

Evaluation of the Community Land Model-Simulated Specific Leaf Area with Observations over China: Impacts on Modeled Gross Primary Productivity


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
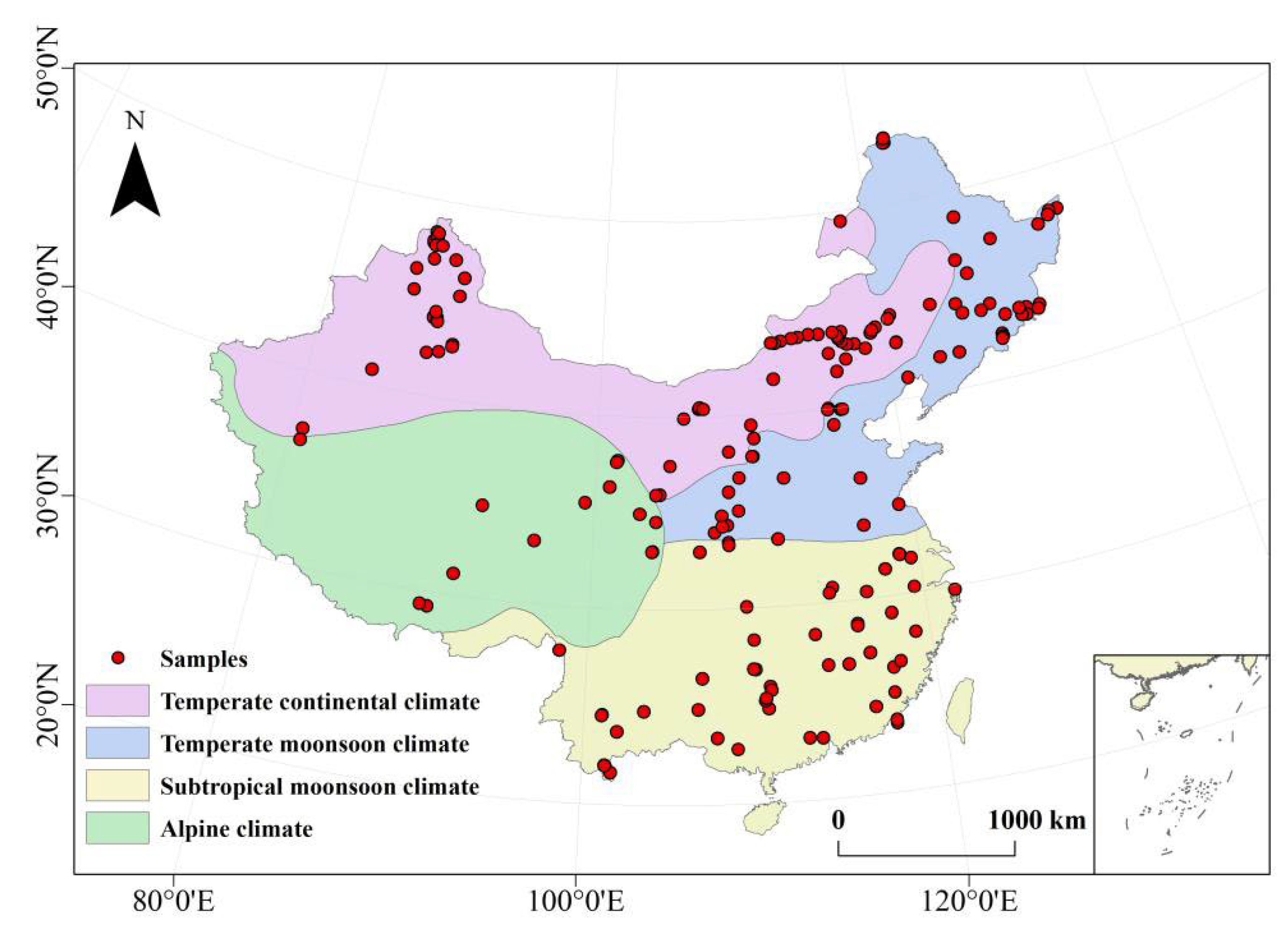
2.1. Data
2.2. Model
2.3. Analysis of the Impact of SLA Variation on Gross Primary Productivity
3. Results
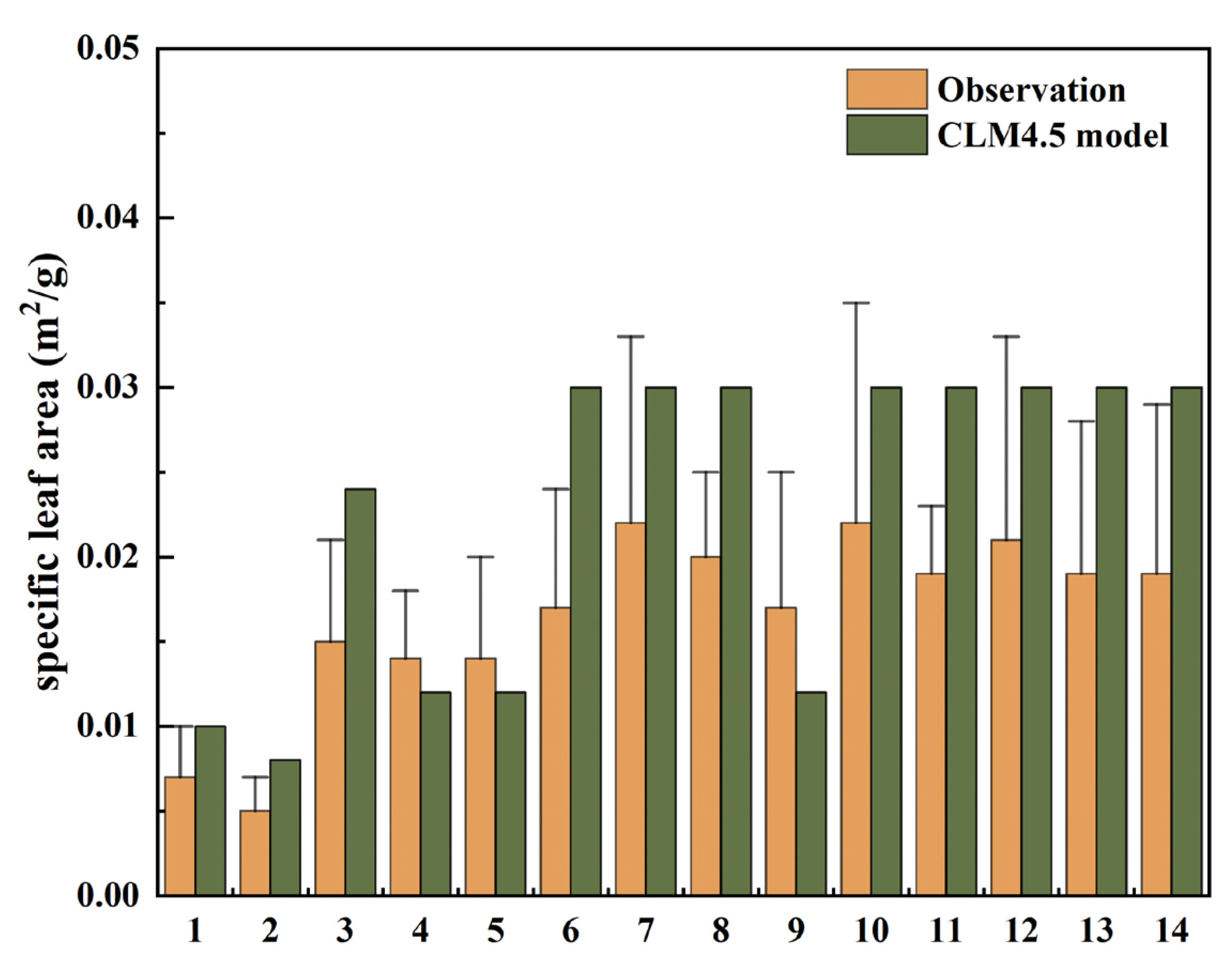
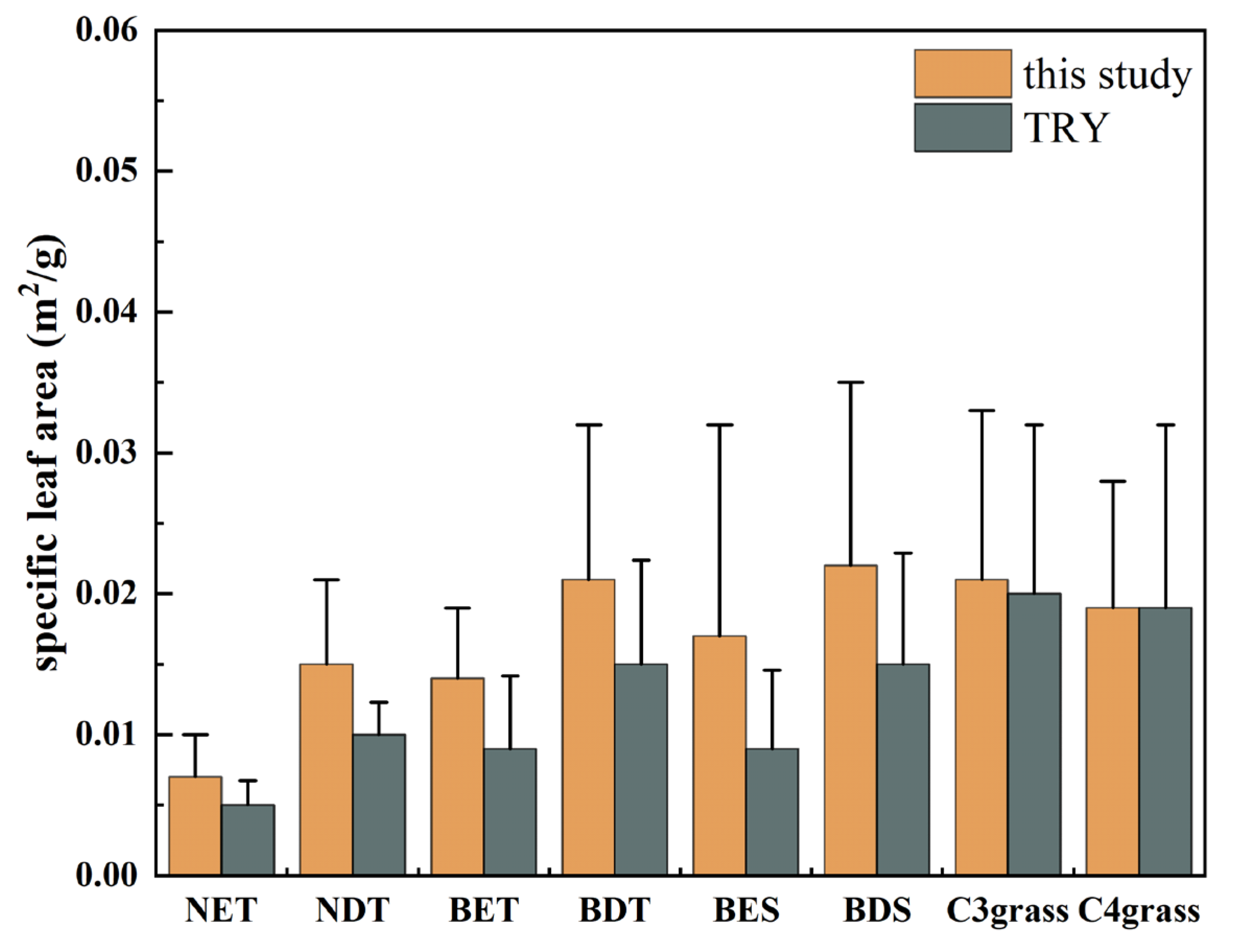
3.1. Comparison of SLA between the CLM Model and Observations over China
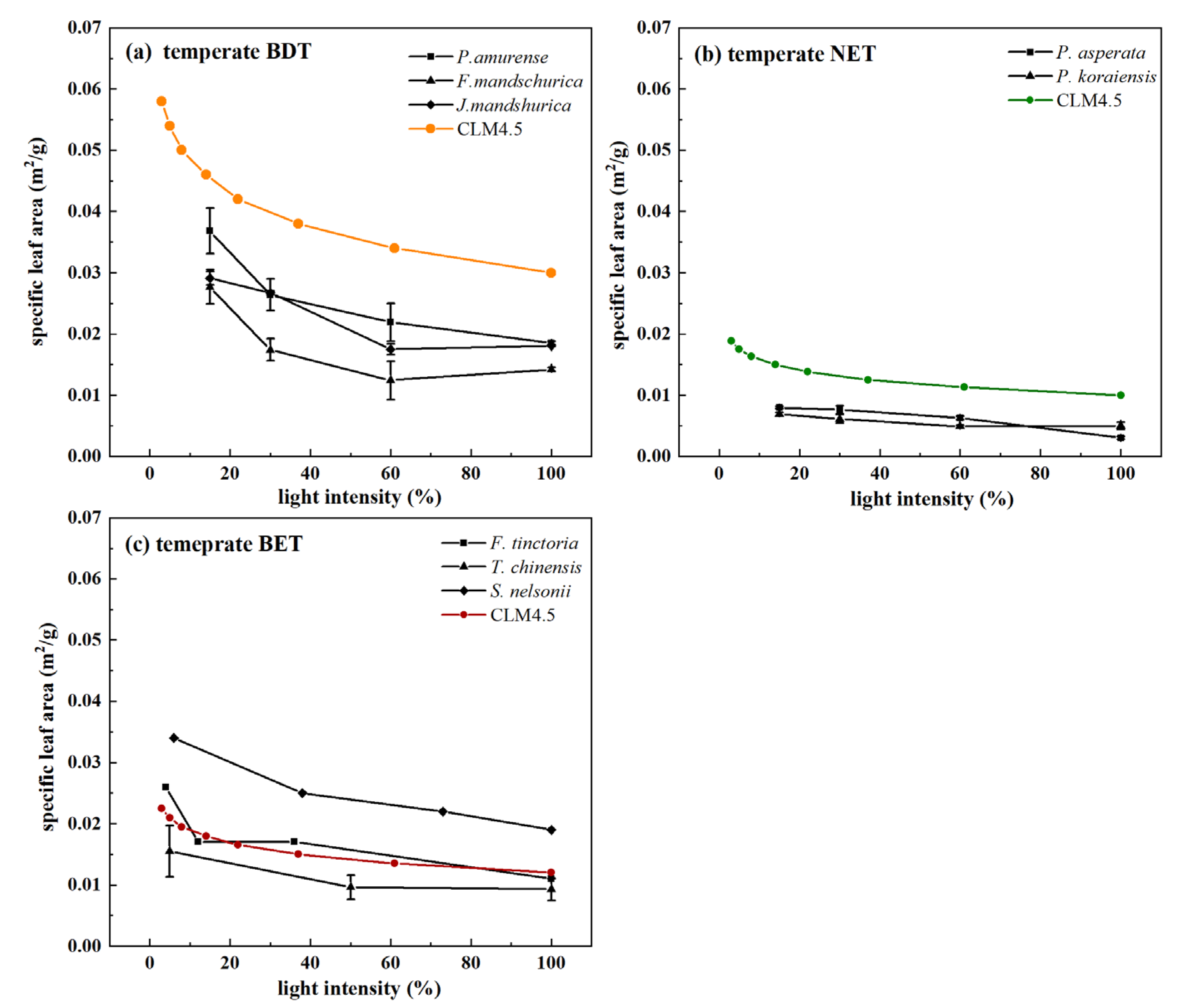
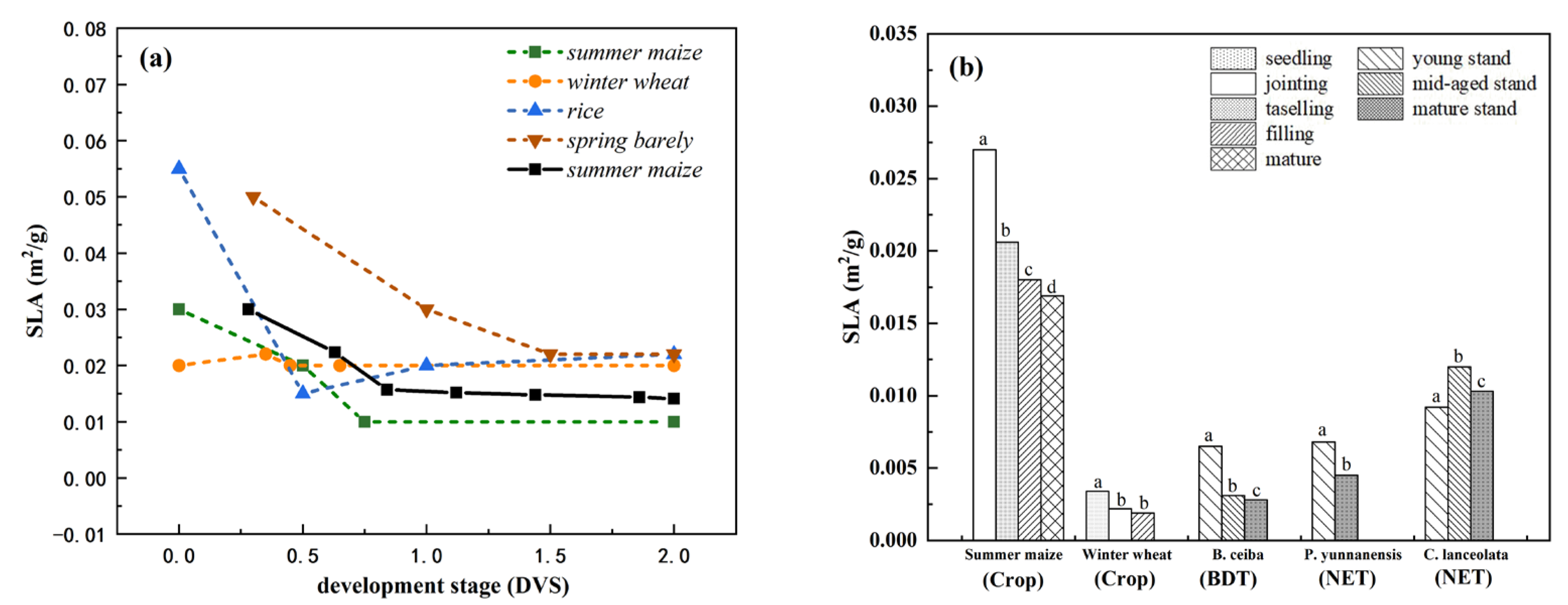
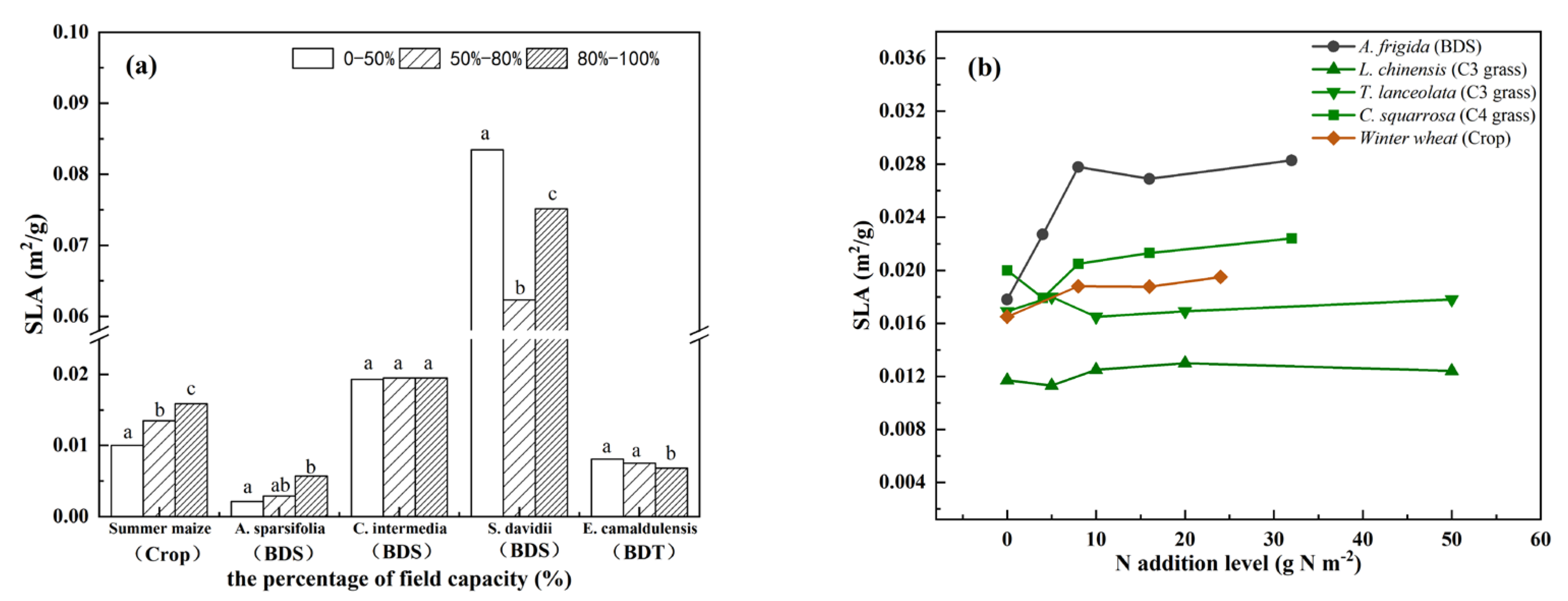
3.2. Interspecific Variation in Observed Plant SLA within Plant Functional Types
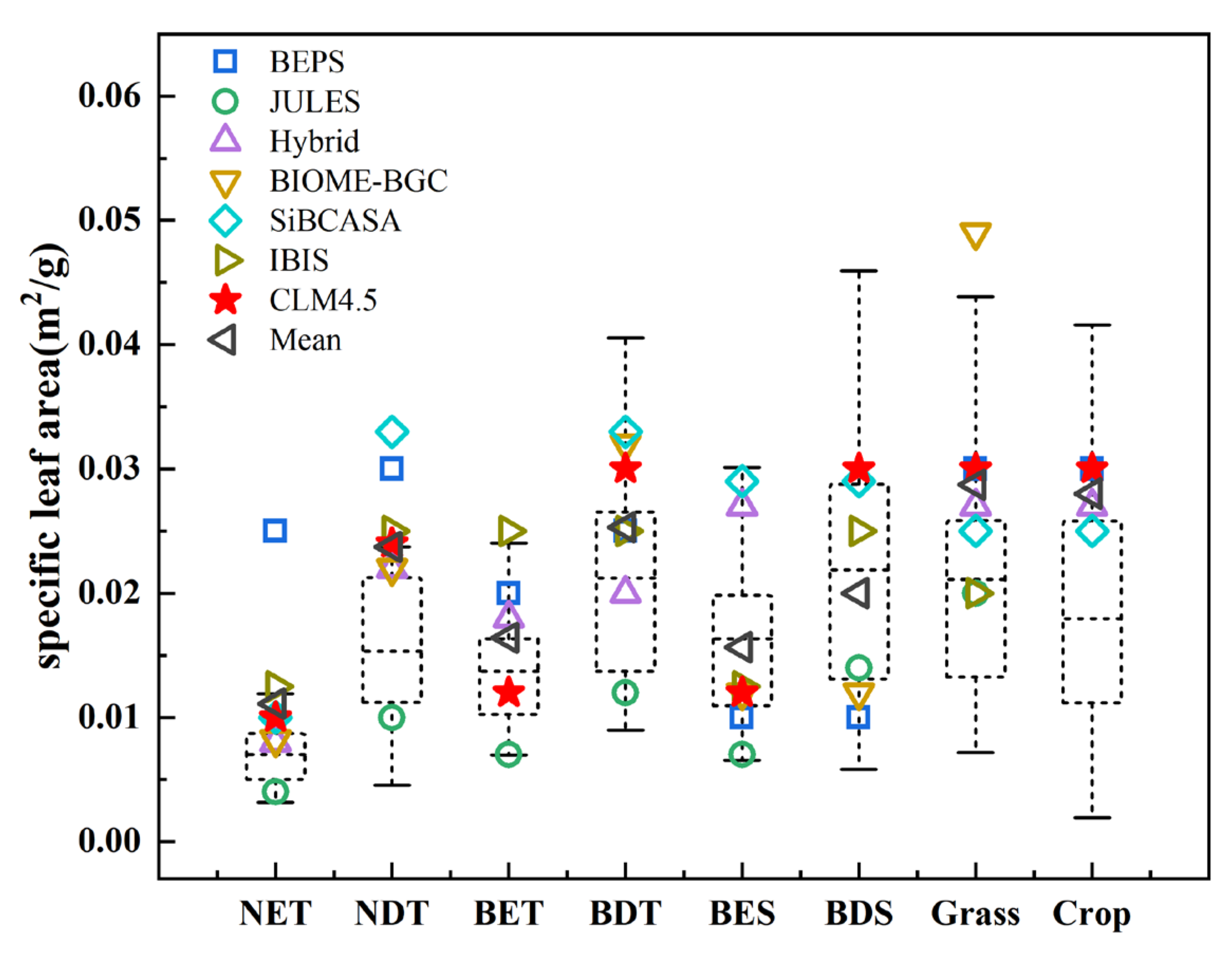
3.3. Variation in the Parameter Values of SLA among Different Terrestrial Biosphere Models
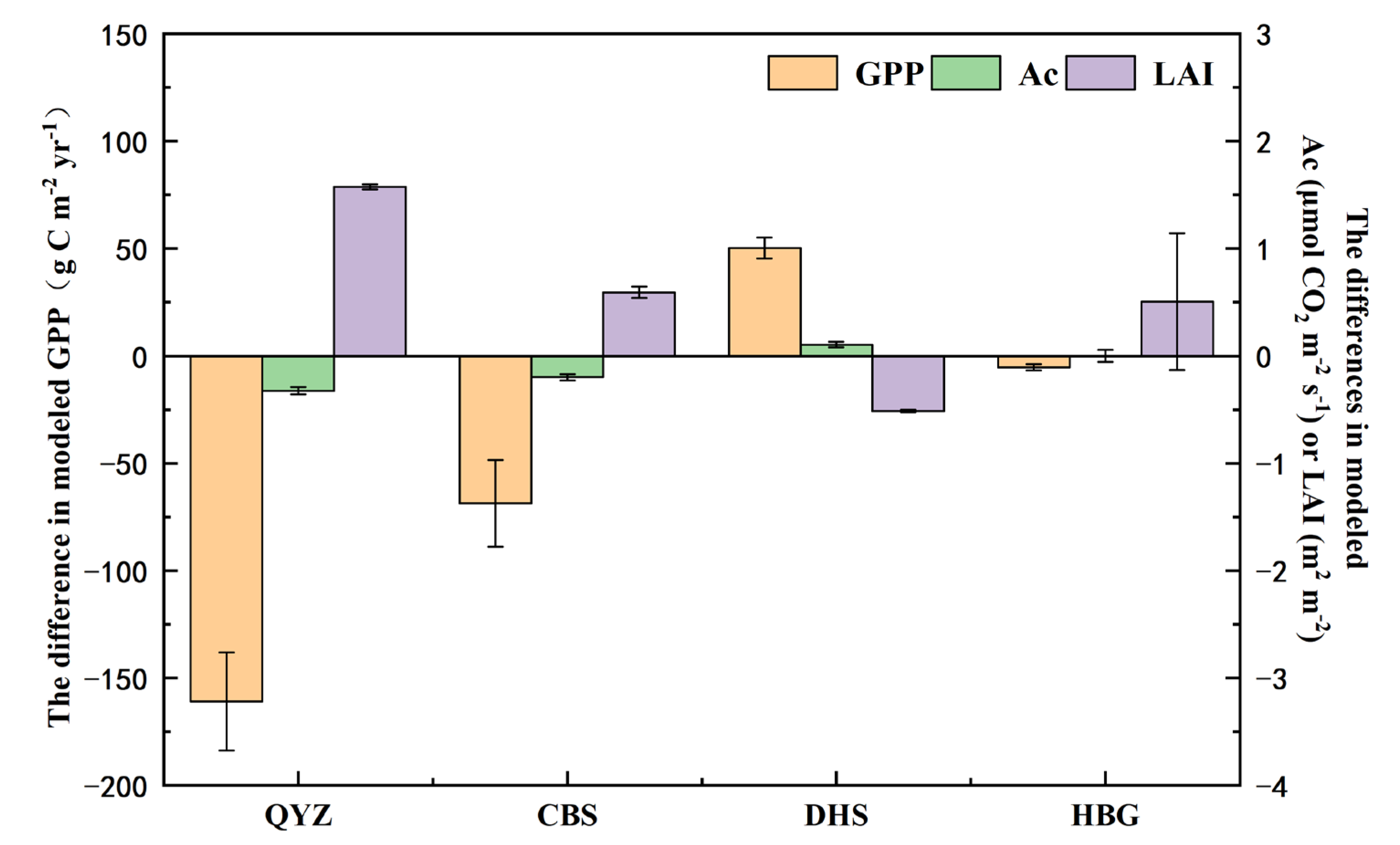
3.4. Impacts of Variation in SLA on Modeled Gross Primary Productivity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Violle, C.; Navas, M.L.; Vile, D.; Kazakou, E.; Fortunel, C.; Hummel, I.; Garnier, E. Let the concept of trait be functional! Oikos 2007, 116, 882–892. [Google Scholar] [CrossRef]
- Lavorel, S.; Garnier, E. Predicting changes in community composition and ecosystem functioning from plant traits: Revisiting the Holy Grail. Funct. Ecol. 2002, 16, 545–556. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Cornelissen, J.H.C.; Falster, D.S.; Groom, P.K.; Hikosaka, K.; Lee, W.; Lusk, C.H.; Niinemets, U.; Oleksyn, J.; et al. Modulation of leaf economic traits and trait relationships by climate. Glob. Ecol. Biogeogr. 2005, 14, 411–421. [Google Scholar] [CrossRef]
- Osnas, J.L.D.; Lichstein, J.W.; Reich, P.B.; Pacala, S.W. Global Leaf Trait Relationships: Mass, Area, and the Leaf Economics Spectrum. Science 2013, 340, 741–744. [Google Scholar] [CrossRef] [PubMed]
- Reich, P.B. The world-wide ‘fast-slow’ plant economics spectrum: A traits manifesto. J. Ecol. 2014, 102, 275–301. [Google Scholar] [CrossRef]
- Westoby, M.; Wright, I.J. Land-plant ecology on the basis of functional traits. Trends Ecol. Evol. 2006, 21, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Finegan, B.; Peña-Claros, M.; Oliveira, A.; Ascarrunz, N.; Bret-Harte, M.S.; Carreño-Rocabado, G.; Casanoves, F.; Díaz, S.; Velepucha, P.E.; Fernandez, F.; et al. Does functional trait diversity predict above-ground biomass and productivity of tropical forests? Testing three alternative hypotheses. J. Ecol. 2015, 103, 191–201. [Google Scholar] [CrossRef] [Green Version]
- Reich, P.B.; Ellsworth, D.S.; Walters, M.B.; Vose, J.M.; Gresham, C.; Volin, J.C.; Bowman, W.D. Generality of leaf trait relationship: A test across six biomes. Ecology 1999, 80, 1955–1969. [Google Scholar] [CrossRef]
