
The Species Richness-Environment Relationship for Cherries (Prunus subgenus Cerasus) across the Northern Hemisphere

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Species Distribution Data
2.2. Environmental Data
2.3. Statistical Analysis
3. Results
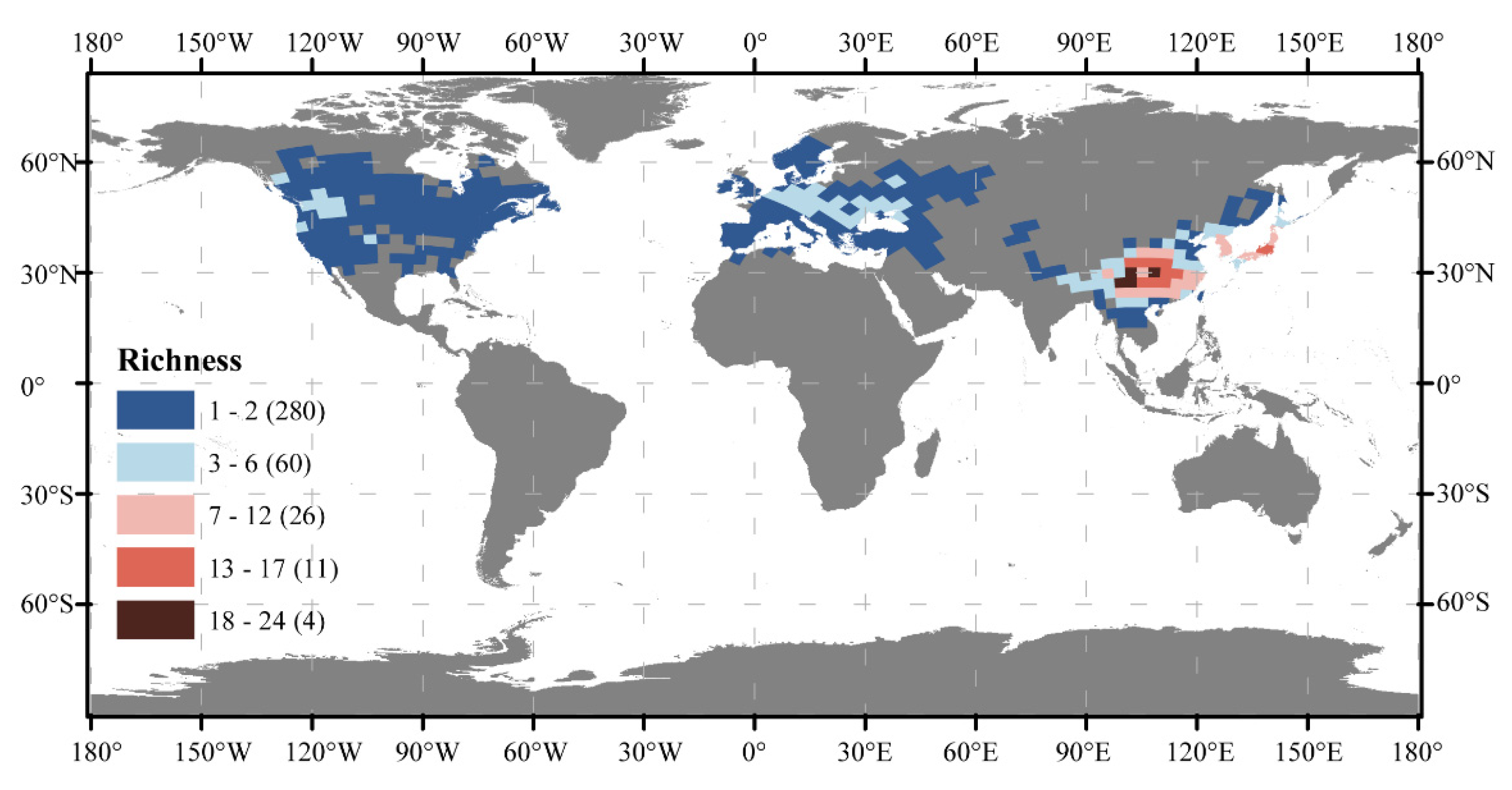
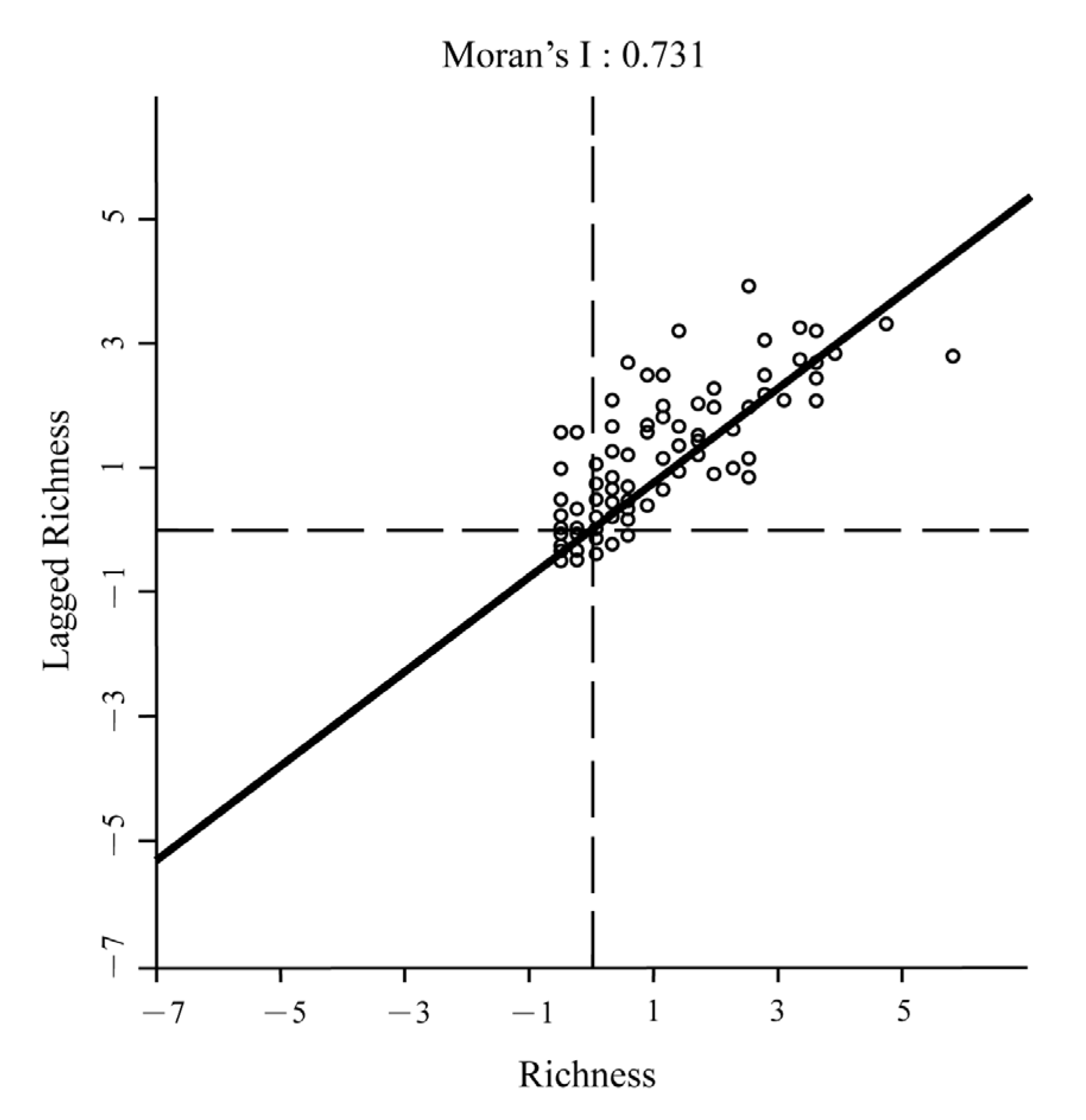
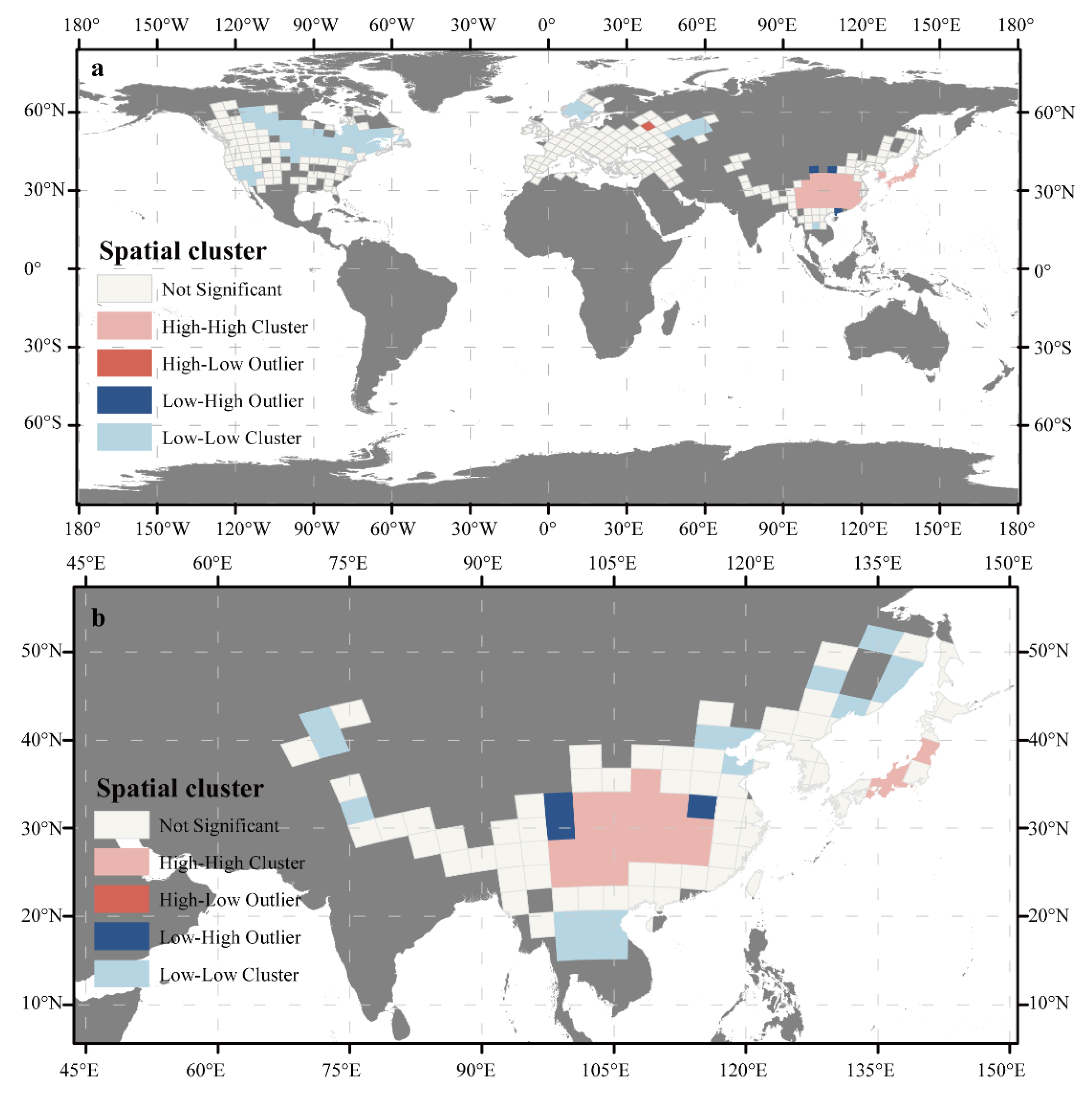
3.1. Global Patterns of Cerasus Species Richness
3.2. Richness–Environment Relationships
4. Discussion
4.1. Supporting Hypothesis
4.2. Geographical Pattern of Cerasus in Eastern Asia
4.3. Geographical Pattern of Cerasus in Europe and North America
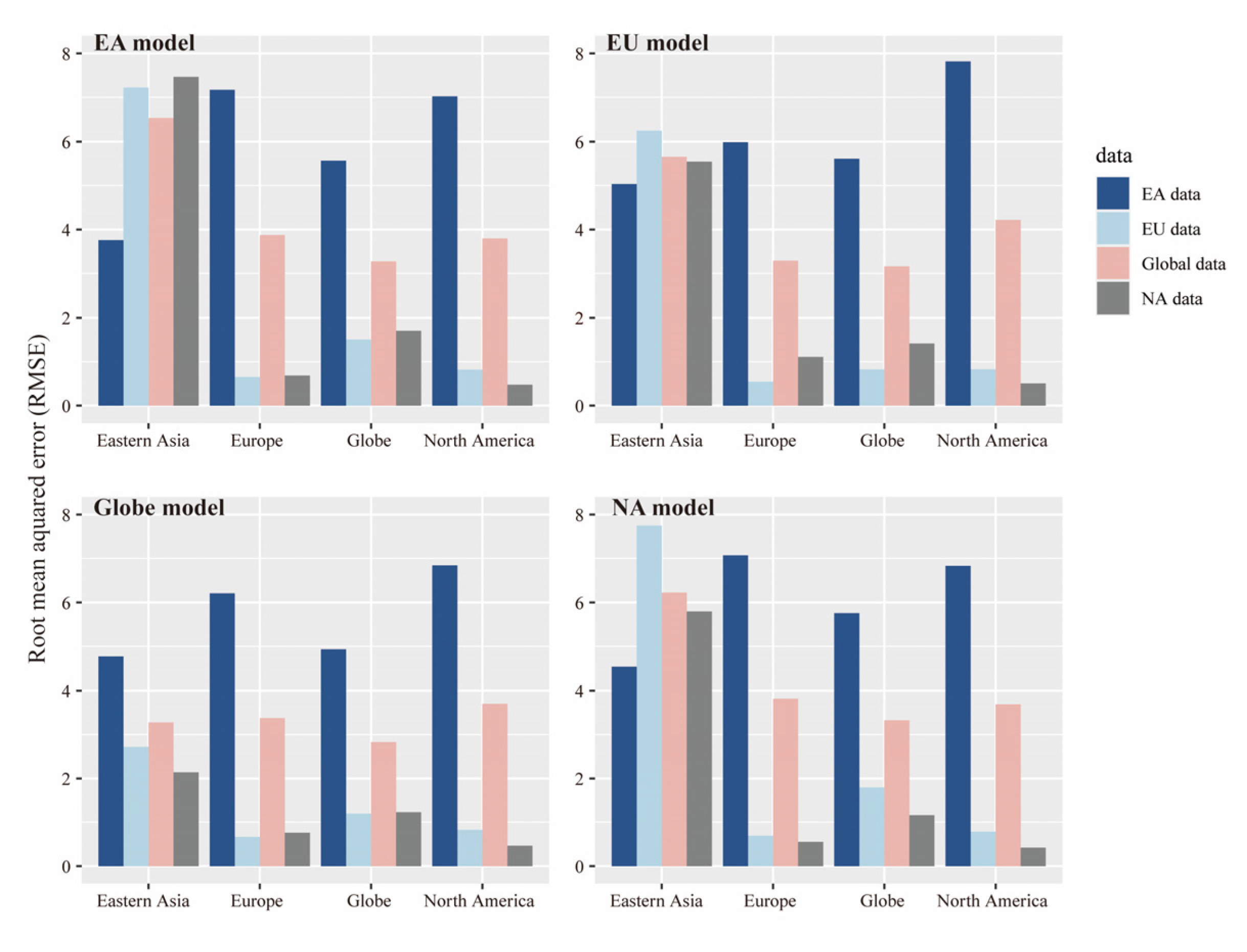
4.4. Differences in Species Richness between North America and East Asia
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- D’Antraccoli, M.; Roma-Marzio, F.; Carta, A.; Landi, S.; Bedini, G.; Chiarucci, A.; Peruzzi, L. Drivers of floristic richness in the Mediterranean: A case study from Tuscany. Biodivers. Conserv. 2019, 28, 1411–1429. [Google Scholar] [CrossRef]
