
Phosphorus Rather than Nitrogen Addition Changed Soil Cyanobacterial Community in a Tropical Secondary Forest of South China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Experimental Design
2.3. Soil Sampling
2.4. Soil Chemical Analysis
2.5. DNA Extraction and Analysis
2.6. Statistical Analysis
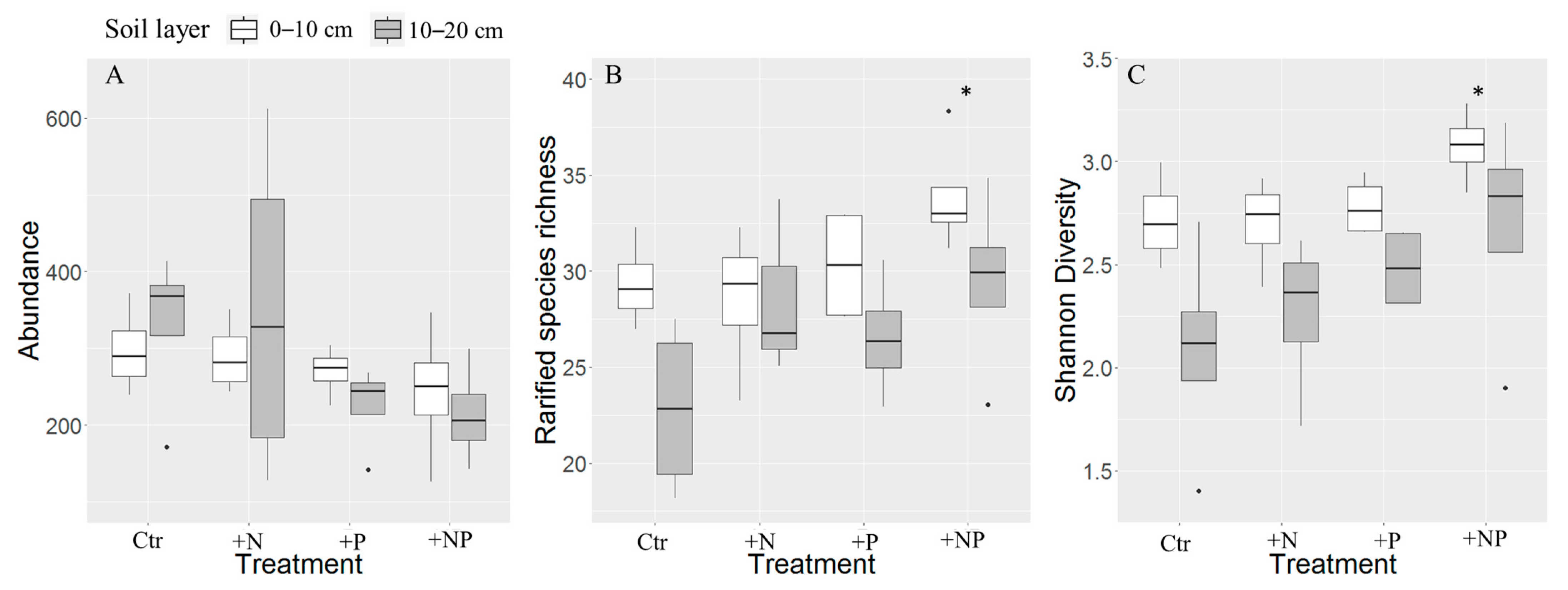
3. Results
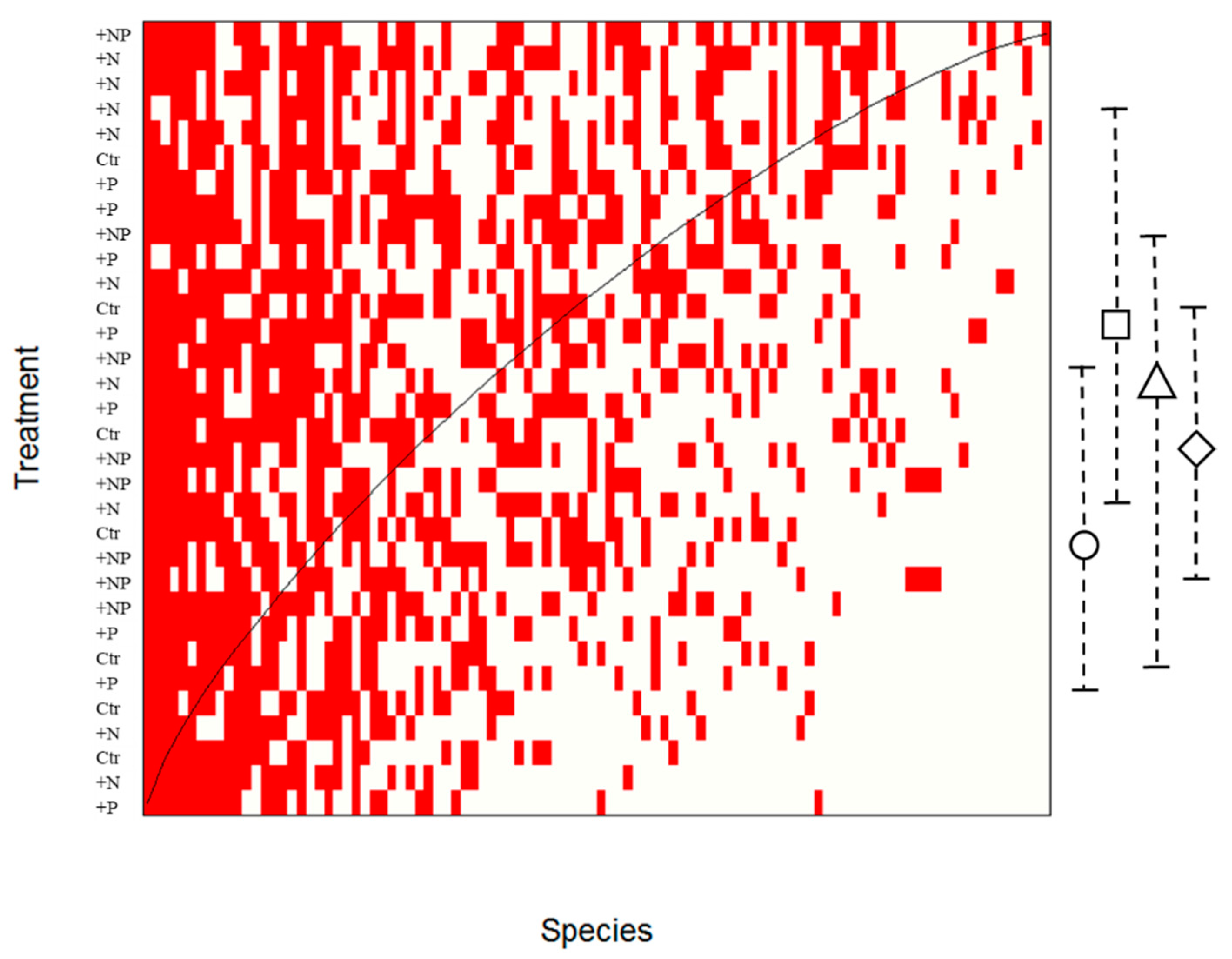
3.1. Soil Cyanobacteria Detected at 0–10 and 10–20 cm Soil Layer
3.2. Effects of NP Addition on Soil Chemical Property
3.3. Effects of NP Addition on Soil Cyanobacterial Community
4. Discussion
4.1. Cyanobacterial Communities at Different Soil Depths
4.2. N Addition Had Minor Effects on Soil Cyanobacterial Community
4.3. P Addition Significantly Changed Soil Cyanobacterial Community
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Whitton, B.A.; Potts, M. Introduction to the Cyanobacteria. In Ecology of Cyanobacteria II; Springer: Dordrecht, The Netherlands, 2012; pp. 1–13. ISBN 978-94-007-3855-3. [Google Scholar]
- Shih, P.M.; Hemp, J.; Ward, L.M.; Matzke, N.J.; Fischer, W.W. Crown group Oxyphotobacteria postdate the rise of oxygen. Geobiology 2017, 15, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Soo, R.M.; Hemp, J.; Parks, D.H.; Fischer, W.W.; Hugenholtz, P. On the origins of oxygenic photosynthesis and aerobic respiration in Cyanobacteria. Science 2017, 355, 1436–1439. [Google Scholar] [CrossRef] [PubMed]
- Moreira, C.; Vasconcelos, V.; Antunes, A. Phylogeny and Biogeography of Cyanobacteria and Their Produced Toxins. Mar. Drugs 2013, 11, 4350–4369. [Google Scholar] [CrossRef] [PubMed]
- Cano-Diaz, C.; Maestre, F.T.; Eldridge, D.J.; Singh, B.K.; Bardgett, R.D.; Fierer, N.; Delgado-Baquerizo, M. Contrasting environmental preferences of photosynthetic and non-photosynthetic soil cyanobacteria across the globe. Glob. Ecol. Biogeogr. 2020, 29, 2025–2038. [Google Scholar] [CrossRef]
- Garcia-Pichel, F.; Belnap, J.; Neuer, S.; Schanz, F. Estimates of global cyanobacterial biomass and its distribution. Algol. Stud. 1995, 109, 213–227. [Google Scholar] [CrossRef]
- Pankratova, E.M. Functioning of cyanobacteria in soil ecosystems. Eurasian Soil Sci. 2006, 39, 118–127. [Google Scholar] [CrossRef]
- Jassey, V.E.J.; Walcker, R.; Kardol, P.; Geisen, S.; Heger, T.; Lamentowicz, M.; Hamard, S.; Lara, E. Contribution of soil algae to the global carbon cycle. New Phytol. 2022, 234, 64–76. [Google Scholar] [CrossRef]
- Lee, S.-M.; Ryu, C.-M. Algae as New Kids in the Beneficial Plant Microbiome. Front. Plant Sci. 2021, 12, 599742. [Google Scholar] [CrossRef]
- Starks, T.L.; Shubert, L.E.; Trainor, F.R. Ecology of soil algae: A review. Phycologia 1981, 20, 65–80. [Google Scholar] [CrossRef]
