

Potential and Constraints on In Vitro Micropropagation of Juniperus drupacea Labill.
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material—Explants Sterilization—Culture Establishment
2.2. Shoot Regeneration, Multiplication, and Elongation
2.3. In Vitro Rooting of Shoots
2.4. Statistical Analysis
3. Results
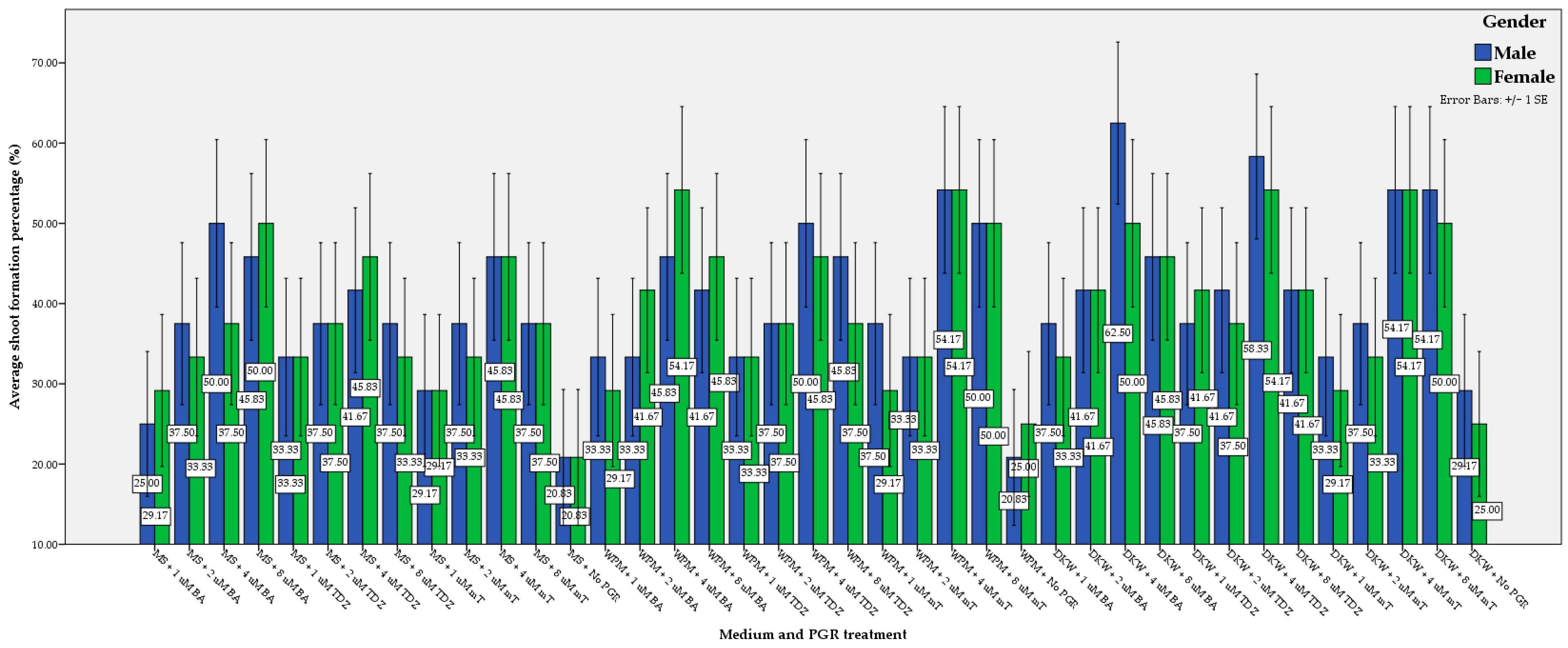
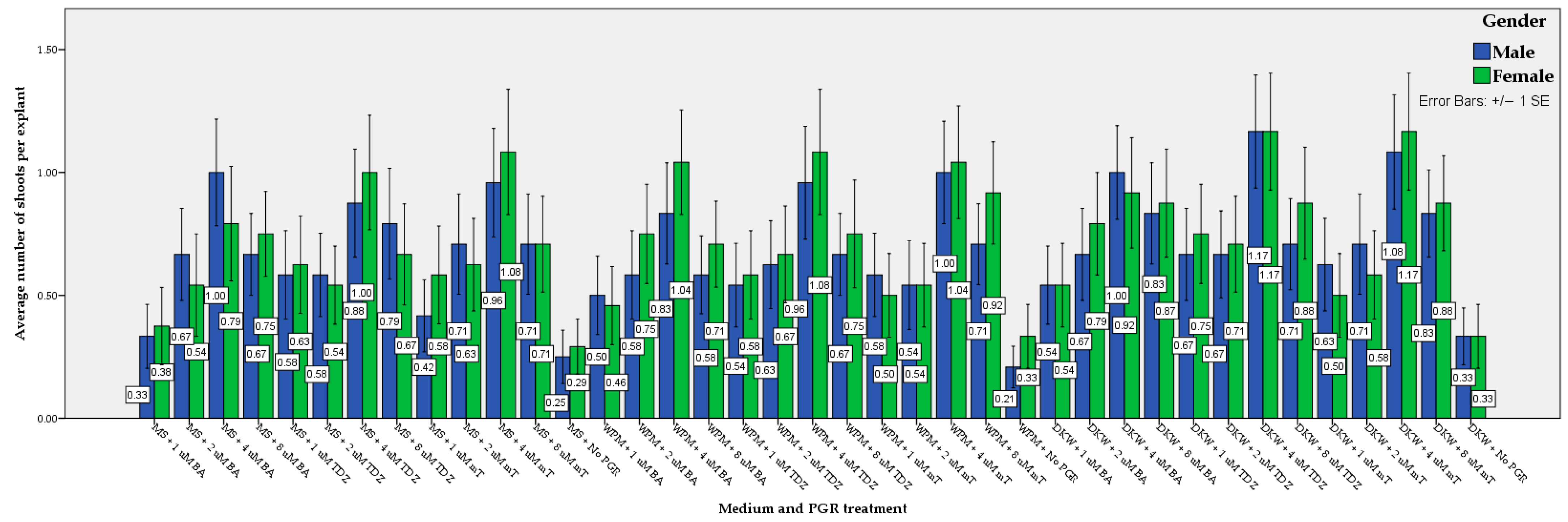
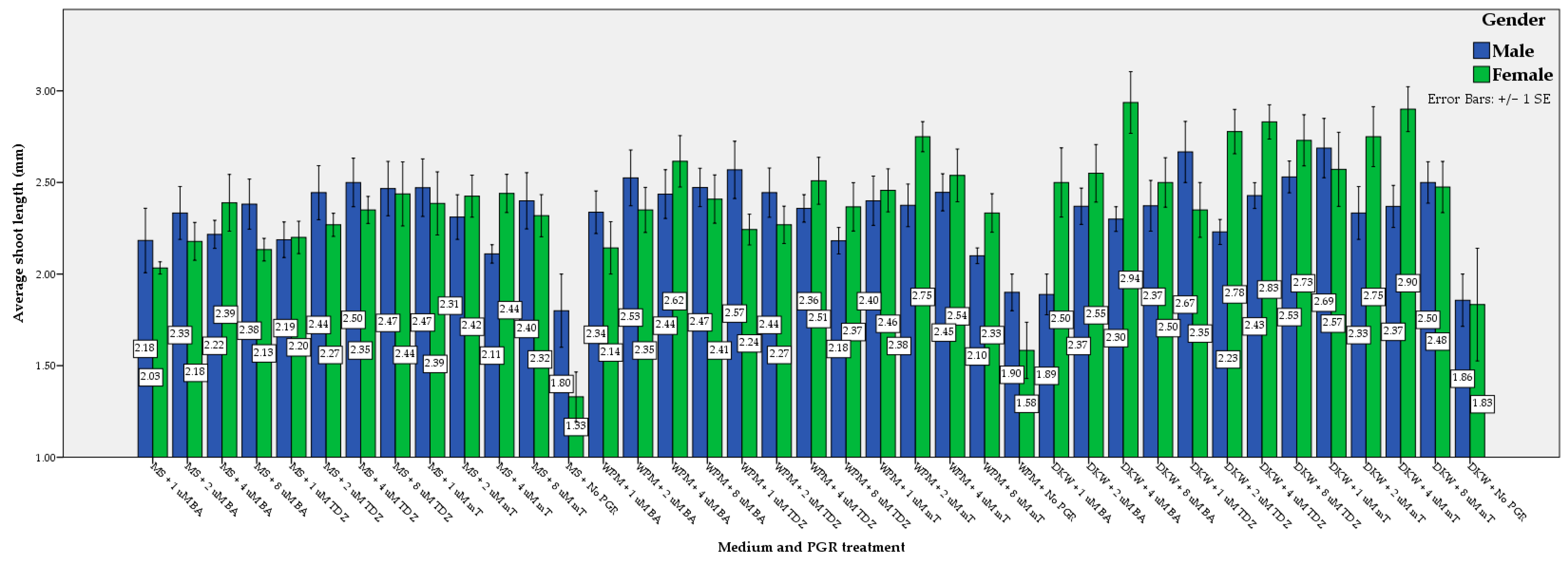
3.1. Shoot Regeneration, Multiplication, and Elongation
3.2. In Vitro Explant Rooting
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Vidakovic, M. Conifers. Morphology and Variation.; Grafičko Zavod Hrvatske: Zagreb, Yugoslavia, 1991; ISBN 0-85198-807-5. [Google Scholar]
- Adams, R.P. Junipers of the World: The Genus Juniperus; Trafford Publishing: Bloomington, IN, USA, 2014; ISBN 1-4907-2325-0. [Google Scholar]
- Sobierajska, K.; Boratyńska, K.; Jasińska, A.; Dering, M.; Ok, T.; Douaihy, B.; Bou Dagher-Kharrat, M.; Romo, Á.; Boratyński, A. Effect of the Aegean Sea Barrier between Europe and Asia on Differentiation in Juniperus drupacea (Cupressaceae). Bot. J. Linn. Soc. 2016, 180, 365–385. [Google Scholar] [CrossRef] [Green Version]
- Coode, M.; Cullen, J. Juniperus L. Flora of Turkey and the East Aegean Islands; Edinburgh University Press: Edinburgh, UK, 1965; Volume 1, pp. 78–84. [Google Scholar]
- Axelrod, D.I. Evolution and Biogeography of Madrean-Tethyan Sclerophyll Vegetation. Ann. Mo. Bot. Gard. 1975, 62, 280–334. [Google Scholar] [CrossRef]
- Kvaček, Z. A New Juniper from the Palaeogene of Central Europe. Feddes Repert. Z. Bot. Taxon. Geobot. 2002, 113, 492–502. [Google Scholar] [CrossRef]
- Palamarev, E. Paleobotanical Evidences of the Tertiary History and Origin of the Mediterranean Sclerophyll Dendroflora. Plant Syst. Evol. 1989, 162, 93–107. [Google Scholar] [CrossRef]
- Boratyński, A.; Browicz, K.; Zieliński, J. Chorology of Trees and Shrubs in Greece; Institute of Dendrology, Polish Academy of Sciences: Kornik, Poland, 1992; ISBN 83-85599-04-5. [Google Scholar]
