
Diversity of Beetles Captured in Pitfall Traps in the Șinca Old-Growth Forest, Brașov County, Romania: Forest Reserve versus Managed Forest



Abstract
:1. Introduction
2. Materials and Methods
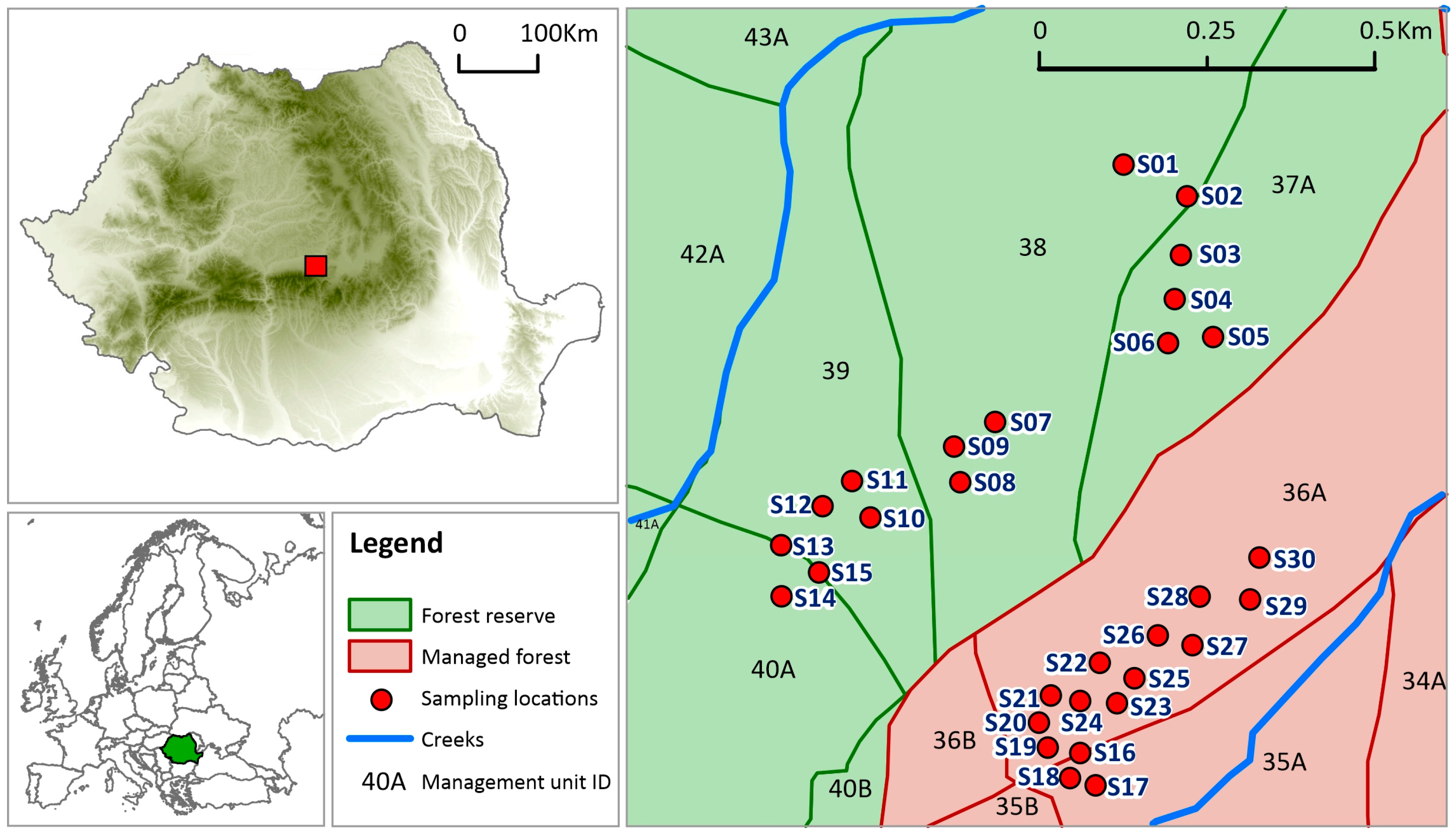
2.1. Study Area
2.2. Sampling Method and Field Design
2.3. Sorting and Identification of Beetles
2.4. Data Analysis
3. Results
3.1. Beetle Abundance
3.2. Beetle Dominance and Constancy
3.3. PERMANOVA Analysis of the Differences between the Two Beetle Communities
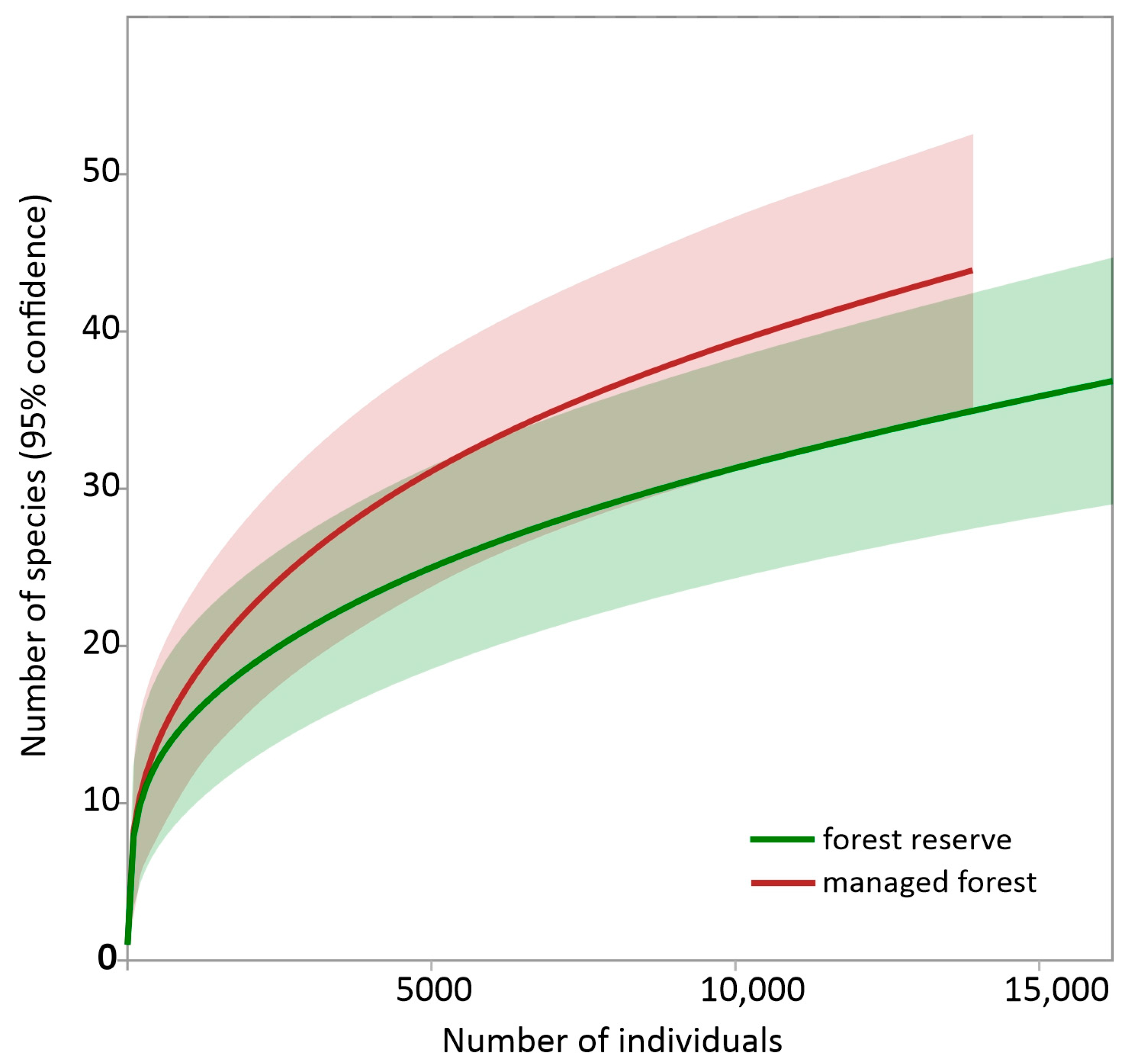
3.4. Beetle Community Diversity

4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lindenmayer, D.B.; Franklin, J.F.; Fischer, J. General management principles and a checklist of strategies to guide forest biodiversity conservation. Biol. Conserv. 2006, 131, 433–445. [Google Scholar] [CrossRef]
- Davies, S.J.; Abiem, I.; Abu Salim, K.; Aguilar, S.; Allen, D.; Alonso, A.; Anderson-Teixeira, K.; Andrade, A.; Arellano, G.; Ashton, P.S.; et al. ForestGEO: Understanding forest diversity and dynamics through a global observatory network. Biol. Conserv. 2021, 253, 108907. [Google Scholar] [CrossRef]
- Immerzeel, W.W.; Lutz, A.F.; Andrade, M.; Bahl, A.; Biemans, H.; Bolch, T.; Hyde, S.; Brumby, S.; Davies, B.J.; Elmore, A.C.; et al. Importance and vulnerability of the world’s water towers. Nature 2020, 577, 364–369. [Google Scholar] [CrossRef] [PubMed]
- Keenan, T.F.; Williams, C.A. The Terrestrial Carbon Sink. Annu. Rev. Env. Resour. 2018, 43, 219–243. [Google Scholar] [CrossRef]
- Europe, F. State of Europe’s forests 2020. Ministerial Conference on the Protection of Forests in Europe, Bratislava, Slovakia. Available online: https://foresteurope.org/state-of-europes-forests/ (accessed on 6 November 2022).