- Kattge, J.; DÍAz, S.; Lavorel, S.; Prentice, I.C.; Leadley, P.; BÖNisch, G.; Garnier, E.; Westoby, M.; Reich, P.B.; Wright, I.J.; et al. TRY- a global database of plant traits. Glob. Change Biol. 2011, 17, 2905–2935. [Google Scholar] [CrossRef]
- Wang, R.L.; Yu, G.R.; He, N.P.; Wang, Q.F.; Zhao, N.; Xu, Z.W. Latitudinal variation of leaf morphological traits from species to communities along a forest transect in eastern China. J. Geogr. Sci. 2016, 26, 15–26. [Google Scholar] [CrossRef]
- Wang, H.; Prentice, I.C.; Ni, J. Data-based modelling and environmental sensitivity of vegetation in China. Biogeosciences 2013, 10, 5817–5830. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.G.; Zhao, M.; Zhang, H.X.; Ren, T.T.; Liu, C.C.; He, N.P. Divergent response and adaptation of specific leaf area to environmental change at different spatial-temporal scales jointly improve plant survival. Glob. Change Biol. 2022, in press. [Google Scholar] [CrossRef] [PubMed]
- Reich, P.B.; Ellsworth, D.S.; Walters, M.B. Leaf structure (specific leaf area) modulates photosynthesis–nitrogen relations: Evidence from within and across species and functional groups. Funct. Ecol. 1998, 12, 948–958. [Google Scholar] [CrossRef]
- Lavorel, S.; Díaz, S.; Cornelissen, J.H.C.; Garnier, E.; Harrison, S.P.; Mclntyre, S.; Pausas, J.G.; Pérez-Harguindeguy, N.; Roumet, C.; Urcelay, C. Plant functional types: Are we getting any closer to the holy grail. In Terrestrial Ecosystems in a Changing World; Canadell, J.G., Pataki, D.E., Pitelka, L.F., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 149–164. [Google Scholar]
- Yang, Y.Z.; Wang, H.; Zhu, Q.; Wen, Z.M.; Peng, C.H.; Lin, G.H. Research progresses in improving dynamic global vegetation models (DGVMs) with plant functional traits. Chin. Sci. Bull. 2018, 63, 2599–2611. [Google Scholar] [CrossRef] [Green Version]
- Raulier, F.; Bernier, P.Y.; Ung, C.-H. Canopy photo-synthesis of sugar maple (Acer saccharum): Comparing big-leaf and multilayer extrapolations of leaf-level measure-ments. Tree Physiol. 1999, 19, 407–442. [Google Scholar] [CrossRef] [Green Version]
- Thornton, P.E.; Zimmermann, N.E. An improved canopy integration scheme for a land surface model with prognostic canopy structure. J. Clim. 2007, 20, 3902–3923. [Google Scholar] [CrossRef]
- Harper, A.B.; Cox, P.M.; Friedlingstein, P.; Wiltshire, A.J.; Jones, C.D.; Sitch, S.; Mercado, L.M.; Groenendijk, M.; Robertson, E.; Kattge, J.; et al. Improved representation of plant functional types and physiology in the Joint UK Land Environment Simulator (JULES v4.2) using plant trait information. Geosci. Model Dev. 2016, 9, 2415–2440. [Google Scholar] [CrossRef] [Green Version]