- Seitz, S.; Nebel, M.; Goebes, P.; Kappeler, K.; Schmidt, K.; Shi, X.; Song, Z.; Webber, C.L.; Weber, B.; Scholten, T. Bryophyte-dominated biological soil crusts mitigate soil erosion in an early successional Chinese subtropical forest. Biogeosciences 2017, 14, 5775–5788. [Google Scholar] [CrossRef]
- Kuwabara, T.; Iwamoto, K.; Hara, H.; Yamaguchi, T.; Mohamad, S.; Abdullah, N.; Khudzari, J.M.; Othman, F. Prevention of soil erosion using microalgae in Malaysia. IOP Conf. Ser. Mater. Sci. Eng. 2021, 1051, 012047. [Google Scholar] [CrossRef]
- Lynch, J.; Bragg, E. Microorganisms and soil aggregate stability. Adv. Soil Sci. 1985, 2, 133–171. [Google Scholar]
- Johansen, J.R. Cryptogamic crusts of semiarid and arid lands of north America. J. Phycol. 1993, 29, 140–147. [Google Scholar] [CrossRef]
- Dixon, R.K.; Solomon, A.M.; Brown, S.; Houghton, R.A.; Trexier, M.C.; Wisniewski, J. Carbon pools and flux of global forest ecosystems. Science 1994, 263, 185–190. [Google Scholar] [CrossRef]
- Grace, J.; Malhi, Y.; Meir, P.; Higuchi, N. Terrestrial Global Productivity; Roy, J., Saugier, B., Mooney, H.A., Eds.; Academic Press: San Diego, CA, USA, 2001; pp. 401–426. ISBN 978-0-12-505290-0. [Google Scholar]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Bonan, G.B. Forests and Climate Change: Forcings, Feedbacks, and the Climate Benefits of Forests. Science 2008, 320, 1444–1449. [Google Scholar] [CrossRef]
- Rehakova, K.; Chlumska, Z.; Dolezal, J. Soil Cyanobacterial and Microalgal Diversity in Dry Mountains of Ladakh, NW Himalaya, as Related to Site, Altitude, and Vegetation. Microb. Ecol. 2011, 62, 337–346. [Google Scholar] [CrossRef]
- Rehakova, K.; Chronakova, A.; Kristufek, V.; Kuchtova, B.; Capkova, K.; Scharfen, J.; Capek, P.; Dolezal, J. Bacterial community of cushion plant Thylacospermum ceaspitosum on elevational gradient in the Himalayan cold desert. Front. Microbiol. 2015, 6, 304. [Google Scholar] [CrossRef]
- Janatkova, K.; Rehakova, K.; Dolezal, J.; Simek, M.; Chlumska, Z.; Dvorsky, M.; Kopecky, M. Community structure of soil phototrophs along environmental gradients in arid Himalaya. Environ. Microbiol. 2013, 15, 2505–2516. [Google Scholar] [CrossRef]
- Lindo, Z.; Nilsson, M.-C.; Gundale, M.J. Bryophyte-cyanobacteria associations as regulators of the northern latitude carbon balance in response to global change. Glob. Chang. Biol. 2013, 19, 2022–2035. [Google Scholar] [CrossRef]
- Soo, R.M.; Woodcroft, B.J.; Parks, D.H.; Tyson, G.W.; Hugenholtz, P. Back from the dead; the curious tale of the predatory cyanobacterium Vampirovibrio chlorellavorus. PeerJ 2015, 3, e968. [Google Scholar] [CrossRef] [PubMed]