- Luick, R.; Reif, A.; Schneider, E.; Grossmann, M.; Fodor, E. Pădurile virgine în inima Europei. Importanța, situația curentă și viitorul pădurilor virgine ale României. Bucov. For. 2021, 21, 105–126. [Google Scholar] [CrossRef]
- Biriș, I.-A.; Teodosiu, M.; Turcu, D.-O.; Merce, O.; Lorenț, A.; Apostol, J.; Marcu, C. 24000 ha of primary beech forests, the Romanian proposal in UNESCO World Heritage. Bucov. For. 2016, 16, 107–116. [Google Scholar]
- Peterken, G.F. Natural Woodland: Ecology and Conservation in Northern Temperate Regions; Cambridge University Press: Cambridge, UK, 1996; p. 522. [Google Scholar]
- Paillet, Y.; Pernot, C.; Boulanger, V.; Debaive, N.; Fuhr, M.; Gilg, O.; Gosselin, F. Quantifying the recovery of old-growth attributes in forest reserves: A first reference for France. For. Ecol. Manag. 2015, 346, 51–64. [Google Scholar] [CrossRef]
- Sabatini, F.M.; Burrascano, S.; Keeton, W.S.; Levers, C.; Lindner, M.; Pötzschner, F.; Verkerk, P.J.; Bauhus, J.; Buchwald, E.; Chaskovsky, O.; et al. Where are Europe’s last primary forests? Divers. Distrib. 2018, 24, 1426–1439. [Google Scholar] [CrossRef] [Green Version]
- Burrascano, S.; Keeton, W.S.; Sabatini, F.M.; Blasi, C. Commonality and variability in the structural attributes of moist temperate old-growth forests: A global review. For. Ecol. Manag. 2013, 291, 458–479. [Google Scholar] [CrossRef]
- Duduman, G.; Barnoaiea, I.; Avăcăriței, D.; Barbu, C.-O.; Coșofreț, V.-C.; Dănilă, I.-C.; Duduman, M.-L.; Măciucă, A.; Drăgoi, M. Aboveground Biomass of Living Trees Depends on Topographic Conditions and Tree Diversity in Temperate Montane Forests from the Slătioara-Rarău Area (Romania). Forests 2021, 12, 1507. [Google Scholar] [CrossRef]
- Vandekerkhove, K.; De Keersmaeker, L.; Menke, N.; Meyer, P.; Verschelde, P. When nature takes over from man: Dead wood accumulation in previously managed oak and beech woodlands in North-western and Central Europe. For. Ecol. Manag. 2009, 258, 425–435. [Google Scholar] [CrossRef]
- Duduman, G. A forest management planning tool to create highly diverse uneven-aged stands. For. Int. J. For. Res. 2011, 84, 301–314. [Google Scholar] [CrossRef]
- Burrascano, S.; Chytrý, M.; Kuemmerle, T.; Giarrizzo, E.; Luyssaert, S.; Sabatini, F.M.; Blasi, C. Current European policies are unlikely to jointly foster carbon sequestration and protect biodiversity. Biol. Conserv. 2016, 201, 370–376. [Google Scholar] [CrossRef]
- Navarro, L.M.; Pereira, H.M. Rewilding Abandoned Landscapes in Europe. Ecosystems 2012, 15, 900–912. [Google Scholar] [CrossRef] [Green Version]
- Duduman, G.; Drăgoi, M. Forest management Planning—Spatial-Temporal Organization; Editura Universității “Ștefan cel Mare” Suceava: Suceava, Romania, 2019; p. X+209. [Google Scholar]
- Jonsson, B.G.; Siitonen, J. Dead wood and sustainable forest management. In Biodiversity in Dead Wood; Jonsson, B.G., Stokland, J.N., Siitonen, J., Eds.; Ecology, Biodiversity and Conservation; Cambridge University Press: Cambridge, UK, 2012; pp. 302–337. [Google Scholar]
- Li, W.-B.; Liu, N.-Y.; Wu, Y.-H.; Zhang, Y.-C.; Xu, Q.; Chu, J.; Wang, S.-Y.; Fang, J. Community composition and diversity of ground beetles (Coleoptera: Carabidae) in Yaoluoping National Nature Reserve. J. Insect Sci. 2017, 17, 114. [Google Scholar] [CrossRef] [Green Version]