- Constantinidis, T.A.; Kalpoutzakis, E. Plant Guide to Mount Parnon and Moustos Wetland Protected Area: Endemic, Rare and Threatened Species; Management Body Mount Parnon: Moustos Wetland, Greece, 2015; ISBN 960-99424-4-X. [Google Scholar]
- Tan, K.; Iatrou, G.; Johsen, B. Endemic Plants of Greece: The Peloponnese; Gads Forlag: Copenhagen, Denmark, 2001; Volume 1. [Google Scholar]
- Bergmeier, E. Plant Communities and Habitat Differentiation in the Mediterranean Coniferous Woodlands of Mt. Parnon (Greece). Folia Geobot. 2002, 37, 309–331. [Google Scholar] [CrossRef]
- Dimopoulos, P.; Raus, T.; Bergmeier, E.; Constantinidis, T.; Iatrou, G.; Kokkini, S.; Strid, A.; Tzanoudakis, D. Vascular Plants of Greece: An Annotated Checklist; Botanic Garden and Botanical Museum Berlin-Dahlem: Berlin, Germany, 2013. [Google Scholar]
- Douaihy, B.; Tarraf, P.; Stephan, J. Juniperus drupacea Labill. Stands in Jabal Moussa Biosphere Reserve, a Pilot Study for Management Guidelines. Plant Sociol. 2017, 54, 39–45. [Google Scholar]
- Hazubska-Przybył, T. Propagation of Juniper Species by Plant Tissue Culture: A Mini-Review. Forests 2019, 10, 1028. [Google Scholar] [CrossRef] [Green Version]
- Farjon, A. A Handbook of the World’s Conifers; Brill: Leiden, The Netherlands, 2010; Volume 1. [Google Scholar]
- Directive, H. Council Directive 92/43/EEC of 21 May 1992 on the Conservation of Natural Habitats and of Wild Fauna and Flora. Off. J. Eur. Union 1992, 206, 7–50. [Google Scholar]
- Gardner, M. Juniperus drupacea. The IUCN Red List of Threatened Species 2013: E. T30311A2792553. 2013. Available online: https://doi.org/10.2305/IUCN.UK.2013-1.RLTS.T30311A2792553.en (accessed on 28 October 2021).
- Rivers, M.; Beech, E.; Bazos, I.; Bogunić, F.; Buira, A.; Caković, D.; Carapeto, A.; Carta, A.; Cornier, B.; Fenu, G. European Red List of Trees; International Union for Conservation of Nature and Natural Resources (IUCN): Fontainebleau, France, 2019; ISBN 2-8317-1986-0. [Google Scholar]
- Gardner, M. Juniperus drupacea. The IUCN Red List of Threatened Species 2017: E.T30311A83932989. 2017. Available online: https://doi.org/10.2305/IUCN.UK.2013-1.RLTS.T30311A2792553.en (accessed on 1 November 2022).