- Benavent-Gonzalez, A.; Raggio, J.; Villagra, J.; Manuel Blanquer, J.; Pintado, A.; Rozzi, R.; Allan Green, T.G.; Sancho, L.G. High nitrogen contribution by Gunnera magellanica and nitrogen transfer by mycorrhizas drive an extraordinarily fast primary succession in sub-Antarctic Chile. New Phytol. 2019, 223, 661–674. [Google Scholar] [CrossRef]
- Gaysina, L.A.; Saraf, A.; Singh, P. Cyanobacteria; Mishra, A.K., Tiwari, D.N., Rai, A.N., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 1–28. ISBN 978-0-12-814667-5. [Google Scholar]
- Mühlsteinová, R.; Hauer, T. Pilot survey of cyanobacterial diversity from the neighborhood of San Gerardo de Rivas, Costa Rica with a brief summary of current knowledge of terrestrial cyanobacteria in Central America. Braz. J. Bot. 2013, 36, 299–307. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Aber, J.D.; Howarth, R.W.; Likens, G.E.; Matson, P.A.; Schindler, D.W.; Schlesinger, W.H.; Tilman, D. Human alteration of the global nitrogen cycle: Sources and consequences. Ecol. Appl. 1997, 7, 737–750. [Google Scholar] [CrossRef]
- Liu, L.; Xu, W.; Lu, X.; Zhong, B.; Guo, Y.; Lu, X.; Zhao, Y.; He, W.; Wang, S.; Zhang, X.; et al. Exploring global changes in agricultural ammonia emissions and their contribution to nitrogen deposition since 1980. Proc. Natl. Acad. Sci. USA 2022, 119, e2121998119. [Google Scholar] [CrossRef]
- LeBauer, D.S.; Treseder, K.K. Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed. Ecology 2008, 89, 371–379. [Google Scholar] [CrossRef]
- Mo, J.; Zhang, W.; Zhu, W.; Gundersen, P.; Fang, Y.; Li, D.; Wang, H. Nitrogen addition reduces soil respiration in a mature tropical forest in southern China. Glob. Chang. Biol. 2008, 14, 403–412. [Google Scholar] [CrossRef]
- Lu, X.; Mo, J.; Gilliam, F.S.; Zhou, G.; Fang, Y. Effects of experimental nitrogen additions on plant diversity in an old-growth tropical forest. Glob. Chang. Biol. 2010, 16, 2688–2700. [Google Scholar] [CrossRef]
- Lu, X.; Mao, Q.; Gilliam, F.S.; Luo, Y.; Mo, J. Nitrogen deposition contributes to soil acidification in tropical ecosystems. Glob. Chang. Biol. 2014, 20, 3790–3801. [Google Scholar] [CrossRef]
- Hobbie, S.E.; Vitousek, P.M. Nutrient limitation of decomposition in Hawaiian forests. Ecology 2000, 81, 1867–1877. [Google Scholar] [CrossRef]
- Wardle, D.A.; Walker, L.R.; Bardgett, R.D. Ecosystem properties and forest decline in contrasting long-term chronosequences. Science 2004, 305, 509–513. [Google Scholar] [CrossRef] [PubMed]
- Camenzind, T.; Haettenschwiler, S.; Treseder, K.K.; Lehmann, A.; Rillig, M.C. Nutrient limitation of soil microbial processes in tropical forests. Ecol. Monogr. 2018, 88, 4–21. [Google Scholar] [CrossRef]