- Olenici, N.; Fodor, E. The diversity of saproxylic beetles’ from the Natural Reserve Voievodeasa forest, North-Eastern Romania. Ann. For. Res. 2021, 64, 31–60. [Google Scholar] [CrossRef]
- Holliday, N. Species responses of carabid beetles (Coleoptera: Carabidae) during post-fire regeneration of boreal forest. Can. Entomol. 1991, 123, 1369–1389. [Google Scholar] [CrossRef]
- Wikars, L.-O. Clear-cutting before burning prevents establishment of the fire-adapted Agonum quadripunctatum (Coleoptera: Carabidae). In Proceedings of the Annales Zoologici Fennici, Helsinki, Finland, 8 December 1995; pp. 375–384. [Google Scholar]
- Saint-Germain, M.; Larrivée, M.; Drapeau, P.; Fahrig, L.; Buddle, C.M. Short-term response of ground beetles (Coleoptera: Carabidae) to fire and logging in a spruce-dominated boreal landscape. For. Ecol. Manag. 2005, 212, 118–126. [Google Scholar] [CrossRef]
- Błońska, E.; Bednarz, B.; Kacprzyk, M.; Piaszczyk, W.; Lasota, J. Effect of scots pine forest management on soil properties and carabid beetle occurrence under post-fire environmental conditions-a case study from Central Europe. For. Ecosyst. 2020, 7, 1–12. [Google Scholar] [CrossRef]
- Cobb, T.; Langor, D.; Spence, J. Biodiversity and multiple disturbances: Boreal forest ground beetle (Coleoptera: Carabidae) responses to wildfire, harvesting, and herbicide. Can. J. For. Res. 2007, 37, 1310–1323. [Google Scholar] [CrossRef]
- Tello, F.; González, M.E.; Micó, E.; Valdivia, N.; Torres, F.; Lara, A.; García-López, A. Short-Interval, Severe Wildfires Alter Saproxylic Beetle Diversity in Andean Araucaria Forests in Northwest Chilean Patagonia. Forests 2022, 13, 441. [Google Scholar] [CrossRef]
- Niemelä, J.; Langor, D.; Spence, J.R. Effects of clear-cut harvesting on boreal ground-beetle assemblages (Coleoptera: Carabidae) in western Canada. Conserv. Biol. 1993, 7, 551–561. [Google Scholar] [CrossRef]
- Work, T.T.; Shorthouse, D.P.; Spence, J.R.; Volney, W.J.A.; Langor, D. Stand composition and structure of the boreal mixedwood and epigaeic arthropods of the Ecosystem Management Emulating Natural Disturbance (EMEND) landbase in northwestern Alberta. Can. J. For. Res. 2004, 34, 417–430. [Google Scholar] [CrossRef]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Pearce, J.L.; Venier, L.A.; McKee, J.; Pedlar, J.; McKenney, D. Influence of habitat and microhabitat on carabid (Coleoptera: Carabidae) assemblages in four stand types. Can. Entomol. 2003, 135, 337–357. [Google Scholar] [CrossRef]
- Koivula, M.J. Useful model organisms, indicators, or both? Ground beetles (Coleoptera, Carabidae) reflecting environmental conditions. Zookeys 2011, 100, 287–317. [Google Scholar] [CrossRef] [Green Version]
- Spence, J.R.; Niemelä, J.K. Sampling carabid assemblages with pitfall traps: The madness and the method. Can. Entomol. 1994, 126, 881–894. [Google Scholar] [CrossRef]
- Jad’ud’ová, J.; Kanianska, R.; Kizeková, M.; Makovníková, J. Evaluation of Habitat Provision On the Basis of Carabidae Diversity in Slovak Permanent Grasslands. IOP Conf. Ser. Earth Environ. Sci. 2016, 44, 052031. [Google Scholar] [CrossRef]
