
Variation in the Basic Density of the Tree Components of Gray Alder and Common Alder



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites and Sampling
2.2. Laboratory Analyses
2.3. Statistical Analysis
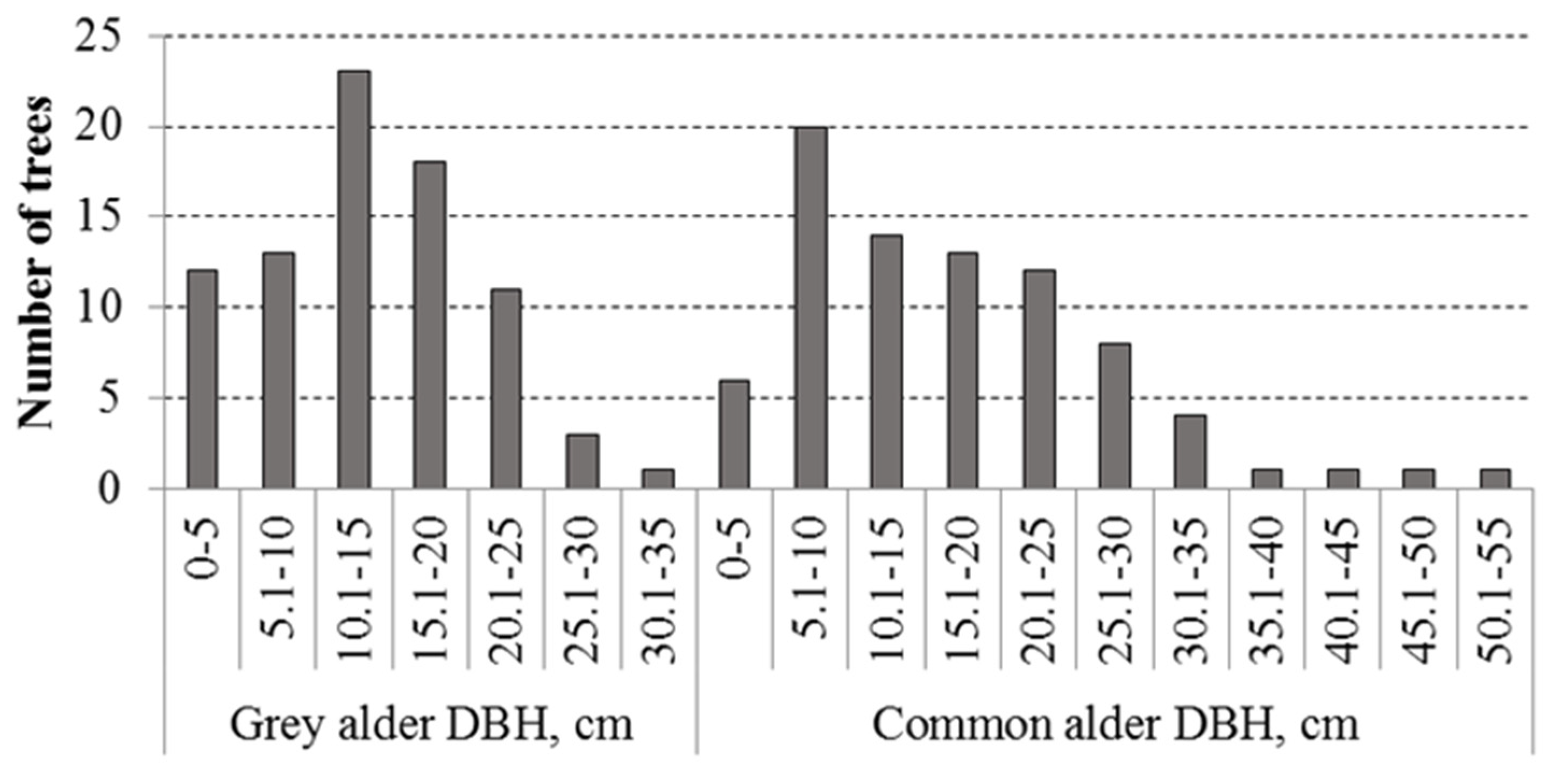
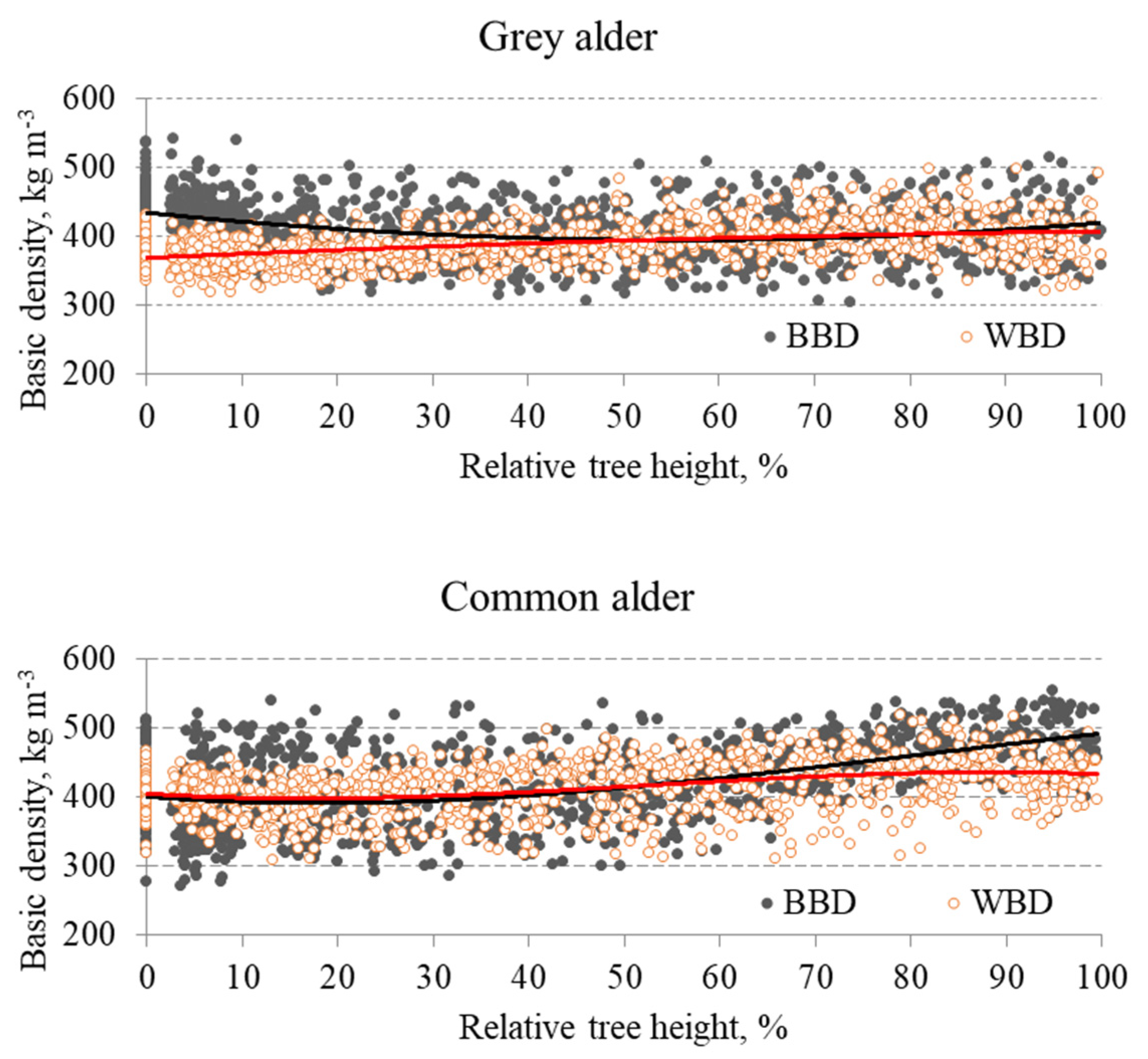
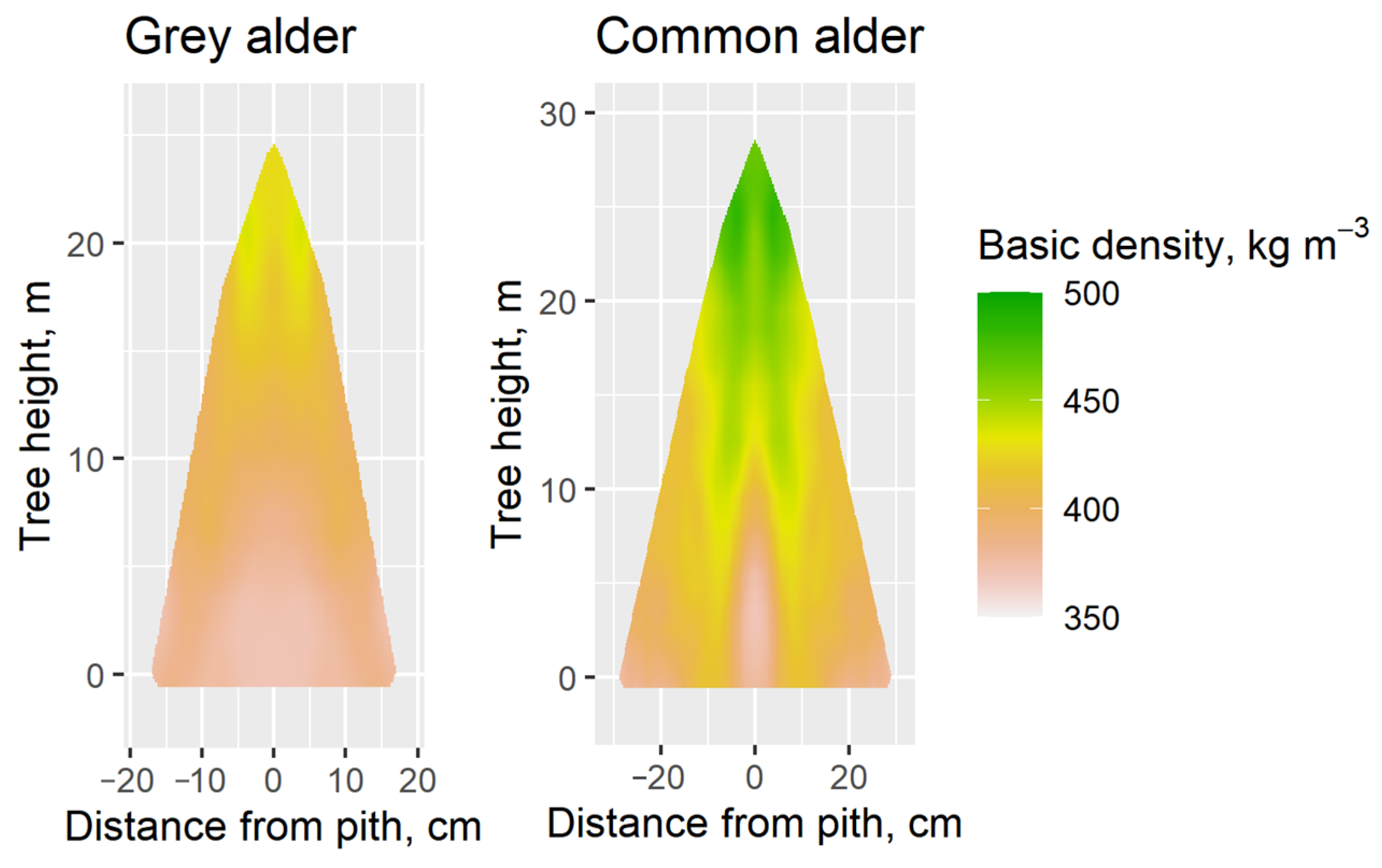
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zobel, B.J.; Jett, J.B. The Importance of Wood Density (Specific Gravity) and Its Component Parts. In Genetics of Wood Production; Springer: Berlin/Heidelberg, Germany, 1995; pp. 78–97. [Google Scholar]
- Saranpää, P. Wood Density and Growth. In Wood Quality and Its Biological Basis; Barnett, J.R., Jeronimidis, G., Eds.; Blackwell Publishing & CRC Press: Boca Raton, FL, USA, 2003; p. 240. [Google Scholar]
- Zink-Sharp, A. The Mechanical Properties of Wood. In Wood Quality and Its Biological Basis; Barnett, J.R., Jeronimidis, G., Eds.; Blackwell Publishing Ltd.: Hoboken, NJ, USA, 2003; pp. 187–211. [Google Scholar]
- Heather, K.; Mackey, G.B.; Lindenmayer, D.B. Re-Evaluation of Forest Biomass Carbon Stocks and Lessons from the World’s Most Carbon-Dense Forests. Proc. Natl. Acad. Sci. USA 2009, 106, 11635–11640. [Google Scholar]
- Pechanec, V.; Štěrbová, L.; Purkyt, J.; Prokopová, M.; Včeláková, R.; Cudlín, O.; Vyvlečka, P.; Cienciala, E.; Cudlín, P. Selected Aspects of Carbon Stock Assessment in Aboveground Biomass. Land 2022, 11, 66. [Google Scholar] [CrossRef]