- Greek Biotope & Wetland Centre. Available online: https://www.ekby.gr/ekby/en/EKBY_PA_en_.html (accessed on 1 August 2022).
- Yucedag, C.; Cetin, M.; Ozel, H.B.; Abo Aisha, A.E.S.; Alrabiti, O.B.M.; AL JAMA, A.M.O. The Impacts of Altitude and Seed Pretreatments on Seedling Emergence of Syrian Juniper (Juniperus drupacea (Labill.) Ant. et Kotschy). Ecol. Process. 2021, 10, 1–6. [Google Scholar] [CrossRef]
- Gaussen, H. Les Gymnospermes Actuelles et Fossiles, Fascicle 10, Les Cuppressacées; Centre National de la Recherche Scientifique, Faculté des Sciences de Toulouse: Toulouse, France, 1968. [Google Scholar]
- Maerki, D.; Frankis, M. Juniperus drupacea in the Peloponnese (Greece). Trip Report and Range Map, with Notes on Phenology, Phylogeny, Palaeontology, History, Types and Use. Bull. CCP 2015, 3, 3–31. [Google Scholar]
- Christensen, K.; Juniperus, L. Flora Hellenica; Strid, A., Tan, K., Eds.; Koeltz Scientific Books: Κönigstein, Germany, 1997; Volume 1, pp. 10–14. [Google Scholar]
- Gültekin, H.C.; Gültekin, Ü.G.; Divrik, A. Andız (Arceuthos drupacea (Labill.) Ant. et. Kotschy.) Tohumlarının Çimlenmesi, Diğer Tohum ve Fidan Özelliklerine Ilişkin Bazı Tespit ve Öneriler. J. Artvin Coruh Univ. For. Fac. 2004, 5, 48–54. [Google Scholar]
- Gómez, M.P.; Segura, J. Morphogenesis in Leaf and Single-Cell Cultures of Mature Juniperus oxycedrus. Tree Physiol. 1996, 16, 681–686. [Google Scholar] [CrossRef] [PubMed]
- Loureiro, J.; Capelo, A.; Brito, G.; Rodriguez, E.; Silva, S.; Pinto, G.; Santos, C. Micropropagation of Juniperus phoenicea from Adult Plant Explants and Analysis of Ploidy Stability Using Flow Cytometry. Biol. Plant. 2007, 51, 7–14. [Google Scholar] [CrossRef]
- Khater, N.; Benbouza, H. Preservation of Juniperus thurifera L.: A Rare Endangered Species in Algeria through in Vitro Regeneration. J. For. Res. 2019, 30, 77–86. [Google Scholar] [CrossRef]
- Kentelky, E. The Analysis of Rooting and Growth Peculiarities of Juniperus Species Propagated by Cuttings. Bull. UASVM Hortic 2011, 68, 380–385. [Google Scholar]
- Hartmann, H.T.; Kester, D.E.; Davies, F.T.; Geneve, R.L. Plant Propagation: Principles and Practices.; Prentice-Hall Inc.: Hoboken, NJ, USA, 1997; ISBN 0-13-261488-X. [Google Scholar]