- Nitu, E. Species diversity of the beetle fauna, a sensitive parameter for ecological monitoring. Maramures Mountains Nature Park (Romania). Transylv. Rev. Syst. Ecol. Res. 2008, 5, 143–154. [Google Scholar]
- Rainio, J.; Niemelä, J. Ground beetles (Coleoptera: Carabidae) as bioindicators. Biodivers. Conserv. 2003, 12, 487–506. [Google Scholar] [CrossRef]
- Kotze, D.J.; O’Hara, R.B. Species decline—But why? Explanations of carabid beetle (Coleoptera, Carabidae) declines in Europe. Oecologia 2003, 135, 138–148. [Google Scholar] [CrossRef]
- Avgın, S.S.; Luff, M.L. Ground beetles (Coleoptera: Carabidae) as bioindicators of human impact. Munis Entomol. Zool. 2010, 5, 209–215. [Google Scholar]
- Lobo, J.M. Estimation of dung beetle biomass (Coleoptera: Scarabaeoidea). Eur. J. Entomol. 1993, 90, 235–238. [Google Scholar]
- Byk, A.; Semkiw, P. Habitat preferences of the forest dung beetle Anoplotrupes stercorosus (Scriba, 1791)(Coleoptera: Geotrupidae) in the Białowieża Forest. Acta Sci. Pol. Silvarum Colendarum Ratio Et Ind. Lignaria 2010, 9, 17–28. [Google Scholar]
- Mittal, I.; Bhati, R. Food preference of some dung beetles (Coleoptera: Scarabaeidae). J. Entomol. Res. 1998, 22, 107–115. [Google Scholar]
- Creighton, J.C.; Bastarache, R.; Lomolino, M.V.; Belk, M.C. Effect of forest removal on the abundance of the endangered American burying beetle, Nicrophorus americanus (Coleoptera: Silphidae). J. Insect Conserv. 2009, 13, 37–43. [Google Scholar] [CrossRef]
- Rubner, K.; Rubner, H.; Rittershofer, F. Grundlagen des naturnahen Waldbaus in Europa. Forstwiss. Cent. 1968, 87, 8–36. [Google Scholar] [CrossRef]
- Keeton, W.S.; Chernyavskyy, M.; Gratzer, G.; Main-Knorn, M.; Shpylchak, M.; Bihun, Y. Structural characteristics and aboveground biomass of old-growth spruce–fir stands in the eastern Carpathian mountains, Ukraine. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2010, 144, 148–159. [Google Scholar] [CrossRef]
- European Economic Area. Developing a Forest Naturalness Indicator for Europe. Concept and Methodology for a High Nature Value (HNV) Forest Indicator. EEA Technical Report No 13/2014; Publications Office of the European Union: Luxembourg, 2014; p. 64. [Google Scholar]
- Chowdhury, S.; Jennions, M.D.; Zalucki, M.P.; Maron, M.; Watson, J.E.M.; Fuller, R.A. Protected areas and the future of insect conservation. Trends Ecol. Evol. 2022, 38, 85–95. [Google Scholar] [CrossRef]
- Canelles, Q.; Aquilué, N.; James, P.M.A.; Lawler, J.; Brotons, L. Global review on interactions between insect pests and other forest disturbances. Landsc. Ecol. 2021, 36, 945–972. [Google Scholar] [CrossRef]
- Jouveau, S.; Toïgo, M.; Giffard, B.; Castagneyrol, B.; van Halder, I.; Vétillard, F.; Jactel, H. Carabid activity-density increases with forest vegetation diversity at different spatial scales. Insect Conserv. Divers. 2020, 13, 36–46. [Google Scholar] [CrossRef]
- Oettel, J.; Lapin, K. Linking forest management and biodiversity indicators to strengthen sustainable forest management in Europe. Ecol. Indic. 2021, 122, 107275. [Google Scholar] [CrossRef]
- Nițu, E.; Olenici, N.; Popa, I.; Nae, A.; Biriș, I.-A. Soil and saproxylic species (Coleoptera, Collembola, Araneae) in primeval forests from the northern part of South-Easthern Carpathians. Ann. For. Res. 2010, 52, 27–54. [Google Scholar]