- Sun, W.; Liu, X. Review on Carbon Storage Estimation of Forest Ecosystem and Applications in China. For. Ecosyst. 2019, 7, 4. [Google Scholar] [CrossRef] [Green Version]
- Vieira, S.A.; Alves, L.F.; Aidar, M.; Araújo, L.S.; Baker, T.; Batista, J.L.F.; Campos, M.C.; Camargo, P.B.; Chave, J.; Delitti, W.B.C.; et al. Estimation of Biomass and Carbon Stocks: The Case of the Atlantic Forest. Biota Neotrop. 2008, 8, 21–29. [Google Scholar] [CrossRef]
- IPCC. IPCC Guidelines for National Greenhouse Gas Inventories; IGES: Hayama, Japan, 2006. [Google Scholar]
- Demol, M.; Calders, K.; Krishna Moorthy, S.M.; van den Bulcke, J.; Verbeeck, H.; Gielen, B. Consequences of Vertical Basic Wood Density Variation on the Estimation of Aboveground Biomass with Terrestrial Laser Scanning. Trees—Struct. Funct. 2021, 35, 671–684. [Google Scholar] [CrossRef]
- Suresh, N.; Stephens, S.A.; Adams, L.; Beck, A.N.; McKinney, A.L.; Varga, T. Extracting Metrics for Three-Dimensional Root Systems: Volume and Surface Analysis from In-Soil X-Ray Computed Tomography Data. JoVE 2016, 110, e53788. [Google Scholar] [CrossRef] [Green Version]
- Sagang, L.B.T.; Momo, S.T.; Libalah, M.B.; Rossi, V.; Fonton, N.; Mofack, G.I.I.; Kamdem, N.G.; Nguetsop, V.F.; Sonké, B.; Ploton, P.; et al. Using Volume-Weighted Average Wood Specific Gravity of Trees Reduces Bias in Aboveground Biomass Predictions from Forest Volume Data. For. Ecol. Manag. 2018, 424, 519–528. [Google Scholar] [CrossRef]
