranged from 0.188 to 0.345, indicating that these parameters were prone to be influenced by the environment (
Table 3). The SOC was negatively correlated with the seed length, width, and weight (
Table 11), which implied small seeds contain more oil than big ones. A similar result was found in
Jatropha curcas [
35]. The positive relationship of the SSSC and the altitude could be attributed to the fact that important environmental conditions and rainfall change rapidly with elevation [
36], and sunlight intensity increases with altitude, which is helpful for the accumulation of soluble sugar. It is hypothesized that
X. sorbifolium growing at higher altitude level contains more soluble sugar in the seeds. However, further research is needed to elucidate the effect of altitude on this species. The SSSC was significantly negatively correlated with the longitude. This may be because rainfall increases with longitude, and more rainfall and less sunlight lead to low seed soluble sugar concentrations. According to the mineral concentrations, N, P, K, Ca, and Mg were the predominant elements in the seeds, and concentrations of Zn, Fe, and Cu were highly and positively correlated (
Table 11), which were presumably due in part to the charge neutralization produced by the phytate polyanion. This anion is responsible for the bulk of P content in seeds. The positive relationship between Zn and Fe concentrations may result from the fact that Zn and Fe mobilization, uptake, distribution, and accumulation in the plant are controlled by the same genetic and molecular mechanisms [
37].
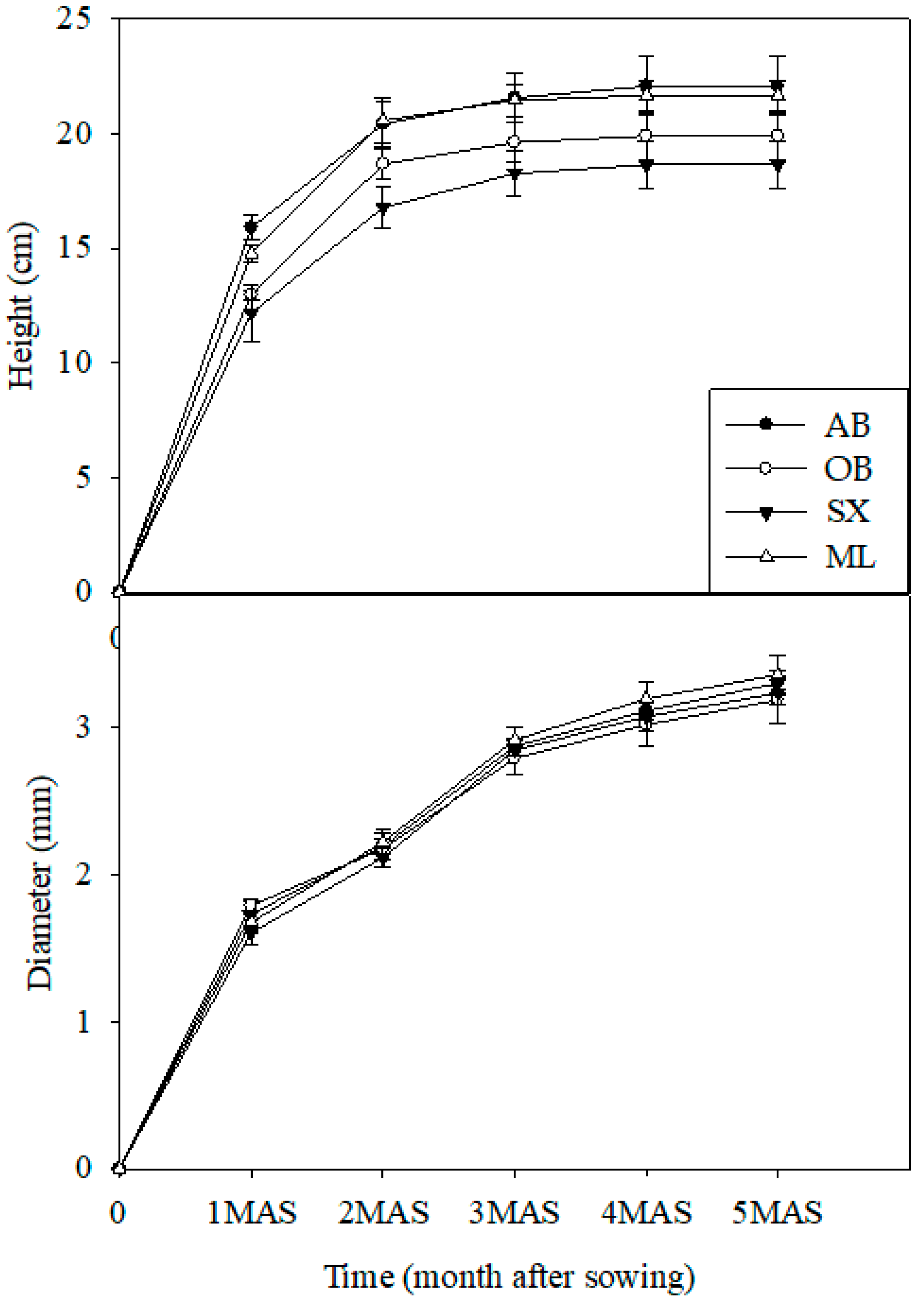
Seedlings of different provenances, when grown under identical environment conditions, often display different patterns of growth [
38,
39,
40]. Provenance variation in the seedling diameter at the nursery stage has been observed for
Dalbergia sissoo Roxb. after 6 months [
41] and for
Tectona grandis after 8 months [
29]. In our study, there was no variation in the seedling diameter throughout the first growing season. Though the seedling height varied significantly at the early stage of growth (1 month and 2 months), the difference disappeared after 3 months. These findings suggest that the expression of genetic potential for seedling growth is species-specific. Meanwhile, the logistic growth model is usually S-shaped, and it includes the emergence stage, seedling stage, rapid growth stage, and late growth stage. In the present study, the seedling diameter and height entered the rapid growth stage quickly probably because the seeds were germinated in advance and the seedlings grew in a greenhouse with ideal temperature and moisture conditions.