- Nitzu, E.; Olenici, N. The first study on the beetle fauna in the Giumalau spruce primeval forest (Eastern Carpathians, Romania), mainly based on a quantitative analysis of terrestrial and saproxylic species. In Buse J., Saproxylic Beetles–Their Role and Diversity in European Woodland and Tree Habitats, Proceedings of the 5th Symposium and Workshop on the Conservation of Saproxylic Beetles, Leuphana University of Lüneburg, Germany, 14–16 June 2008; Pensoft: Sofia, Bulgaria, 2009; pp. 27–48. [Google Scholar]
- Sima, G.; Gherman, S.; Banu, C. Forest Management Plan of the Public and Private Property Forest Fund of the Șinca Commune, Brașov County; S.C. Omni S.R.L.: Timișoara, Romania, 2015; p. 546. [Google Scholar]
- Guțu, O.; Urdea, S.; Gotea, C.; Lorenț, A.; Struțeanu, M.; Matei, S. Șinca Old Gowth Forest. 2015, 2. Available online: https://padurivirgine.ro/wp-content/uploads/2021/06/pliant-Codrii-Seculari-de-la-Sinca-ME_GO-revizuit.pdf (accessed on 3 November 2022).
- Petritan, I.C.; Commarmot, B.; Hobi, M.L.; Petritan, A.M.; Bigler, C.; Abrudan, I.V.; Rigling, A. Structural patterns of beech and silver fir suggest stability and resilience of the virgin forest Sinca in the Southern Carpathians, Romania. For. Ecol. Manag. 2015, 356, 184–195. [Google Scholar] [CrossRef]
- M.M.A.P. Technical rules on the choice and application of treatments and the Guide to good practice on the choice and application of treatments. Monitorul Oficial al României 2022, 190(994bis), 10–70. [Google Scholar]
- Bense, U. Longhorn Beetles: Illustrated Key to the Cerambycidae an Vesperidae of Europe; Margraf Verlag: Weikersheim, Germany, 1995; p. 512. [Google Scholar]
- Lohse, G.A.; Freude, H.; Harde, K.W.; Klausnitzer, B. Die Käfer Mitteleuropas. 12: Supplementbd. 1: Mit Katalogteil; Spektrum Akad; Verlag: Krefeld, Germany, 1989; p. 346. [Google Scholar]
- Löbl, I.; Smetana, A. Catalogue of Palaearctic Coleoptera; Apollo Books: Stenstrup, Denmark, 2003; Volume 6. [Google Scholar]
- Trautner, J.; Geigenmüller, K. Tiger Beetles, Ground Beetles. Illustrated Key to the Cicindelidae and Carabidae of Europe; Triops Verlag: Nördlingen, Germany, 1987; p. 488. [Google Scholar]
- Chao, A.; Gotelli, N.J.; Hsieh, T.C.; Sander, E.L.; Ma, K.H.; Colwell, R.K.; Ellison, A.M. Rarefaction and extrapolation with Hill numbers: A framework for sampling and estimation in species diversity studies. Ecol. Monogr. 2014, 84, 45–67. [Google Scholar] [CrossRef] [Green Version]
- Colwell, R.K.; Coddington, J.A. Estimating terrestrial biodiversity through extrapolation. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1994, 345, 101–118. [Google Scholar] [CrossRef] [Green Version]
- Magurran, A.E. Measuring Biological Diversity; Wiley: Hoboken, NJ, USA, 2004; p. 256. [Google Scholar]
- Engelmann, H.D. Zur Dominanzklassifizierung von Bodenarthropoden. Pedobiologia 1978, 18, 378–380. [Google Scholar]
- Anderson, M.J. Permutational Multivariate Analysis of Variance (PERMANOVA). Wiley StatsRef: Statistics Reference Online. pp. 1–15. Available online: https://onlinelibrary.wiley.com/doi/full/10.1002/9781118445112.stat07841 (accessed on 2 November 2022).