- Wramneby, A.; Smith, B.; Zaehle, S.; Sykes, M.T. Parameter uncertainties in the modeling of vegetation dynamics-effects on tree community structure and ecosystem functioning in European forest biomes. Ecol. Modell. 2008, 216, 277–290. [Google Scholar] [CrossRef]
- Cui, E.Q.; Huang, K.; Arain, M.A.; Fisher, J.B.; Huntzinger, D.N.; Ito, A.; Luo, Y.Q.; Jain, A.K.; Mao, J.F.; Michalak, A.M.; et al. Vegetation Functional Properties Determine Uncertainty of Simulated Ecosystem Productivity: A Traceability Analysis in the East Asian Monsoon Region. Glob. Biogeochem. Cycles 2019, 33, 668–689. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.C.; Liu, D.; Dong, W.J.; Cai, W.W.; Yuan, W.P. Accurate representation of leaf longevity is important for simulating ecosystem carbon cycle. Basic Appl. Ecol. 2016, 17, 396–407. [Google Scholar] [CrossRef]
- Feng, X.; Liu, G.; Chen, J.M.; Chen, M.; Liu, J.; Ju, W.M.; Sun, R.; Zhou, W. Net primary productivity of China’s terrestrial ecosystems from a process model driven by remote sensing. J. Environ. Manag. 2007, 85, 563–573. [Google Scholar] [CrossRef]
- White, M.A.; Thornton, P.E.; Running, S.W.; Nemani, R.R. Parameterization and sensitivity analysis of the BIOME-BGC terrestrial ecosystem model: Net primary production controls. Earth Interact. 2000, 4, 1–85. [Google Scholar] [CrossRef]
- Schaefer, K.; Collatz, G.J.; Tans, P.; Denning, A.S.; Baker, I.; Berry, J.; Prihodko, L. Combined Simple Biosphere/Carnegie-Ames-Stanford Approach terrestrial carbon cycle model. J. Geophys. Res. Biogeosci. 2006, 113, 603. [Google Scholar] [CrossRef]
- Oleson, K.W.; Lawrence, D.M.; Bonan, G.B.; Drewniak, B.; Huang, M.Y.; Koven, C.D.; Levis, S.; Li, F.; Riley, W.J.; Subin, Z.M.; et al. Technical Description of Version 4.5 of the Community Land Model (CLM); Ncar Technical Note NCAR/TN-503 + STR; National Center for Atmospheric Research: Boulder, CO, USA, 2013. [Google Scholar]
- Kucharik, C.J.; Foley, J.A.; Delire, C.; Fisher, V.A.; Coe, M.T.; Lenters, J.D.; Young-Molling, C.; Ramankutty, N.; Norman, J.M.; Gower, S.T. Testing the performance of a dynamic global ecosystem model: Water balance, carbon balance, and vegetation structure. Glob. Biogeochem. Cycles 2000, 14, 795–825. [Google Scholar] [CrossRef]
- Zhang, L.; Mao, J.F.; Shi, X.Y.; Ricciuto, D.; He, H.L.; Thornton, P.; Yu, G.R.; Li, P.; Liu, M.; Ren, X.L.; et al. Evaluation of the Community Land Model simulated carbon and water fluxes against observations over ChinaFLUX sites. Agric. For. Meteorol. 2016, 226, 174–185. [Google Scholar] [CrossRef] [Green Version]