- Berhe, D.; Negash, L. Asexual Propagation of Juniperus Procera from Ethiopia: A Contribution to the Conservation of African Pencil Cedar. For. Ecol. Manag. 1998, 112, 179–190. [Google Scholar] [CrossRef]
- Ragonezi, C.; Klimaszewska, K.; Castro, M.R.; Lima, M.; de Oliveira, P.; Zavattieri, M.A. Adventitious Rooting of Conifers: Influence of Physical and Chemical Factors. Trees 2010, 24, 975–992. [Google Scholar] [CrossRef] [Green Version]
- Gómez, M.P.; Segura, J. Axillary Shoot Proliferation in Cultures of Explants from Mature Juniperus oxycedrus Trees. Tree Physiol. 1995, 15, 625–628. [Google Scholar] [CrossRef]
- Harry, I.S.; Pulido, C.M.; Thorpe, T.A. Plantlet Regeneration from Mature Embryos of Juniperus cedrus. Plant Cell Tissue Organ Cult. 1995, 41, 75–78. [Google Scholar] [CrossRef]
- Castro, M.R.; Belo, A.F.; Afonso, A.; Zavattieri, M.A. Micropropagation of Juniperus navicularis, an Endemic and Rare Species from Portugal SW Coast. Plant Growth Regul. 2011, 65, 223–230. [Google Scholar] [CrossRef] [Green Version]
- Gómez, M.P.; Segura, J. Factors Controlling Adventitious Bud Induction and Plant Regeneration in Mature Juniperus oxycedrus Leaves Cultured In Vitro. Plant 1994, 30, 210–218. [Google Scholar] [CrossRef]
- Al-Ramamneh, E.A.; Dura, S.; Daradkeh, N. Propagation Physiology of Juniperus phoenicea L. from Jordan Using Seeds and in Vitro Culture Techniques: Baseline Information for a Conservation Perspective. Afr. J. Biotechnol. 2012, 11, 7684–7692. [Google Scholar] [CrossRef]
- Zaidi, M.A.; Khan, S.; Jahan, N.; Yousafzai, A.; Mansoor, A. Micropropagation and Conservation of Three Juniperus Species (Cupressaceae). Pak. J. Bot. 2012, 44, 301–304. [Google Scholar]
- Kashani, S.A.; Asrar, M.; Leghari, S.K. In Vitro Callus Induction and Shoot Formation of Juniperus excelsa of Ziarat, Balochistan, Pakistan. Fuuast J. Biol. 2018, 8, 203–208. [Google Scholar]
- Momeni, M.; Ganji-Moghadam, E.; Kazemzadeh-Beneh, H.; Asgharzadeh, A. Direct Organogenesis from Shoot Tip Explants of Juniperus Polycarpos L.: Optimizing Basal Media and Plant Growth Regulators on Proliferation and Root Formation. Plant Cell Biotechnol. Mol. Biol 2018, 19, 40–50. [Google Scholar]