- Zar, J.H. Biostatistical Analysis, 5th ed.; Pearson Prentice Hall: Upper Saddle River, NJ, USA, 2010. [Google Scholar]
- Taguchi, Y.H.; Oono, Y. Relational patterns of gene expression via non-metric multidimensional scaling analysis. Bioinformatics 2005, 21, 730–740. [Google Scholar] [CrossRef]
- Commission, E.; Environment, D.-G.f.; Sundseth, K. The EU Birds and Habitats Directives: For Nature and People in Europe; Publications Office: Luxembourg, Belgium, 2018. [Google Scholar]
- Schulze, E.D. Effects of forest management on biodiversity in temperate deciduous forests: An overview based on Central European beech forests. J. Nat. Conserv. 2018, 43, 213–226. [Google Scholar] [CrossRef]
- Kazerani, F.; Farashiani, M.E.; Sagheb-Talebi, K.; Thorn, S. Forest management alters alpha-, beta-, and gamma diversity of saproxylic flies (Brachycera) in the Hyrcanian forests, Iran. For. Ecol. Manag. 2021, 496, 119444. [Google Scholar] [CrossRef]
- McGeoch, M.A.; Schroeder, M.; Ekbom, B.; Larsson, S. Saproxylic beetle diversity in a managed boreal forest: Importance of stand characteristics and forestry conservation measures. Divers. Distrib. 2007, 13, 418–429. [Google Scholar] [CrossRef]
- Janssen, P.; Fortin, D.; Hébert, C. Beetle diversity in a matrix of old-growth boreal forest: Influence of habitat heterogeneity at multiple scales. Ecography 2009, 32, 423–432. [Google Scholar] [CrossRef]
- Rischen, T.; Frenzel, T.; Fischer, K. Biodiversity in agricultural landscapes: Different non-crop habitats increase diversity of ground-dwelling beetles (Coleoptera) but support different communities. Biodivers. Conserv. 2021, 30, 3965–3981. [Google Scholar] [CrossRef]
- Brygadyrenko, V. Evaluation of ecological niches of abundant species of Poecilus and Pterostichus (Coleoptera: Carabidae) in forests of steppe zone of Ukraine. Entomol. Fenn. 2016, 27, 81–100. [Google Scholar] [CrossRef] [Green Version]
- Secretariat, G. GBIF Backbone Taxonomy. Checklist Dataset. Available online: https://www.gbif.org/dataset (accessed on 3 December 2022).
- Matern, A.; Drees, C.; Kleinwächter, M.; Assmann, T. Habitat modelling for the conservation of the rare ground beetle species Carabus variolosus (Coleoptera, Carabidae) in the riparian zones of headwaters. Biol. Conserv. 2007, 136, 618–627. [Google Scholar] [CrossRef]
- Esh, M.; Oxbrough, A. Macrohabitat associations and phenology of carrion beetles (Coleoptera: Silphidae, Leiodidae: Cholevinae). J. Insect Conserv. 2021, 25, 123–136. [Google Scholar] [CrossRef]
- Leather, S.R.; Day, K.R.; Salisbury, A.N. The biology and ecology of the large pine weevil, Hylobius abietis (Coleoptera: Curculionidae): A problem of dispersal? Bull. Entomol. Res. 1999, 89, 3–16. [Google Scholar] [CrossRef]
- Starzyk, J. Bionomics, ecology and economic importance of the fir weevil, Pissodes piceae (III.)(Col., Curculionidae) in mountain forests. J. Appl. Entomol. 1996, 120, 65–75. [Google Scholar] [CrossRef]
- Hendriks, P. Life cycle length of the lesser stag beetle (Coleoptera: Lucanidae: Dorcus parallelipipedus). Entomol. Ber. 2019, 79, 208–216. [Google Scholar]
- Kašák, J.; Mazalová, M.; Šipoš, J.; Foit, J.; Hučín, M.; Kuras, T. Habitat preferences of Ceruchus chrysomelinus, an endangered relict beetle of the natural Central European montane forests. Insect Conserv. Divers. 2019, 12, 206–215. [Google Scholar] [CrossRef]