Identifying the association between the seed traits and seedling quality parameters would allow us to manipulate the breeding process by selecting seeds. Positive correlations of diameter with the SL, SW, and 1000SW depicted that the provenance which yielded big seeds tended to show greater seedling diameter as compared to the provenance with small seeds (
Table 13). A similar relationship has been obtained in many species [
12,
42]. To a certain extent, the larger seeds are known to have greater cotyledons with more mobilizable carbohydrate reserves and nutrients [
43]; therefore, large seeds could provide an advantage for early seedling establishment. The SSSC showed significantly positive correlations with the RL and RA, and SB, indicating that soluble sugar could provide a nutrition supply for seedling root and stem growth. The advantages provided by the large seeds were also reflected in the seedling traits. In this study, the seedlings developed from large seeds showed a greater survival ratio because of the more water and nutrients absorbed by the larger root diameter and volume. Therefore, among all the seedling traits, the root diameter is considered to be the most effective factor for predicting the future growth state [
19].
Heritability provides an index of the relative roles of heredity and environment in the expression of various traits [
44,
45]. In our study, the SL, SW, SDW, 1000SW, D, and RB had relatively strong heritability (
Table 3 and
Table 6). Therefore, these traits could be improved through clonal selection. On the other hand, these traits are still weakly correlated with environmental and geographic factors. Therefore, future research directions should be: first, carry out further molecular studies to verify genotypic and environmental effects; second, test field growth performance of all provenances more fully in subsequent years across a much broader range of environmental conditions; third, select and analyze additional provenances to get more precise correlation with geographic data.