- Fox, A.M.; Hoar, T.J.; Anderson, J.L.; Arellano, A.F.; Smith, W.K.; Litvak, M.E.; MacBean, N.; Schimel, D.S.; Moore, D.J.P. Evaluation of a Data Assimilation System for Land Surface Models Using CLM4.5. J. Adv. Model. Earth Syst. 2018, 10, 2471–2494. [Google Scholar] [CrossRef]
- Post, H.; Vrugt, J.A.; Fox, A.; Vereecken, H.; Franssen, H.-J.H. Estimation of Community Land Model parameters for an improved assessment of net carbon fluxes at European sites. Biogeosciences 2017, 122, 661–689. [Google Scholar] [CrossRef] [Green Version]
- Mao, J.F.; Fu, W.T.; Shi, X.Y.; Ricciuto, D.M.; Fisher, J.B.; Dickinson, R.E.; Wei, Y.X.; Shem, W.; Piao, S.L.; Wang, K.C. Disentangling climatic and anthropogenic controls on global terrestrial evapotranspiration trends. Environ. Res. Lett. 2015, 10, 094008. [Google Scholar] [CrossRef]
- Shi, X.Y.; Mao, J.F.; Thornton, P.E.; Huang, M.Y. Spatiotemporal patterns of evapotranspiration in response to multiple environmental factors simulated by the community land model. Environ. Res. Lett. 2013, 8, 024012. [Google Scholar] [CrossRef]
- Lawrence, P.J.; Chase, T.N. Representing a new MODIS consistent land surface in the community land model (CLM 3.0). J. Geophys. Res. Biogeosci. 2007, 112, G01023. [Google Scholar] [CrossRef]
- Scanlon, B.R.; Zhang, Z.; Save, H.; Sun, A.Y.; Schmied, H.M.; van Beek, L.P.H.; Wiese, D.N.; Wada, Y.; Long, D.; Reedy, R.C.; et al. Global models underestimate large decadal declining and rising water storage trends relative to GRACE satellite data. Proc. Natl. Acad. Sci. USA 2018, 115, E1080–E1089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.W.; Lu, H.; Yang, K.; Han, M.L.; Wright, J.S.; Chen, Y.Y.; Yu, L.; Xu, S.M.; Huang, X.M.; Gong, W. The Evaluation of SMAP Enhanced Soil Moisture Products Using High-Resolution Model Simulations and In-Situ Observations on the Tibetan Plateau. Remote sens. 2018, 10, 535. [Google Scholar] [CrossRef] [Green Version]
- Farquhar, G.D.; von Caemmerer, S.; Berry, J.A. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collatz, G.J.; Ribas-Carbo, M.; Berry, J.A. Coupled photosynthesis-stomatal conductance model for leaves of C4 plants. Aust. J. Plant Physiol. 1992, 19, 519–538. [Google Scholar] [CrossRef]
- Thornton, P.E.; Lamarque, J.-F.; Rosenbloom, N.A.; Mahowald, N.M. Influence of carbon-nitrogen cycle coupling on land model response to CO2 fertilization and climate variability. Glob. Biogeochem. Cycles 2007, 21, B002868. [Google Scholar] [CrossRef]
- Ali, A.A.; Xu, C.; Rogers, A.; Fisher, R.A.; Wullschleger, S.D.; Massoud, E.; Vrugt, J.A.; Muss, J.D.; McDowell, N.; Fisher, J. A global scale mechanistic model of photosynthetic capacity (LUNA V1. 0). Geosci. Mod. Dev. 2016, 9, 587–606. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, D.M.; Fisher, R.A.; Koven, C.D.; Oleson, K.W.; Swenson, S.C.; Bonan, G.; Collier, N.; Ghimire, B.; van Kampenhout, L.; Kennedy, D.; et al. The Community Land Model version 5: Description of new features, benchmarking, and impact of forcing uncertainty. J. Adv. Model. Earth Syst. 2019, 11, 4245–4287. [Google Scholar] [CrossRef] [Green Version]