- Huang, W.-J.; Zhou, G.-Y.; Liu, J.-X. Nitrogen and phosphorus status and their influence on aboveground production under increasing nitrogen deposition in three successional forests. Acta Oecologica-Int. J. Ecol. 2012, 44, 20–27. [Google Scholar] [CrossRef]
- Li, J.; Li, Z.; Wang, F.; Zou, B.; Chen, Y.; Zhao, J.; Mo, Q.; Li, Y.; Li, X.; Xia, H. Effects of nitrogen and phosphorus addition on soil microbial community in a secondary tropical forest of China. Biol. Fertil. Soils 2015, 51, 207–215. [Google Scholar] [CrossRef]
- Ma, X.; Zhou, Z.; Chen, J.; Xu, H.; Ma, S.; Dippold, M.A.; Kuzyakov, Y. Long-term nitrogen and phosphorus fertilization reveals that phosphorus limitation shapes the microbial community composition and functions in tropical montane forest soil. Sci. Total Environ. 2023, 854, 158709. [Google Scholar] [CrossRef]
- Ma, S.; Chen, X.; Su, H.; Xing, A.; Chen, G.; Zhu, J.; Zhu, B.; Fang, J. Phosphorus addition decreases soil fungal richness and alters fungal guilds in two tropical forests. Soil Biol. Biochem. 2022, 175, 108836. [Google Scholar] [CrossRef]
- Viles, H.A. Blue-green algae and terrestrial limestone weathering on Aldabra Atoll: An SEM and light microscope study. Earth Surf. Process. Landf. 1987, 12, 319–330. [Google Scholar] [CrossRef]
- Depaivaazevedo, M.T. Edaphic blue-green-algae from the São-Paulo Botanical Graden, Brazil. Arch. Hydrobiol. 1991, 64, 503–526. [Google Scholar]
- Liu, L.; Gao, Y.; Yang, W.; Liu, J.; Wang, Z. Community metagenomics reveals the processes of nutrient cycling regulated by microbial functions in soils with P fertilizer input. Plant Soil 2023, 1–16. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, H.; Wang, J.; Wang, J.; Zhang, Y. Long-term fertilization with high nitrogen rates decreased diversity and stability of diazotroph communities in soils of sweet potato. Appl. Soil Ecol. 2022, 170, 104266. [Google Scholar] [CrossRef]
- Liengen, T. Environmental factors influencing the nitrogen fixation activity of free-living terrestrial cyanobacteria from a high arctic area, Spitsbergen. Can. J. Microbiol. 1999, 45, 573–581. [Google Scholar] [CrossRef]
- Wang, J.; Bao, J.T.; Su, J.Q.; Li, X.R.; Chen, G.X.; Ma, X.F. Impact of inorganic nitrogen additions on microbes in biological soil crusts. Soil Biol. Biochem. 2015, 88, 303–313. [Google Scholar] [CrossRef]
- Kuske, C.R.; Yeager, C.M.; Johnson, S.; Ticknor, L.O.; Belnap, J. Response and resilience of soil biocrust bacterial communities to chronic physical disturbance in arid shrublands. ISME J. 2012, 6, 886–897. [Google Scholar] [CrossRef]