- Longuetaud, F.; Mothe, F.; Santenoise, P.; Diop, N.; Dlouha, J.; Fournier, M.; Deleuze, C. Patterns of Within-Stem Variations in Wood Specific Gravity and Water Content for Five Temperate Tree Species. Ann. For. Sci. 2017, 74, 64. [Google Scholar] [CrossRef]
- Mosedale, J.R.; Charrier, B.; Janin, G. Genetic Control of Wood Colour, Density and Heartwood Ellagitannin Concentration in European Oak (Quercus Petraea and Q. Robur). Forestry 1996, 69, 111–124. [Google Scholar] [CrossRef] [Green Version]
- Moya, R.; Marín, J.D.; Murillo, O.; Leandro, L. Wood Physical Properties, Color, Decay Resistance and Stiffness in Tectona Grandis Clones with Evidence of Genetic Control. Silvae Genet. 2013, 62, 142–152. [Google Scholar] [CrossRef] [Green Version]
- Billard, A.; Bauer, R.; Mothe, F.; Colin, F.; Christine, D.; Longuetaud, F. Vertical Variations in Wood Basic Density for Two Softwood Species. Eur. J. For. Res. 2021, 140, 1401–1416. [Google Scholar] [CrossRef]
- Wiemann, G.; Michael, C.; Williamson, G.B. Wood Specific Gravity Variation with Height and Its Implications for Biomass Estimation; Research Paper, FPL-RP-677; U.S. Department of Agriculture, Forest Service, Forest Products Laboratory: Madison, WI, USA, 2014.
- Woodcock, D.; Shier, A. Wood Specific Gravity and Its Radial Variations: The Many Ways to Make a Tree. Trees—Struct. Funct. 2002, 16, 437–443. [Google Scholar] [CrossRef]
- Liepiņš, J.; Liepiņš, K. Mean Basic Density and Its Axial Variation in Scots Pine, Norway Spruce and Birch Stems. In Proceedings of the Research for Rural Development, Jelgava, Latvia, 17–19 May 2017; Volume 1, pp. 21–27. [Google Scholar]