- Li, M.C.; Shu, J.J.; Sun, Y.R. Responses of specific leaf area of dominant tree species in Northeast China secondary forests to light intensity. Chin. J. Ecol. 2009, 28, 1437–1442. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Feng, Y.L. Relationship between leaf photosynthetic capacity and specific leaf weight, nitrogen content and allocation of two ficus species under different light intensities. J. Plant Physiol. Mol. Biol. 2004, 30, 269–276. [Google Scholar]
- Liu, W.D.; Li, S.F.; Su, L.; Su, J. Variation and correlations of leaf traits of two Taxus species with different shade tolerance along the light gradient. Pol. J. Ecol. 2013, 61, 329–339. [Google Scholar]
- Deloso, B.E.; Marler, T.E. Bi-Pinnate compound Serianthes nelsonii leaf-level plasticity magnifies leaflet-level plasticity. Biology 2020, 9, 333. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.; Huang, Y.M.; Pan, Y.P.; Xiang, X.; Huo, J.X.; Meng, D.H.; Wang, Y.Y.; Yun, C. Differential Investment Strategies in Leaf Economic Traits Across Climate Regions Worldwide. Front. Plant Sci. 2022, 13, 798035. [Google Scholar] [CrossRef]
- Heberling, J.M.; Fridley, J.D. Biogeographic constraints on the world-wide leaf economics spectrum. Glob. Ecol. Biogeogr. 2012, 21, 1137–1146. [Google Scholar] [CrossRef]
- Liu, K.J.; He, N.P.; Hou, J.H. Spatial patterns and influencing factors of specific leaf area in typical temperate forests. Acta Ecol. Sin. 2022, 42, 872–883. [Google Scholar] [CrossRef]
- Auger, S.; Shipley, B. Inter-specific and intra-specific trait variation along short environmental gradients in an old-growth temperate forest. J. Veg. Sci. 2013, 24, 419–428. [Google Scholar] [CrossRef]
- Mudrak, O.; Dolezal, J.; Vitova, A.; Leps, J. Variation in plant functional traits is best explained by the species identity: Stability of trait-based species ranking across meadow management regimes. Funct. Ecol. 2019, 33, 746–755. [Google Scholar] [CrossRef]
- Poorter, H.; Niinemets, U.; Poorter, L.; Wright, I.J.; Villar, R. Causes and consequences of variation in leaf mass per area (LMA): A meta-analysis. New Phytol. 2009, 182, 565–588. [Google Scholar] [CrossRef]
- Yang, Y.Z.; Gou, R.K.; Li, W.; Kassout, J.; Wu, J.; Wang, L.M.; Peng, C.H.; Lin, G.H. Leaf Trait Covariation and Its Controls: A Quantitative Data Analysis Along a Subtropical Elevation Gradient. J. Geophys. Res. Biogeosci. 2021, 126, e2021JG006378. [Google Scholar] [CrossRef]