- Zhang, J.; Zhou, J.; Lambers, H.; Li, Y.; Li, Y.; Qin, G.; Wang, M.; Wang, J.; Li, Z.; Wang, F. Nitrogen and phosphorus addition exerted different influences on litter and soil carbon release in a tropical forest. Sci. Total Environ. 2022, 832, 155049. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Sayer, E.J.; Li, Z.; Mo, Q.; Li, Y.; Ding, Y.; Wang, J.; Lu, X.; Tang, J.; Wang, F. Nutrient limitation of woody debris decomposition in a tropical forest: Contrasting effects of N and P addition. Funct. Ecol. 2016, 30, 295–304. [Google Scholar] [CrossRef]
- Wang, F.; Li, J.; Wang, X.; Zhang, W.; Zou, B.; Neher, D.A.; Li, Z. Nitrogen and phosphorus addition impact soil N2O emission in a secondary tropical forest of South China. Sci. Rep. 2014, 4, 5615. [Google Scholar] [CrossRef]
- Ding, M.; Yi, W.; Liao, L.; Martens, R.; Insam, H. Effect of afforestation on microbial biomass and acitivity in soils of tropical China. Soil Biol. Biochem. 1992, 24, 865–872. [Google Scholar] [CrossRef]
- Ren, H.; Li, Z.; Shen, W.; Yu, Z.; Peng, S.; Liao, C.; Ding, M.; Wu, J. Changes in biodiversity and ecosystem function during the restoration of a tropical forest in south China. Sci. China Ser. C Life Sci. 2007, 50, 277–284. [Google Scholar] [CrossRef]
- Liu, L.; Gundersen, P.; Zhang, T.; Mo, J. Effects of phosphorus addition on soil microbial biomass and community composition in three forest types in tropical China. Soil Biol. Biochem. 2012, 44, 31–38. [Google Scholar] [CrossRef]
- Wright, S.J.; Yavitt, J.B.; Wurzburger, N.; Turner, B.L.; Tanner, E.V.J.; Sayer, E.J.; Santiago, L.S.; Kaspari, M.; Hedin, L.O.; Harms, K.E.; et al. Potassium, phosphorus, or nitrogen limit root allocation, tree growth, or litter production in a lowland tropical forest. Ecology 2011, 92, 1616–1625. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: http://www.R-project.org/ (accessed on 1 June 2023).
- Amend, A.S.; Seifert, K.A.; Bruns, T.D. Quantifying microbial communities with 454 pyrosequencing: Does read abundance count? Mol. Ecol. 2010, 19, 5555–5565. [Google Scholar] [CrossRef] [PubMed]
- Camenzind, T.; Hempel, S.; Homeier, J.; Horn, S.; Velescu, A.; Wilcke, W.; Rillig, M.C. Nitrogen and phosphorus additions impact arbuscular mycorrhizal abundance and molecular diversity in a tropical montane forest. Glob. Chang. Biol. 2014, 20, 3646–3659. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Liu, X.; Zhou, S. Nitrogen addition altered the plant-arbuscular mycorrhizal fungi network through reducing redundant interactions in an alpine meadow. Soil Biol. Biochem. 2022, 171, 108727. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; O’Hara, R.B. Vegan: Community Ecology Package 2015. Available online: https://cran.r-project.org/web/packages/vegan/vegan.pdf (accessed on 1 September 2019).