- Petráš, R.; Mecko, J.; Krupová, D.; Slamka, M.; Pažitný, A. Aboveground Biomass Basic Density of Softwoods Tree Species. Wood Res. 2019, 64, 205–212. [Google Scholar]
- Repola, J. Models for Vertical Wood Density of Scots Pine, Norway Spruce and Birch Stems, and Their Application to Determine Average Wood Density. Silva Fenn. 2006, 40, 673–685. [Google Scholar] [CrossRef] [Green Version]
- Billard, A.; Bauer, R.; Mothe, F.; Jonard, M.; Colin, F.; Longuetaud, F. Improving Aboveground Biomass Estimates by Taking into Account Density Variations between Tree Components. Ann. For. Sci. 2020, 77, 1–18. [Google Scholar] [CrossRef]
- Dibdiakova, J.; Vadla, K. Basic Density and Moisture Content of Coniferous Branches and Wood in Northern Norway. In Proceedings of the EPJ Web of Conferences, Maastricht, The Netherlands, 17–20 April 2012; Volume 33. [Google Scholar]
- Gryc, V.; Horáček, P.; Šezingerová, J.; Vavrčík, H. Basic Density of Spruce Wood, Wood with Bark, and Bark of Branches in Locations in the Czech Republic. Wood Res. 2011, 56, 23–32. [Google Scholar]
- Miles, P.D. Specific Gravity and Other Properties of Wood and Bark for 156 Tree Species Found in North America; US Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2009; Volume 38.
- Erickson, J.R. The Moisture Content and Specific Gravity of the Bark and Wood of Northern Pulpwood Species; Research Note NC-141; US Department of Agriculture, Forest Service, North Central Forest Experiment Station: Saint Paul, MN, USA, 1972; pp. 1–3.