- Koven, C.D.; Knox, R.G.; Fisher, R.A.; Chambers, J.Q.; Christoffersen, B.O.; Davies, S.J.; Detto, M.; Dietze, M.C.; Faybishenko, B.; Holm, J.; et al. Benchmarking and parameter sensitivity of physiological and vegetation dynamics using the Functionally Assembled Terrestrial Ecosystem Simulator (FATES) at Barro Colorado Island, Panama. Biogeosciences 2020, 17, 3017–3044. [Google Scholar] [CrossRef]
- De Wit, A.; Boogaard, H.; Fumagalli, D.; Janssen, S.; Knapen, R.; van Kraalingen, D.; Supit, I.; van der Raymond, W.; van Diepen, K. 25 years of the WOFOST cropping systems model. Agric. Syst. 2019, 168, 154–167. [Google Scholar] [CrossRef]
- Wu, R.; Yan, X.H.; Li, Y.N. Effects of Film Mulching and Supplementary Irrigation on Chlorophyll and Specific Leaf Area of Winter Wheat. Water Sav. Irrig. 2018, 9, 27–36. [Google Scholar]
- Cao, Y.H.; Shi, J.G.; Zhu, K.L.; Dong, S.T.; Liu, P.; Zhao, B.; Zhan, gJ.W. Effects of sowing depth on canopy structure and photosynthetic characteristics of summer maize. J. Maize. Sci. 2016, 24, 102–109. [Google Scholar] [CrossRef]
- Yang, Q.; Zhu, R.J.; Yang, C.Y.; Li, S.J.; Cheng, X.P. Variation in leaf functional traits of Bombax ceiba Linnaeus communities based on tree structure. Acta Ecol. Sin. 2022, 42, 2834–2842. [Google Scholar] [CrossRef]
- Zhang, L.; Luo, T.X.; Deng, K.M.; Li, W.H. Vertical variations in specific leaf area and leaf dry matter content with canopy height in Pinus yunnanensis. J. Beijing For. Univ. 2008, 30, 40–44. [Google Scholar] [CrossRef]
- Peng, X.; Yan, W.D.; Wang, F.Q.; Wang, G.J.; Yu, F.Y.; Zhao, M.F. Specific leaf area estimation model building based on leaf dry matter content of Cunninghamia lanceolata. Chin. J. Plant Ecol. 2018, 42, 209–219. [Google Scholar] [CrossRef] [Green Version]
- Rad, M.H.; Assare, M.H.; Banakar, M.H.; Soltani, M. Effects of Different Soil Moisture Regimes on Leaf Area Index, Specific Leaf Area and Water use Efficiency in Eucalyptus (Eucalyptus camaldulensis Dehnh) under Dry Climatic Conditions. Asian J. Plant Sci. 2011, 10, 294–300. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.Z.; Zhang, J.B.; Zhang, H.; Zhao, J.H.; Wu, Q.C.; Zhao, Z.H.; Cai, T.Y. Mechanisms for the relationships between water-use efficiency and carbon isotope composition and specific leaf area of maize (Zea mays L.) under water stress. Plant Growth Regul. 2015, 77, 233–243. [Google Scholar] [CrossRef]
- Huang, C.B.; Zeng, F.J.; Lei, J.Q. Growth and functional trait responses of Alhagi sparsifolia seedlings to water and nitrogen addition. Acta Prataculturae Sin. 2016, 25, 150–160. [Google Scholar] [CrossRef]