- Fett-Neto, A.G.; Fett, J.P.; Goulart, L.W.V.; Pasquali, G.; Termignoni, R.R.; Ferreira, A.G. Distinct Effects of Auxin and Light on Adventitious Root Development in Eucalyptus Saligna and Eucalyptus Globulus. Tree Physiol. 2001, 21, 457–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenwood, M.S.; Cui, X.; Xu, F. Response to Auxin Changes during Maturation-related Loss of Adventitious Rooting Competence in Loblolly Pine (Pinus taeda) Stem Cuttings. Physiol. Plant. 2001, 111, 373–380. [Google Scholar] [CrossRef]
- Bielenin, M. Rooting and Gas Exchange of Conifer Cuttings Treated with Indolebutyric Acid. J. Fruit Ornam. Plant Res. 2003, 11, 99–106. [Google Scholar]
- Henrique, A.; Campinhos, E.N.; Ono, E.O.; de Pinho, S.Z. Effect of Plant Growth Regulators in the Rooting of Pinus Cuttings. Braz. Arch. Biol. Technol. 2006, 49, 189–196. [Google Scholar] [CrossRef]
- Harry, I.S.; Thorpe, T.A. In Vitro Culture of Forest Trees. In Plant Cell and Tissue Culture; Vasil, I.K., Thorpe, T.A., Eds.; Springer Netherlands: Dordrecht, The Netherlands, 1994; pp. 539–560. ISBN 978-94-017-2681-8. [Google Scholar]
- Bertsouklis, K.; Paraskevopoulou, A.; Zarkadoula, N. In Vitro Propagation of Juniperus phoenicea L. Acta Hortic. 2019, 1298, 331–334. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Lloyd, G.; McCown, B. Commercially-Feasible Micropropagation of Mountain Laurel, Kalmia Latifolia, by Use of Shoot-Tip Culture. Comb. Proc.-Int. Plant Propagator’s Soc. 1980, 30, 421–427. [Google Scholar]
- Driver, J.A.; Kuniyuki, A.H. In Vitro Propagation of Paradox Walnut Rootstock. HortScience 1984, 19, 507–509. [Google Scholar] [CrossRef]
- Salih, A.M.; Al-Qurainy, F.; Khan, S.; Tarroum, M.; Nadeem, M.; Shaikhaldein, H.O.; Alabdallah, N.M.; Alansi, S.; Alshameri, A. Mass Propagation of Juniperus procera Hoechst. Ex Endl. From Seedling and Screening of Bioactive Compounds in Shoot and Callus Extract. BMC Plant Biol. 2021, 21, 192. [Google Scholar] [CrossRef]
- Ge, X.; Chu, Z.; Lin, Y.; Wang, S. A Tissue Culture System for Different Germplasms of Indica Rice. Plant Cell Rep. 2006, 25, 392–402. [Google Scholar] [CrossRef]