- Official Statistics of Latvia. Inventoried Forest Area and Standing Timber by Main TREE Species. Available online: https://data.stat.gov.lv/pxweb/en/OSP_PUB/START__NOZ__ME__MEP/MEP060/ (accessed on 12 November 2022).
- Ahti, T.; Hämet-Ahti, L.; Jalas, J. Vegetation Zones and Their Sections in Northwestern Europe. Ann. Bot. Fenn. 1968, 5, 169–211. [Google Scholar]
- Official Statistics of Latvia. Forest Regeneration and Afforestation. Available online: https://stat.gov.lv/en/statistics-themes/business-sectors/forestry/8918-forest-regeneration-and-afforestation?themeCode=ME (accessed on 12 November 2022).
- FAO. Global Forest Resources Assessment 2020: Main Report; FAO: Rome, Italy, 2020; p. 165. [Google Scholar]
- Claessens, H.; Oosterbaan, A.; Savill, P.; Rondeux, J. A Review of the Characteristics of Black Alder (Alnus glutinosa (L.) Gaertn.) and Their Implications for Silvicultural Practices. Forestry 2010, 83, 163–175. [Google Scholar] [CrossRef] [Green Version]
- Salca, E. Black Alder (Alnus glutinosa L.)—A Resource for Value-Added Products in Furniture Industry Under European Screening. Curr. For. Rep. 2019, 5, 41–54. [Google Scholar] [CrossRef]
- Aosaar, J.; Varik, M.; Uri, V. Biomass Production Potential of Grey Alder (Alnus incana (L.) Moench.) in Scandinavia and Eastern Europe: A Review. Biomass Bioenergy 2012, 45, 11–26. [Google Scholar] [CrossRef]
- Rytter, L.; Rytter, R.M. Growth and Carbon Capture of Grey Alder (Alnus incana (L.) Moench.) under North European Conditions–Estimates Based on Reported Research. For. Ecol. Manag. 2016, 373, 56–65. [Google Scholar] [CrossRef]
- Arhipova, N.; Gaitnieks, T.; Donis, J.; Stenlid, J.; Vasaitis, R. Decay, Yield Loss and Associated Fungi in Stands of Grey Alder (Alnus incana) in Latvia. Forestry 2011, 84, 337–348. [Google Scholar] [CrossRef]
- Uri, V.; Kukumägi, M.; Aosaar, J.; Varik, M.; Becker, H.; Soosaar, K.; Morozov, G.; Ligi, K.; Padari, A.; Ostonen, I.; et al. Carbon Budgets in Fertile Grey Alder (Alnus incana (L.) Moench.) Stands of Different Ages. For. Ecol. Manag. 2017, 396, 55–67. [Google Scholar] [CrossRef]
- Johansson, T. Stem Volume Equations and Basic Density for Grey Alder and Common Alder in Sweden. Forestry 2005, 78, 249–262. [Google Scholar] [CrossRef]
- Chow, P.; Rolfe, G.L.; Bajwa, D.S. Stem Chemical Compositions of Juvenile Elaeagnus and Alnus Species. Can. J. Bot. 1999, 77, 1398–1400. [Google Scholar]
- Klevinska, V.; Bikova, T. Comparison of the Properties of Black and Grey Juvenile Alder Wood. Holz Als Roh-Und Werkst. 1999, 57, 246. [Google Scholar] [CrossRef]
- Aosaar, J.; Varik, M.; Lõhmus, K.; Uri, V. Stemwood Density in Young Grey Alder (Alnus incana (L.) Moench) and Hybrid Alder (Alnus Hybrida a. Br.) Stands Growing on Abandoned Agricultural Land. Balt For. 2011, 17, 89–94. [Google Scholar]
- Hakkila, P. Basic Density, Bark Percentage and Dry Matter Content of Grey Alder (Alnus incana). Commun. Inst. For. Fenn. 1970, 71, 31. [Google Scholar]
- Liepiņš, J.; Liepiņš, K.; Lazdiņš, A. Equations for Estimating the Above- and Belowground Biomass of Grey Alder (Alnus incana (L.) Moench.) and Common Alder (Alnus glutinosa L.) in Latvia. Scand. J. For. Res. 2021, 36, 389–400. [Google Scholar] [CrossRef]
- Zālītis, P.; Jansons, J. Latvian Forest Typology and Its Origin; DU Akadēmiskais Apgāds Saule: Daugavpils, Latvia, 2013; p. 167. [Google Scholar]