- Xiao, C.W.; Sun, O.J.; Zhou, G.S.; Zhao, J.Z.; Wu, G. Interactive effects of elevated CO2 and drought stress on leaf water potential and growth in Caragana intermedia. Trees 2005, 19, 712–721. [Google Scholar] [CrossRef]
- Wu, F.Z.; Bao, W.K.; Li, F.L.; Wu, N. Effects of water stress and nitrogen supply on leaf gas exchange and fluorescence parameters of Sophora davidii seedlings. Photosynthetica 2008, 46, 40–48. [Google Scholar] [CrossRef]
- Huang, J.Y.; Yu, H.L.; Yuan, Z.Y.; Li, L.H. Effects of long-term increased soil N on leaf traits of several species in typical Inner Mongolian grassland. Acta Ecol. Sin. 2009, 32, 1419–1427. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Yang, G.; Zhang, Y.; Qin, S.; Dong, J.; Cui, Y.; Liu, X.; Zheng, P.; Wang, R. Leaf Functional Traits of Two Species Affected by Nitrogen Addition Rate and Period Not Nitrogen Compound Type in a Meadow Grassland. Front. Plant Sci. 2022, 13, 841464. [Google Scholar] [CrossRef] [PubMed]
- Ratjen, A.M.; Kage, H. Is mutual shading a decisive factor for differences in overall canopy specific leaf area of winter wheat crops? Field Crops Res. 2013, 149, 338–346. [Google Scholar] [CrossRef]
- Van Bodegom, P.M.; Douma, J.C.; Verheijen, L.M. A fully traits-based approach to modeling global vegetation distribution. Proc. Natl. Acad. Sci. USA 2014, 111, 13733–13738. [Google Scholar] [CrossRef] [Green Version]
- Thomas, H.J.D.; Myers-Smith, I.H.; Bjorkman, A.D.; Elmendorf, S.C.; Blok, D.; Cornelissen, J.H.C.; Forbes, B.C.; Hollister, R.D.; Normand, S.; Prevey, J.S.; et al. Traditional plant functional groups explain variation in economic but not size-related traits across the tundra biome. Glob. Ecol. Biogeogr. 2019, 28, 78–95. [Google Scholar] [CrossRef] [Green Version]
- Fisher, R.A.; Muszala, S.; Verteinstein, M.; Lawrence, P.; Xu, C.; McDowell, N.G.; Knox, R.G.; Koven, C.; Holm, J.; Rogers, B.M.; et al. Taking off the training wheels: The properties of a dynamic vegetation model without climate envelopes, CLM4.5(ED). Geosci. Model Dev. 2015, 8, 3593–3619. [Google Scholar] [CrossRef] [Green Version]
- Verheijen, L.M.; Brovkin, V.; Aerts, R.; Bonisch, G.B.; Cornelissen, J.H.C.; Kattge, J.; Reich, P.B.; Wright, I.J.; van Bodegom, P.M. Impacts of trait variation through observed trait-climate relationships on performance of an Earth system model: A conceptual analysis. Biogeosciences 2013, 10, 5497–5515. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.Z.; Zhu, Q.A.; Peng, C.H.; Wang, H.; Xue, W.; Lin, G.H.; Wen, Z.M.; Chang, J.; Wang, M.; Liu, G.B.; et al. A novel approach for modelling vegetation distributions and analysing vegetation sensitivity through trait-climate relationships in China. Sci. Rep. 2016, 6, 24110. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site Name | QYZ | CBS | DHS | HBG |