- Thorpe, T.A.; Harry, I.S.; Kumar, P.P. Application of Micropropagation to Forestry. In Micropropagation: Technology and Application; Debergh, P.C., Zimmerman, R.H., Eds.; Springer Netherlands: Dordrecht, The Netherlands, 1991; pp. 311–336. ISBN 978-94-009-2075-0. [Google Scholar]
- Al-Ramamneh, E.A.-D.; Daradkeh, N.; Rababah, T.; Pacurar, D.; Al-Qudah, M. Effects of Explant, Media and Growth Regulators on in Vitro regeneration and Antioxidant Activity of Juniperus phoenicea. Aust. J. Crop Sci. 2017, 11, 828–837. [Google Scholar] [CrossRef]
- Oliveira, P.; Barriga, J.; Cavaleiro, C.; Peixe, A.; Potes, A.Z. Sustained in Vitro Root Development Obtained in Pinus pinea L. Inoculated with Ectomycorrhizal Fungi. For. Int. J. For. Res. 2003, 76, 579–587. [Google Scholar] [CrossRef]
- Renau-Morata, B.; Ollero, J.; Arrillaga, I.; Segura, J. Factors Influencing Axillary Shoot Proliferation and Adventitious Budding in Cedar. Tree Physiol. 2005, 25, 477–486. [Google Scholar] [CrossRef] [Green Version]
- Negussie, A. In Vitro Induction of Multiple Buds in Tissue Culture of Juniperus excelsa. For. Ecol. Manag. 1997, 98, 115–123. [Google Scholar] [CrossRef]
- Brito, G.M.O. Micropropagaçao de Duas Espécies Autóctones Da Ilha de Porto Santo (Olea europaea L. ssp. Maderensis Lowe e Juniperus phoenica L.) e Estudo Da Resposta de Rebentos in Vitro a Stress Osmótico. Master’s Thesis, Universidade de Aveiro, Aveiro, Portugal, 2000. [Google Scholar]
- Smith, N.G.; Wareing, P.F. The Rooting of Actively Growing and Dormant Leafy Cuttings in Relation to Endogenous Hormone Levels and Photoperiod. New Phytol. 1972, 71, 483–500. [Google Scholar] [CrossRef]
- Hyndman, S.F.; Hasegawa, P.M.; Bressan, R.A. Of Mineral Salts. HortScience 1982, 17, 82–83. [Google Scholar] [CrossRef]
- Sha Valli Khan, P.S.; Hausman, J.F.; Rao, K.R. Effect of Agar, MS Medium Strength, Sucrose and Polyamines on in Vitro Rooting of Syzygium Alternifolium. Biol. Plant. 1999, 42, 333–340. [Google Scholar] [CrossRef]