- Kraft, G. Beiträge Zur Lehre von Den Durchforstungen, Schlagstellungen Und Lichtungshieben; Klindworth: Hannover, Germany, 1884; p. 147. [Google Scholar]
- FAO. Manual for Visual Assessment of Forest Crown Condition. In Food and Agriculture Organization of the United Nations, Pristina; FAO: Rome, Italy, 2014; p. 17. [Google Scholar]
- Liepiņš, J.; Lazdiņš, A.; Liepiņš, K. Equations for Estimating Above- and Belowground Biomass of Norway Spruce, Scots Pine, Birch Spp. and European Aspen in Latvia. Scand J. For. Res. 2018, 33, 58–70. [Google Scholar] [CrossRef]
- Ilic, J.; Boland, D.; McDonald, M.; Downes, G.; Blakemore, P. Woody Density Phase 1—State of Knowledge; NCAS Technical Report No.18; NCAS: Canberra, Australia, 2000; p. 218. [Google Scholar]
- Jagodziński, A.M.; Dyderski, M.K.; Gęsikiewicz, K.; Horodecki, P. Consequences of Different Sample Drying Temperatures for Accuracy of Biomass Inventories in Forest Ecosystems. Sci. Rep. 2020, 10, 16009. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Weiskittel, A.R.; MacFarlane, D.W.; Radtke, P.J.; Affleck, D.L.R.; Temesgen, H.; Woodall, C.W.; Westfall, J.A.; Coulston, J.W. A Call to Improve Methods for Estimating Tree Biomass for Regional and National Assessments. J. For. 2015, 113, 414–424. [Google Scholar] [CrossRef] [Green Version]
- Dietz, P. Dichte Und Rindengehalt von Industrieholz. Holz Als Roh-Und Werkst. 1975, 33, 135–141. [Google Scholar] [CrossRef]
- Cīrule, D.; Alksne, A.; Lavnikoviča, I.; Antons, A.; Pavlovičs, G.; Dolacis, J. Comparison of the Physical Properties of Grey Alder (Alnus incana (L.) Moench) and Black Alder (Alnus glutinosa (L.) Gaertn.) Wood in Latvia and Elsewhere. Ann. Wars. Univ. Life Sci. For. Wood Technol. 2008, 63, 129–132. [Google Scholar]
- Miezīte, O.; Dreimanis, A. Investigations of Grey Alder (Alnus incana (L.) Moench) Biomass. In Proceedings of the Research for Rural Development 2006, International Scientific Cinference Proceedings, Jelgava, Latvia, 19–22 May 2006; pp. 271–275. [Google Scholar]
- Köster, K.; Metslaid, M.; Engelhart, J.; Köster, E. Dead Wood Basic Density, and the Concentration of Carbon and Nitrogen for Main Tree Species in Managed Hemiboreal Forests. For. Ecol. Manag. 2015, 354, 35–42. [Google Scholar] [CrossRef]
- Soge, A.O.; Popoola, O.I.; Adetoyinbo, A.A. Detection of Wood Decay and Cavities in Living Trees: A Review. Can. J. For. Res. 2020, 51, 937–947. [Google Scholar] [CrossRef]
- Johansson, T. Biomass Equations for Hybrid Larch Growing on Farmland. Biomass Bioenergy 2013, 49, 152–159. [Google Scholar] [CrossRef]
- Repola, J. Biomass Equations for Birch in Finland. Silva Fenn. 2008, 42, 605–624. [Google Scholar] [CrossRef]
- Lopes, O.P.; Zannato, B.; Barbosa, J.d.A.; Angeles, G.; Longui, E.L. Hydraulic Structure and Wood Density from Root to the Crown in in Three Native Brazilian Tree Species. Sci. Electron. Arch. 2022, 15. [Google Scholar] [CrossRef]
- Longui, E.L.; Galão, A.T.D.; Rajput, K.S.; de Melo, A.C.G. Anatomical Investigation of Root, Stem and Branch Wood in 10-Year-Old Inga Laurina in the Context of Anatomical Adaptation to Hydraulic and Mechanical Stresses. Anales Biología 2018, 40, 31–39. [Google Scholar] [CrossRef]
- Schuldt, B.; Leuschner, C.; Brock, N.; Horna, V. Changes in Wood Density, Wood Anatomy and Hydraulic Properties of the Xylem along the Root-to-Shoot Flow Path in Tropical Rainforest Trees. Tree Physiol. 2013, 33, 161–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, Z.-X.; Zhang, S.-B.; Hao, G.-Y.; Ferry Slik, J.W.; Cao, K.-F. Hydraulic Conductivity Traits Predict Growth Rates and Adult Stature of 40 Asian Tropical Tree Species Better than Wood Density. J. Ecol. 2012, 100, 732–741. [Google Scholar] [CrossRef]