- De Caceres, M.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef] [PubMed]
- Metting, B. The systematics and ecology of soil algae. Bot. Rev. 1981, 47, 195–312. [Google Scholar] [CrossRef]
- Fierer, N.; Schimel, J.P.; Holden, P.A. Variations in microbial community composition through two soil depth profiles. Soil Biol. Biochem. 2003, 35, 167–176. [Google Scholar] [CrossRef]
- Schultz, M.; Porembski, S.; Budel, B. Diversity of rock-inhabiting cyanobacterial lichens: Studies on granite inselbergs along the Orinoco and in Guyana. Plant Biol. 2000, 2, 482–495. [Google Scholar] [CrossRef]
- Neustupa, J.; Skaloud, P. Diversity of subaerial algae and cyanobacteria growing on bark and wood in the lowland tropical forests of Singapore. Plant Ecol. Evol. 2010, 143, 51–62. [Google Scholar] [CrossRef]
- Cano-Diaz, C.; Maestre, F.T.; Wang, J.; Li, J.; Singh, B.K.; Ochoa, V.; Gozalo, B.; Delgado-Baquerizo, M. Effects of vegetation on soil cyanobacterial communities through time and space. New Phytol. 2022, 234, 435–448. [Google Scholar] [CrossRef]
- Aleksakhina, T.; Shtina, E. Terrestrial Algae of Forest Biogeocoenoses (Pochvennye vodorosli lesnykh Biogeotsenozov); Nauka: Moscow, Russia, 1984; p. 174. [Google Scholar]
- Hoffmann, L. Algae of terrestrial habitats. Bot. Rev. 1989, 55, 77–105. [Google Scholar] [CrossRef]
- Khaybullina, L.S.; Gaysina, L.A.; Johansen, J.R.; Krautová, M. Examination of the terrestrial algae of the Great Smoky Mountains National Park, USA. Fottea 2010, 10, 201–215. [Google Scholar] [CrossRef]
- Dodds, W.K.; Gudder, D.A.; Mollenhauer, D. The ecology of Nostoc. J. Phycol. 1995, 31, 2–18. [Google Scholar] [CrossRef]
- Yu, S.; Mo, Q.; Chen, Y.; Li, Y.; Li, Y.; Zou, B.; Xia, H.; Jun, W.; Li, Z.; Wang, F. Effects of seasonal precipitation change on soil respiration processes in a seasonally dry tropical forest. Ecol. Evol. 2020, 10, 467–479. [Google Scholar] [CrossRef] [PubMed]
- Sun, R.; Zhang, P.; Riggins, C.W.; Zabaloy, M.C.; Rodriguez-Zas, S.; Villami, M.B. Long-Term N Fertilization Decreased Diversity and Altered the Composition of Soil Bacterial and Archaeal Communities. Agronomy 2019, 9, 574. [Google Scholar] [CrossRef]
- Rejmankova, E.; Komarkova, J. Response of cyanobacterial mats to nutrient and salinity changes. Aquat. Bot. 2005, 83, 87–107. [Google Scholar] [CrossRef]
- Ramirez, K.S.; Lauber, C.L.; Knight, R.; Bradford, M.A.; Fierer, N. Consistent effects of nitrogen fertilization on soil bacterial communities in contrasting systems. Ecology 2010, 91, 3463–3470. [Google Scholar] [CrossRef]
- Ramirez, K.S.; Craine, J.M.; Fierer, N. Consistent effects of nitrogen amendments on soil microbial communities and processes across biomes. Glob. Chang. Biol. 2012, 18, 1918–1927. [Google Scholar] [CrossRef]
- Hall, S.J.; Matson, P.A. Nitrogen oxide emissions after nitrogen additions in tropical forests. Nature 1999, 400, 152–155. [Google Scholar] [CrossRef]
- Zhang, Y.; Shen, H.; He, X.; Thomas, B.W.; Lupwayi, N.Z.; Hao, X.; Thomas, M.C.; Shi, X. Fertilization Shapes Bacterial Community Structure by Alteration of Soil pH. Front. Microbiol. 2017, 8, 1325. [Google Scholar] [CrossRef]
- Chen, D.; Lan, Z.; Hu, S.; Bai, Y. Effects of nitrogen enrichment on belowground communities in grassland: Relative role of soil nitrogen availability vs. soil acidification. Soil Biol. Biochem. 2015, 89, 99–108. [Google Scholar] [CrossRef]
- Reynaud, P.A. Ecology of nitrogen-fixing cyanobacteria in dry tropical habitats of West Africa: A multivariate analysis. Plant Soil 1987, 98, 203–220. [Google Scholar] [CrossRef]