- Fadel, D. Effect of Different Strength of Medium on Organogenesis, Phenolic Accumulation and Antioxidant Activity of Spearmint (Mentha spicata L.). TOHORTJ 2010, 3, 31–35. [Google Scholar] [CrossRef]
- Ioannidis, K.; Dadiotis, E.; Mitsis, V.; Melliou, E.; Magiatis, P. Biotechnological Approaches on Two High CBD and CBG Cannabis sativa L. (Cannabaceae) Varieties: In Vitro Regeneration and Phytochemical Consistency Evaluation of Micropropagated Plants Using Quantitative 1H-NMR. Molecules 2020, 25, 5928. [Google Scholar] [CrossRef]
- Ordas, R.; Rodríguez, A.; Rodríguez, R.; Sanchez Tames, R. Desarrollo de Tecnicas de Cultivo «in Vitro» Para La Micropropagation de Variedades de Manzana Sidrera. In Anales de edafologia y agrobiologia; Consejo Superior de Investigaciones Científicas. Instituto Nacional de Edafología y Agrobiología: Madrid, Spain, 1984; Volume 43, pp. 905–917. [Google Scholar]
- Trewavas, A.J.; Cleland, R.E. Is Plant Development Regulated by Changes in the Concentration of Growth Substances or by Changes in the Sensitivity to Growth Substances? Trends Biochem. Sci. 1983, 8, 354–357. [Google Scholar] [CrossRef]
- Klimaszewska, K.; Trontin, J.-F.; Becwar, M.R.; Devillard, C.; Park, Y.-S.; Lelu-Walter, M.-A. Recent Progress in Somatic Embryogenesis of Four Pinus Spp. Tree For. Sci. Biotechnol. 2007, 1, 11–25. [Google Scholar]
- Helmersson, A.; von Arnold, S. Embryogenic Cell Lines of Juniperus communis; Easy Establishment and Embryo Maturation, Limited Germination. Plant Cell Tissue Organ Cult. PCTOC 2009, 96, 211–217. [Google Scholar] [CrossRef]
- Von Arnold, S.; Egertsdotter, U.; Ekberg, I.; Gupta, P.; Mo, H.; Nörgaard, J. Somatic Embryogenesis in Norway Spruce (Picea abies). Somat. Embryog. Woody Plants 1995, 3, 17–36. [Google Scholar]
- Von Arnold, S.; Clapham, D. Spruce Embryogenesis. In Plant Embryogenesis; Springer: Berlin/Heidelberg, Germany, 2008; pp. 31–47. [Google Scholar]
- Salaj, T.; Klubicová, K.; Matusova, R.; Salaj, J. Somatic Embryogenesis in Selected Conifer Trees Pinus nigra Arn. and Abies Hybrids. Front. Plant Sci. 2019, 10, 13. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment Concentration | |||
|---|---|---|---|
| PGR | MS Medium | WPM Medium | DKW Medium |