- Zanne, A.E.; Westoby, M.; Falster, D.S.; Ackerly, D.D.; Loarie, S.R.; Arnold, S.E.J.; Coomes, D.A. Angiosperm Wood Structure: Global Patterns in Vessel Anatomy and Their Relation to Wood Density and Potential Conductivity. Am. J. Bot. 2010, 97, 207–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobsen, A.L.; Agenbag, L.; Esler, K.J.; Pratt, R.B.; Ewers, F.W.; Davis, S.D. Xylem Density, Biomechanics and Anatomical Traits Correlate with Water Stress in 17 Evergreen Shrub Species of the Mediterranean-Type Climate Region of South Africa. J. Ecol. 2007, 95, 171–183. [Google Scholar] [CrossRef]
- Houston Durrant, T.; de Rigo, D.; Caudullo, G. Alnus Incana in Europe: Distribution, Habitat, Usage and Threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publications Office of the EU: Luxembourg, 2016; pp. 66–67. [Google Scholar]
- Houston Durrant, T.; de Rigo, D.; Caudullo, G. Alnus Glutinosa in Europe: Distribution, Habitat, Usage and Threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publications Office of the EU: Luxembourg, 2016; pp. 64–65. [Google Scholar]
- Winter, H. Specific Gravity of Inner and Outer Beech Bark. Eur. J. Wood Wood Prod. 2020, 78, 413–416. [Google Scholar] [CrossRef] [Green Version]
- MacFarlane, D.W. Functional Relationships Between Branch and Stem Wood Density for Temperate Tree Species in North America. Front. For. Glob. Chang. 2020, 3, 63. [Google Scholar] [CrossRef]
- Dahle, G.A.; Grabosky, J.C. Variation in Modulus of Elasticity (E) along Acer platanoides L. (Aceraceae) Branches. Urban For. Urban Green 2010, 9, 227–233. [Google Scholar] [CrossRef]
- Tsai, C.-C.; Hung, L.-F.; Chien, C.-T.; Chen, S.-J.; Huang, Y.-S.; Kuo-Huang, L.-L. Biomechanical Features of Eccentric Cambial Growth and Reaction Wood Formation in Broadleaf Tree Branches. Trees—Struct. Funct. 2012, 26, 1585–1595. [Google Scholar] [CrossRef]
- Ramos-Rivera, J.; Rahardjo, H.; Tsen-Tieng, D.L.; Xuefeng, N.; King, F.Y. Mechanical Response of the Real Tree Root Architecture under Lateral Load. Can. J. For. Res. 2020, 50, 595–607. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gray Alder | Common Alder | |||||||
|---|---|---|---|---|---|---|---|---|
| Mean | Std | Min | Max | Mean | Std | Min | Max | |
| Aboveground | ||||||||
| N a = 81 | N = 81 | |||||||
| DBH b, cm | 13.5 | 6.7 | 2.2 | 30.6 | 16.6 | 10.6 | 2.2 | 53.9 |
| H c, m | 15.1 | 6 | 3.9 | 25.6 | 16.4 | 8.2 | 2.8 | 28.6 |
| Age d | 29.4 | 14.9 | 4 | 64 | 37.5 | 29.3 | 4 | 115 |
| Belowground | ||||||||
| N = 28 | N = 27 | |||||||
| DBH, cm | 11.9 | 7.5 | 2.2 | 29.1 | 13.5 | 7.3 | 2.3 | 28.5 |
| H, m | 13.5 | 7.1 | 3.9 | 25.6 | 14.3 | 7.2 | 2.8 | 26.2 |
| Age | 24.2 | 13.9 | 4 | 49 | 31.0 | 25.3 | 4 | 102 |
| Gray Alder | Common Alder | p-Value of Inter-Species Differences | |
|---|---|---|---|
| Stem | |||
| Stem with bark (SD) | 384.45 ± 2.05 | 405.53 ± 2.86 | <0.001 |
| Stem wood | 382.54 ± 2.10 | 403.37 ± 3.52 | <0.001 |
| Stem bark | 403.25 ± 3.80 | 414.23 ± 5.31 | 0.218 |
| Branch | |||
| Average | 407.97 ± 2.61 | 435.38 ± 2.84 | <0.001 |
| Lower (B1) | 424.78 ± 2.73 | 444.57 ± 2.75 | <0.001 |
| Middle (B2) | 403.19 ± 2.78 | 428.60 ± 3.63 | <0.001 |
| Upper (B3) | 395.93 ± 3.48 | 432.97 ± 3.32 | <0.001 |