|---|---|---|---|---|
| latitude (E) | 26.74 | 42.40 | 23.17 | 37.67 |
| longitude (N) | 115.06 | 128.10 | 112.57 | 101.33 |
| elevation (m) | 102 | 738 | 300 | 3327 |
| plant functional type | temperate NET | temperate BDT | temperate BET | C3 grass |
| simulated years | 2003–2008 | 2003–2008 | 2003–2008 | 2003–2008 |
| PFT | Maximum SLA Value (m2/g) | Minimum SLA Value (m2/g) | Coefficient of Variation (%) | Number of Samples (n) |
|---|---|---|---|---|
| temperate NET | 0.002 | 0.016 | 42.86 | 69 |
| boreal NET | 0.003 | 0.008 | 40.0 | 10 |
| boreal NDT | 0.005 | 0.024 | 40.0 | 11 |
| tropical BET | 0.004 | 0.027 | 28.57 | 103 |
| temperate BET | 0.0002 | 0.037 | 42.86 | 277 |
| tropical BDT | 0.007 | 0.032 | 41.18 | 9 |
| temperate BDT | 0.002 | 0.081 | 50.0 | 501 |
| boreal BDT | 0.005 | 0.028 | 25.0 | 4 |
| temperate BES | 0.004 | 0.048 | 47.06 | 221 |
| temperate BDS | 0.002 | 0.010 | 59.09 | 454 |
| boreal BDS | 0.004 | 0.025 | 21.05 | 6 |
| C3 grass | 0.002 | 0.082 | 57.14 | 822 |
| C4 grass | 0.003 | 0.052 | 47.37 | 126 |
| rainfed crop | 0.002 | 0.042 | 52.63 | 19 |
| Site | PFT | Relative Change | CV of SLA (%) | GPP (%) | Ac (%) | LAI (%) |
|---|---|---|---|---|---|---|
| QYZ | temperate NET | R1 | 42.9 | 7.0 | 43.9 | 14.1 |
| R2 | 60.0 | 18.5 | 31.0 | 61.7 | ||
| CBS | temperate BDT | R1 | 50.0 | 6.3 | 30.9 | 16.9 |
| R2 | 29.4 | 8.8 | 24.5 | 43.8 | ||
| DHS | temperate BET | R1 | 42.9 | 8.0 | 37.3 | 14.7 |
| R2 | 42.8 | 14.1 | 24.6 | 63.7 | ||
| HBG | C3 grass | R1 | 57.1 | 3.3 | 57.9 | 48.4 |
| R2 | 34.3 | 1.6 | 3.3 | 46.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, Y.; Zhang, L.; Li, P.; Ren, X.; He, H.; Lv, Y.; Ma, Y. Evaluation of the Community Land Model-Simulated Specific Leaf Area with Observations over China: Impacts on Modeled Gross Primary Productivity. Forests 2023, 14, 164. https://doi.org/10.3390/f14010164
Zheng Y, Zhang L, Li P, Ren X, He H, Lv Y, Ma Y. Evaluation of the Community Land Model-Simulated Specific Leaf Area with Observations over China: Impacts on Modeled Gross Primary Productivity. Forests. 2023; 14(1):164. https://doi.org/10.3390/f14010164
Chicago/Turabian StyleZheng, Yuanhao, Li Zhang, Pan Li, Xiaoli Ren, Honglin He, Yan Lv, and Yuping Ma. 2023. "Evaluation of the Community Land Model-Simulated Specific Leaf Area with Observations over China: Impacts on Modeled Gross Primary Productivity" Forests 14, no. 1: 164. https://doi.org/10.3390/f14010164
APA StyleZheng, Y., Zhang, L., Li, P., Ren, X., He, H., Lv, Y., & Ma, Y. (2023). Evaluation of the Community Land Model-Simulated Specific Leaf Area with Observations over China: Impacts on Modeled Gross Primary Productivity. Forests, 14(1), 164. https://doi.org/10.3390/f14010164