- Yakovlev, E.; Hokkanen, T. Cyllodes ater (Coleoptera, Nitidulidae) found again in Finland. Entomol. Fenn. 1994, 5, 203–204. [Google Scholar] [CrossRef] [Green Version]
- Bekchiev, R.; Crevecoeur, L.; Gielen, K.; Bosmans, B.; Smets, K.; Kostova, R. One Hundred Years of Solitude: Rediscovery of the Rare and Protected Beetle Rhysodes sulcatus (Fabricius, 1787)(Coleoptera: Rhysodidae) in Bulgaria. Acta Zool. Bulg. 2020, 72, 381–384. [Google Scholar]
- Coddington, J.A.; Agnarsson, I.; Miller, J.A.; Kuntner, M.; Hormiga, G. Undersampling bias: The null hypothesis for singleton species in tropical arthropod surveys. J. Anim. Ecol. 2009, 78, 573–584. [Google Scholar] [CrossRef] [PubMed]
- Wong, M.K.L.; Guénard, B.; Lewis, O.T. Trait-based ecology of terrestrial arthropods. Biol. Rev. 2019, 94, 999–1022. [Google Scholar] [CrossRef] [Green Version]
- Washington, H.G. Diversity, biotic and similarity indices: A review with special relevance to aquatic ecosystems. Water Res. 1984, 18, 653–694. [Google Scholar] [CrossRef]
- Tóthmérész, B. Comparison of different methods for diversity ordering. J. Veg. Sci. 1995, 6, 283–290. [Google Scholar] [CrossRef]
- Byriel, D.B.; Ro-Poulsen, H.; Kepfer-Rojas, S.; Hansen, A.K.; Hansen, R.R.; Justesen, M.J.; Kristensen, E.; Møller, C.B.; Schmidt, I.K. Contrasting responses of multiple insect taxa to common heathland management regimes and old-growth successional stages. Biodivers. Conserv. 2022, 31. [Google Scholar] [CrossRef]
- Larcenaire, C.; Wang, F.; Holásková, I.; Turcotte, R.; Gutensohn, M.; Park, Y.-L. Effects of Forest Management on the Insect Assemblage of Black Cherry (Prunus serotina) in the Allegheny National Forest. Plants 2022, 11, 2596. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stand | Management Units | Area (ha) | Altitude (m) | Expo-sition | Slope (g) | Composition (%) | Age (Years) | Canopy Cover | Mean of Volume of Living Trees (m3 × ha−1) | Mean of Volume of Dead Wood (m3 × ha−1) |
|---|---|---|---|---|---|---|---|---|---|---|
| Forest reserve | 37A, 38, 39, 40A | 98.2 | 850–1350 | N | 35–40 | 70 Fagus sylvatica 30 Abies alba | 100–180 | 0.7 | 574 | 134.9 [53] |
| Managed forest | 35A, 36A | 45.4 | 950–1250 | N | 25–35 | 60 Fagus sylvatica 40 Abies alba | 130–180 | 0.5 | 382 | No data |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Isaia, G.; Dragomir, I.-M.; Duduman, M.-L. Diversity of Beetles Captured in Pitfall Traps in the Șinca Old-Growth Forest, Brașov County, Romania: Forest Reserve versus Managed Forest. Forests 2023, 14, 60. https://doi.org/10.3390/f14010060
Isaia G, Dragomir I-M, Duduman M-L. Diversity of Beetles Captured in Pitfall Traps in the Șinca Old-Growth Forest, Brașov County, Romania: Forest Reserve versus Managed Forest. Forests. 2023; 14(1):60. https://doi.org/10.3390/f14010060
Chicago/Turabian StyleIsaia, Gabriela, Ionuț-Marian Dragomir, and Mihai-Leonard Duduman. 2023. "Diversity of Beetles Captured in Pitfall Traps in the Șinca Old-Growth Forest, Brașov County, Romania: Forest Reserve versus Managed Forest" Forests 14, no. 1: 60. https://doi.org/10.3390/f14010060
APA StyleIsaia, G., Dragomir, I. -M., & Duduman, M. -L. (2023). Diversity of Beetles Captured in Pitfall Traps in the Șinca Old-Growth Forest, Brașov County, Romania: Forest Reserve versus Managed Forest. Forests, 14(1), 60. https://doi.org/10.3390/f14010060