| Stump | 413.88 ± 7.43 | 381.26 ± 8.01 | 0.011 |
| Coarse roots | |||
| Average | 358.53 ± 4.62 | 292.93 ± 8.41 | <0.001 |
| Thicker end (CR1) | 396.15 ± 4.49 | 330.24 ± 11.03 | <0.001 |
| Middle (CR2) | 366.87 ± 5.19 | 295.41 ± 11.71 | <0.001 |
| Smaller end (CR3) | 312.59 ± 11.19 | 253.15 ± 10.50 | 0.001 |
| Small roots | |||
| Average | 317.40 ± 3.93 | 230.55 ± 4.73 | <0.001 |
| Thicker end (SR1) | 317.76 ± 7.59 | 242.79 ± 9.15 | <0.001 |
| Middle (SR2) | 322.76 ± 6.86 | 225.59 ± 6.52 | <0.001 |
| Smaller end (SR3) | 321.11 ± 6.58 | 218.54 ± 7.51 | <0.001 |
| Gray Alder Aboveground Components | |||||||
| Age * | DBH | H | SD | WBD | BBD | BR | |
| Age | 1 | 0.902 | 0.908 | 0.31 | 0.222 | 0.578 | 0.365 |
| DBH | <0.001 | 1 | 0.94 | 0.268 | 0.181 | 0.585 | 0.419 |
| H | <0.001 | <0.001 | 1 | 0.288 | 0.206 | 0.56 | 0.408 |
| SD | 0.005 | 0.016 | 0.009 | 1 | 0.977 | 0.458 | 0.512 |
| WBD | 0.046 | 0.106 | 0.065 | <0.001 | 1 | 0.293 | 0.45 |
| BBD | <0.001 | <0.001 | <0.001 | <0.001 | 0.008 | 1 | 0.499 |
| BR | 0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 1 |
| Gray Alder Belowground Components | |||||||
| Age | DBH | H | STP | CR | SR | ||
| Age | 1 | 0.934 | 0.943 | −0.072 | −0.178 | 0.008 | |
| DBH | <0.001 | 1 | 0.95 | −0.248 | −0.193 | −0.013 | |
| H | <0.001 | <0.001 | 1 | −0.207 | −0.175 | −0.055 | |
| STP | 0.731 | 0.23 | 0.321 | 1 | 0.53 | 0.448 | |
| CR | 0.396 | 0.353 | 0.404 | 0.007 | 1 | 0.375 | |
| SR | 0.969 | 0.952 | 0.793 | 0.026 | 0.065 | 1 | |
| Common Alder Aboveground Components | |||||||
| Age | DBH | H | SD | WBD | BBD | BR | |
| Age | 1 | 0.874 | 0.874 | 0.561 | 0.715 | −0.794 | 0.659 |
| DBH | <0.001 | 1 | 0.941 | 0.506 | 0.646 | −0.712 | 0.698 |
| H | <0.001 | <0.001 | 1 | 0.513 | 0.669 | −0.756 | 0.659 |
| SD | <0.001 | <0.001 | <0.001 | 1 | 0.959 | −0.302 | 0.784 |
| WBD | <0.001 | <0.001 | <0.001 | <0.001 | 1 | −0.528 | 0.808 |
| BBD | <0.001 | <0.001 | <0.001 | 0.006 | <0.001 | 1 | −0.377 |
| BR | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.001 | 1 |
| Common Alder Belowground Components | |||||||
| Age | DBH | H | STP | CR | SR | ||
| Age | 1 | 0.837 | 0.757 | 0.169 | 0.425 | 0.047 | |
| DBH | <0.001 | 1 | 0.834 | 0.072 | 0.299 | −0.198 | |
| H | <0.001 | <0.001 | 1 | 0.274 | 0.577 | −0.113 | |
| STP | 0.429 | 0.737 | 0.195 | 1 | 0.396 | 0.137 | |
| CR | 0.039 | 0.155 | 0.004 | 0.057 | 1 | 0.243 | |
| SR | 0.829 | 0.351 | 0.598 | 0.52 | 0.25 | 1 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liepiņš, K.; Liepiņš, J.; Ivanovs, J.; Bārdule, A.; Jansone, L.; Jansons, Ā. Variation in the Basic Density of the Tree Components of Gray Alder and Common Alder. Forests 2023, 14, 135. https://doi.org/10.3390/f14010135
Liepiņš K, Liepiņš J, Ivanovs J, Bārdule A, Jansone L, Jansons Ā. Variation in the Basic Density of the Tree Components of Gray Alder and Common Alder. Forests. 2023; 14(1):135. https://doi.org/10.3390/f14010135
Chicago/Turabian StyleLiepiņš, Kaspars, Jānis Liepiņš, Jānis Ivanovs, Arta Bārdule, Līga Jansone, and Āris Jansons. 2023. "Variation in the Basic Density of the Tree Components of Gray Alder and Common Alder" Forests 14, no. 1: 135. https://doi.org/10.3390/f14010135
APA StyleLiepiņš, K., Liepiņš, J., Ivanovs, J., Bārdule, A., Jansone, L., & Jansons, Ā. (2023). Variation in the Basic Density of the Tree Components of Gray Alder and Common Alder. Forests, 14(1), 135. https://doi.org/10.3390/f14010135