- Sun, Q.; Zheng, Y.; Yu, T.; Wu, Y.; Yang, J.; Wu, Z.; Wu, J.; Li, S. Responses of soil diazotrophic diversity and community composition of nodulating and non-nodulating peanuts (Arachis hypogaea L.) to nitrogen fertilization. Acta Agron. Sin. 2022, 48, 2575–2587. [Google Scholar]
- Duah-Yentumi, S.; Ronn, R.; Christensen, S. Nutrients limiting microbial growth in a tropical forest soil of Ghana under different management. Appl. Soil Ecol. 1998, 8, 19–24. [Google Scholar] [CrossRef]
- Yanni, Y.G.; Abdelrahman, A.A.M. Assessing phosphorus fertilization of rice in the nile delta involving nitrogen and cyanobacteria. Soil Biol. Biochem. 1993, 25, 289–293. [Google Scholar] [CrossRef]
- Roger, P.A.; Kulasooriya, S.A. Biue-Green Algae and Rice; The International Rice Research Institute: Manila, Philippines, 1980; pp. 29–30. [Google Scholar]
- Chapin, D.M.; Bliss, L.C.; Bledsoe, L.J. Environmental-regulation of nitrogen-fixation in a high artic lowland ecosystem. Can. J. Bot.-Rev. Can. Bot. 1991, 69, 2744–2755. [Google Scholar] [CrossRef]
- Benner, J.W.; Vitousek, P.M. Development of a diverse epiphyte community in response to phosphorus fertilization. Ecol. Lett. 2007, 10, 628–636. [Google Scholar] [CrossRef]
- Cerna, B.; Rejmankova, E.; Snyder, J.M.; Santruckova, H. Heterotrophic nitrogen fixation in oligotrophic tropical marshes: Changes after phosphorus addition. Hydrobiologia 2009, 627, 55–65. [Google Scholar] [CrossRef]
- Rejmankova, E.; Macek, P.; Epps, K. Wetland ecosystem changes after three years of phosphorus addition. Wetlands 2008, 28, 914–927. [Google Scholar] [CrossRef]
- Santruckova, H.; Rejmankova, E.; Pivnickova, B.; Snyder, J.M. Nutrient enrichment in tropical wetlands: Shifts from autotrophic to heterotrophic nitrogen fixation. Biogeochemistry 2010, 101, 295–310. [Google Scholar] [CrossRef]
- Verbruggen, E.; van der Heijden, M.G.A.; Weedon, J.T.; Kowalchuk, G.A.; Roling, W.F.M. Community assembly, species richness and nestedness of arbuscular mycorrhizal fungi in agricultural soils. Mol. Ecol. 2012, 21, 2341–2353. [Google Scholar] [CrossRef]
- Hillebrand, H.; Bennett, D.M.; Cadotte, M.W. Consequences of dominance: A review of evenness effects on local and regional ecosystem processes. Ecology 2008, 89, 1510–1520. [Google Scholar] [CrossRef] [PubMed]
- Song, M.-H.; Zong, N.; Jiang, J.; Shi, P.-L.; Zhang, X.-Z.; Gao, J.-Q.; Zhou, H.-K.; Li, Y.-K.; Loreau, M. Nutrient-induced shifts of dominant species reduce ecosystem stability via increases in species synchrony and population variability. Sci. Total Environ. 2019, 692, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Siddique, I.; Guimaraes Vieira, I.C.; Schmidt, S.; Lamb, D.; Reis Carvalho, C.J.; Figueiredo, R.D.O.; Blomberg, S.; Davidson, E.A. Nitrogen and phosphorus additions negatively affect tree species diversity in tropical forest regrowth trajectories. Ecology 2010, 91, 2121–2131. [Google Scholar] [CrossRef] [PubMed]
- Matson, P.A.; McDowell, W.H.; Townsend, A.R.; Vitousek, P.M. The globalization of N deposition: Ecosystem consequences in tropical environments. Biogeochemistry 1999, 46, 67–83. [Google Scholar] [CrossRef]
- Ma, S.; Chen, G.; Tang, W.; Xing, A.; Chen, X.; Xiao, W.; Zhou, L.; Zhu, J.; Li, Y.; Zhu, B.; et al. Inconsistent responses of soil microbial community structure and enzyme activity to nitrogen and phosphorus additions in two tropical forests. Plant Soil 2021, 460, 453–468. [Google Scholar] [CrossRef]
- Turner, B.L.; Wright, S.J. The response of microbial biomass and hydrolytic enzymes to a decade of nitrogen, phosphorus, and potassium addition in a lowland tropical rain forest. Biogeochemistry 2014, 117, 115–130. [Google Scholar] [CrossRef]