5. Conclusions
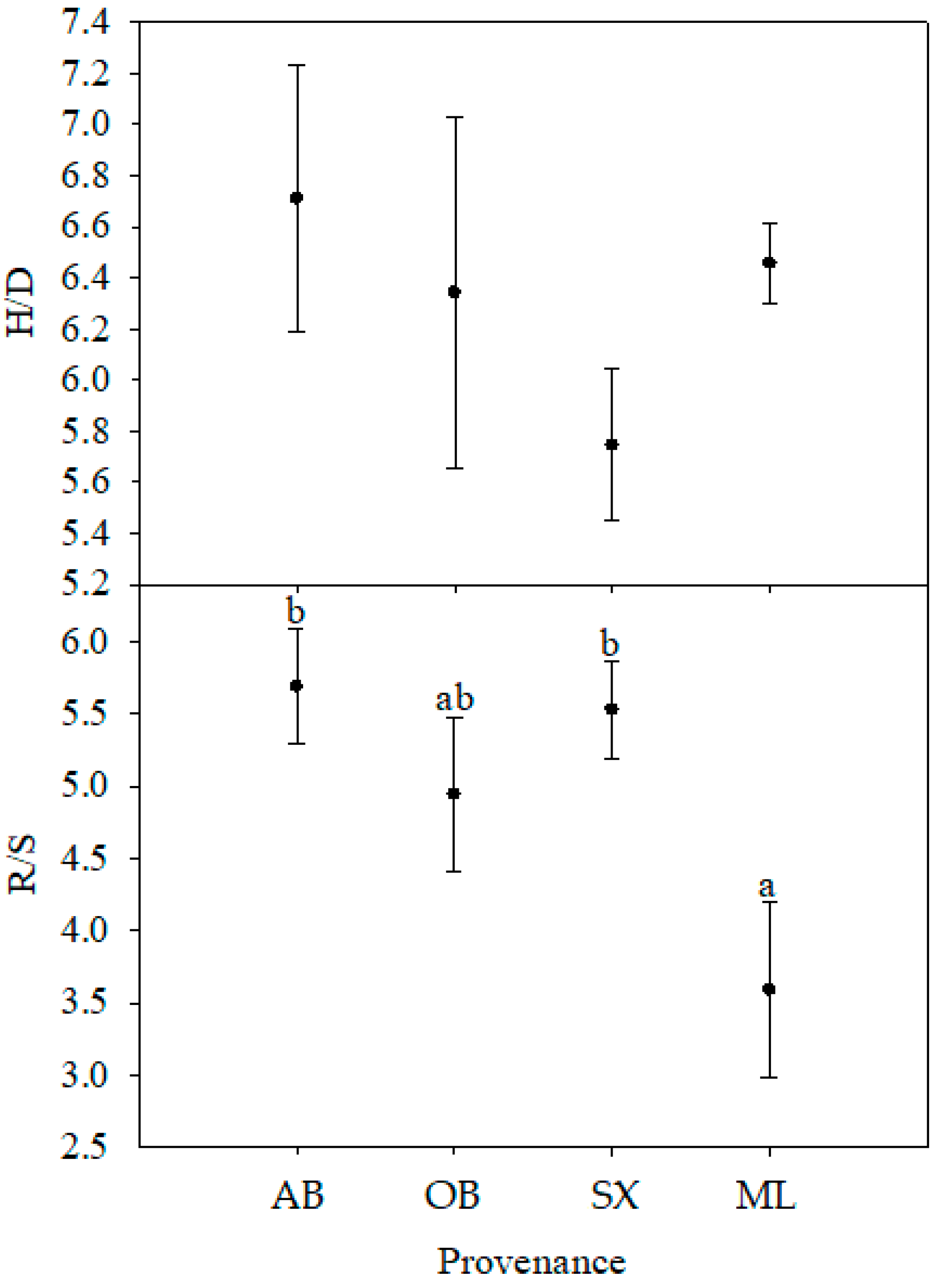
The present study indicated that seed- and seedling-related traits varied considerably among the provenances of X. sorbifolium. Seeds from the OB provenance had the largest size and the lowest SOC and Mg concentrations. The ML provenance demonstrated the maximum values of oil, soluble sugar, P, K, and Mg concentrations, diameter, root length, area, and volume, and comparatively higher values of seed size parameters and biomass. The H2 was high in seed and seedling morphological parameters (seed length, width, weight, seedling diameter, and root biomass). These parameters were also weakly correlated with environmental factors. Seed length, width, and weight were positively correlated with each other. Seed length and width showed negative correlation with the SOC, which meant smaller seeds contain more oil. Additionally, the SSSC was negatively correlated with the longitude and positively correlated with the altitude, indicating that seeds collected from a higher altitude may contain more soluble sugar. Among the four provenances, the ML provenance was recommended as an ideal material for the further improvement program according to the assessment of seed and seedling traits.


 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}