| BA or TDZ or m-T | Control | Control | Control |
| 1.0 μM | 1.0 μM | 1.0 μM | |
| 2.0 μM | 2.0 μM | 2.0 μM | |
| 4.0 μM | 4.0 μM | 4.0 μM | |
| 8.0 μM | 8.0 μM | 8.0 μM | |
| Treatment Concentration | |||||
|---|---|---|---|---|---|
| PGR | MS WPM DKW Media | PGR | MS WPM DKW Media | PGR | MS WPM DKW Media |
| IBA | Control | NAA | Control | IAA | Control |
| 1.0 μM | |||||
| 2.0 μM | 1.0 μM | 0.5 μM | |||
| 4.0 μM | 2.0 μM | 1.0 μM | |||
| 8.0 μM | 4.0 μM | 2.0 μM | |||
| 16.0 μM | 8.0 μM | 4.0 μM | |||
| 32.0 μM | |||||
| Average Percentage of Blastogenesis (%) | Average Number of Shoots per Explant | Average Shoot Length per Explant (mm) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Explant Gender | Female | Male | Overall | Female | Male | Overall | Female | Male | Overall | |
| N | 312 | 312 | 624 | 312 | 312 | 624 | 312 | 312 | 624 | |
| Nutrient Medium | DKW | 41.35 a | 44.23 a | 42.79 a | 0.78 a | 0.76 a | 0.77 a | 1.08 a | 1.04 a | 1.06 a |
| WPM | 39.74 ab | 39.74 ab | 39.74 ab | 0.72 ab | 0.66 ab | 0.68 ab | 0.95 ab | 0.95 ab | 0.95 ab | |
| MS | 35.90 b | 36.86 b | 36.38 b | 0.66 b | 0.64 b | 0.66 b | 0.80 b | 0.86 b | 0.83 b | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ioannidis, K.; Tomprou, I.; Panayiotopoulou, D.; Boutsios, S.; Daskalakou, E.N. Potential and Constraints on In Vitro Micropropagation of Juniperus drupacea Labill. Forests 2023, 14, 142. https://doi.org/10.3390/f14010142
Ioannidis K, Tomprou I, Panayiotopoulou D, Boutsios S, Daskalakou EN. Potential and Constraints on In Vitro Micropropagation of Juniperus drupacea Labill. Forests. 2023; 14(1):142. https://doi.org/10.3390/f14010142
Chicago/Turabian StyleIoannidis, Kostas, Ioanna Tomprou, Danae Panayiotopoulou, Stefanos Boutsios, and Evangelia N. Daskalakou. 2023. "Potential and Constraints on In Vitro Micropropagation of Juniperus drupacea Labill." Forests 14, no. 1: 142. https://doi.org/10.3390/f14010142
APA StyleIoannidis, K., Tomprou, I., Panayiotopoulou, D., Boutsios, S., & Daskalakou, E. N. (2023). Potential and Constraints on In Vitro Micropropagation of Juniperus drupacea Labill. Forests, 14(1), 142. https://doi.org/10.3390/f14010142