- Zhao, M.; Cong, J.; Cheng, J.; Qi, Q.; Sheng, Y.; Ning, D.; Lu, H.; Wyckoff, K.N.; Deng, Y.; Li, D.; et al. Soil Microbial Community Assembly and Interactions Are Constrained by Nitrogen and Phosphorus in Broadleaf Forests of Southern China. Forests 2020, 11, 285. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, C.; Yu, W.W.; Turak, A.; Chen, D.W.; Huang, Y.; Ao, J.H.; Jiang, Y.; Huang, Z.R. Effects of Nitrogen and Phosphorus Inputs on Soil Bacterial Abundance, Diversity, and Community Composition in Chinese Fir Plantations. Front. Microbiol. 2018, 9, 1543. [Google Scholar] [CrossRef]
- Ma, S.; Chen, G.; Tian, D.; Du, E.; Xiao, W.; Jiang, L.; Zhou, Z.; Zhu, J.; He, H.; Zhu, B.; et al. Effects of seven-year nitrogen and phosphorus additions on soil microbial community structures and residues in a tropical forest in Hainan Island, China. Geoderma 2020, 361, 114034. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, F.; Li, J.; Zou, B.; Wang, X.; Li, Z.; Fu, S. Effects of experimental nitrogen and/or phosphorus additions on soil nematode communities in a secondary tropical forest. Soil Biol. Biochem. 2014, 75, 1–10. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Abundance | Species Richness | Shannon’s Diversity | ||||
|---|---|---|---|---|---|---|
| Coefficient | p | Coefficient | p | Coefficient | p | |
| N addition | −6.50 | 0.851 | 2.18 | 0.102 | 0.17 | 0.136 |
| P addition | −78.75 | 0.023 | 3.39 | 0.011 | 0.31 | 0.005 |
| 10–20 cm | 0.44 | 0.900 | −0.44 | 0.001 | −0.04 | <0.001 |
| Soil Property | N Addition | P Addition | 10–20 cm | |
|---|---|---|---|---|
| pH | coefficient | −0.191 | 0.251 | 0.034 |
| p | 0.029 | 0.004 | <0.001 | |
| TC | coefficient | 0.054 | −0.336 | −0.061 |
| p | 0.680 | 0.007 | <0.001 | |
| TN | coefficient | 0.127 | −0.339 | −0.068 |
| p | 0.080 | <0.001 | <0.001 | |
| TP | coefficient | −0.013 | 0.284 | −0.011 |
| p | 0.708 | <0.001 | 0.009 | |
| NH4+-N | coefficient | 0.653 | −0.970 | −0.102 |
| p | 0.114 | 0.018 | 0.016 | |
| NO3−-N | coefficient | 4.708 | −7.412 | 0.047 |
| p | 0.004 | <0.001 | 0.741 | |
| Pextrac | coefficient | −0.802 | 20.601 | −0.065 |
| p | 0.106 | <0.001 | 0.160 |
| Species | Treatment | p |
|---|---|---|
| Richelia sp. SL21 | +NP | 0.041 |
| unclassified gene 1 | +NP | 0.034 |
| Nostoc sp. 3335mG | +N and +NP | 0.027 |
| unclassified gene 2 | +P and +NP | 0.046 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zou, X.; Jiang, X.; Chen, C.; Kuang, C.; Ye, J.; Qin, S.; Cheng, J.; Liu, G.; Wang, F.; Yu, S. Phosphorus Rather than Nitrogen Addition Changed Soil Cyanobacterial Community in a Tropical Secondary Forest of South China. Forests 2023, 14, 2216. https://doi.org/10.3390/f14112216
Zou X, Jiang X, Chen C, Kuang C, Ye J, Qin S, Cheng J, Liu G, Wang F, Yu S. Phosphorus Rather than Nitrogen Addition Changed Soil Cyanobacterial Community in a Tropical Secondary Forest of South China. Forests. 2023; 14(11):2216. https://doi.org/10.3390/f14112216
Chicago/Turabian StyleZou, Xiangbo, Xinyu Jiang, Chuangting Chen, Cao Kuang, Ji Ye, Shiwei Qin, Jiong Cheng, Guangli Liu, Faming Wang, and Shiqin Yu. 2023. "Phosphorus Rather than Nitrogen Addition Changed Soil Cyanobacterial Community in a Tropical Secondary Forest of South China" Forests 14, no. 11: 2216. https://doi.org/10.3390/f14112216
APA StyleZou, X., Jiang, X., Chen, C., Kuang, C., Ye, J., Qin, S., Cheng, J., Liu, G., Wang, F., & Yu, S. (2023). Phosphorus Rather than Nitrogen Addition Changed Soil Cyanobacterial Community in a Tropical Secondary Forest of South China. Forests, 14(11), 2216. https://doi.org/10.3390/f14112216