- Gotelli, N.J.; Colwell, R.K. Quantifying biodiversity: Procedures and pitfalls in the measurement and comparison of species richness. Ecol. Lett. 2001, 4, 379–391. [Google Scholar] [CrossRef] [Green Version]
- Grytnes, J.A.; Vetaas, O.R. Species richness and altitude: A comparison between null models and interpolated plant species richness along the Himalayan altitudinal gradient, Nepal. Am. Nat. 2002, 159, 294–304. [Google Scholar] [CrossRef]
- Krogsgaard Svendsen, I. The Effects that the Current Climate Crisis have on the Biogeography and Environment, Needed Adaptations and Conservation. Am. J. BioScience 2020, 8, 20–27. [Google Scholar] [CrossRef]
- Currie, D.J.; Mittelbach, G.G.; Cornell, H.V.; Field, R.; Guegan, J.F.; Hawkins, B.A.; Kaufman, D.M.; Kerr, J.T.; Oberdorff, T.; O’Brien, E.; et al. Predictions and tests of climate-based hypotheses of broad-scale variation in taxonomic richness. Ecol. Lett. 2004, 7, 1121–1134. [Google Scholar] [CrossRef]
- Field, R.; O’Brien, E.M.; Whittaker, R.J. Global models for.predicting woody plant richness from climate: Development and evaluation. Ecology 2005, 86, 2263–2277. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, E.M.; Field, R.; Whittaker, R.J. Climatic gradients in woody plant (tree and shrub) diversity: Water-energy dynamics, residual variation, and topography. Oikos 2000, 89, 588–600. [Google Scholar] [CrossRef]
- Tews, J.; Brose, U.; Grimm, V.; Tielborger, K.; Wichmann, M.C.; Schwager, M.; Jeltsch, F. Animal species diversity driven by habitat heterogeneity/diversity: The importance of keystone structures. J. Biogeogr. 2004, 31, 79–92. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.H.; Fang, J.Y.; Tang, Z.Y.; Lin, X. Patterns, determinants and models of woody plant diversity in China. Proc. R. Soc. B-Biol. Sci. 2011, 278, 2122–2132. [Google Scholar] [CrossRef]
- Hawkins, B.A.; Rodriguez, M.A.; Weller, S.G. Global angiosperm family richness revisited: Linking ecology and evolution to climate. J. Biogeogr. 2011, 38, 1253–1266. [Google Scholar] [CrossRef]
- Hawkins, B.A.; Field, R.; Cornell, H.V.; Currie, D.J.; Guegan, J.F.; Kaufman, D.M.; Kerr, J.T.; Mittelbach, G.G.; Oberdorff, T.; O’Brien, E.M.; et al. Energy, water, and broad-scale geographic patterns of species richness. Ecology 2003, 84, 3105–3117. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.H.; Tang, Z.Y.; Fang, J.Y. The species-energy hypothesis as a mechanism for species richness pattern. Biodivers. Sci. 2009, 17, 613–624. [Google Scholar] [CrossRef]
- Wright, D.H. Species-Energy Theory: An Extension of Species-Area Theory. Oikos 1983, 41, 496–506. [Google Scholar] [CrossRef] [Green Version]
- Jetz, W.; Rahbek, C. Geographic range size and determinants of avian species richness. Science 2002, 297, 1548–1551. [Google Scholar] [CrossRef] [Green Version]
- Kreft, H.; Jetz, W. Global patterns and determinants of vascular plant diversity. Proc. Natl. Acad. Sci. USA 2007, 104, 5925–5930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whittaker, R.J.; Willis, K.J.; Field, R. Scale and species richness: Towards a general, hierarchical theory of species diversity. J. Biogeogr. 2001, 28, 453–470. [Google Scholar] [CrossRef] [Green Version]
- Suissa, J.S.; Sundue, M.A.; Testo, W.L. Mountains, climate and niche heterogeneity explain global patterns of fern diversity. J. Biogeogr. 2021, 48, 1296–1308. [Google Scholar] [CrossRef]
- Fang, W.J.; Cai, Q.; Zhao, Q.; Ji, C.J.; Zhu, J.L.; Tang, Z.Y.; Fang, J.Y. Species richness patterns and the determinants of larch forests in China. Plant Divers. 2022, 44, 436–444. [Google Scholar] [CrossRef]
- Rao, M.D.; Manuel, J.S.; Xiang, X.; Zhang, M.; Mi, X.; Zhang, J.; Ma, K.; Jens-Christian, S. Environmental and evolutionary drivers of diversity patterns in the tea family (Theaceae s.s.) across China. Ecol. Evol. 2018, 8, 11663–11676. [Google Scholar] [CrossRef]
- Xu, X.T.; Dimitrov, D.; Shrestha, N.; Rahbek, C.; Wang, Z.H. A consistent species richness-climate relationship for oaks across the Northern Hemisphere. Glob. Ecol. Biogeogr. 2019, 28, 1051–1066. [Google Scholar] [CrossRef]
- Chin, S.W.; Shaw, J.; Haberle, R.; Wen, J.; Potter, D. Diversification of almonds, peaches, plums and cherries–Molecular systematics and biogeographic history of Prunus (Rosaceae). Mol. Phylogenetics Evol. 2014, 76, 34–48. [Google Scholar] [CrossRef]
- McCune, L.M.; Kubota, C.; Stendell-Hollis, N.R.; Thomson, C.A. Cherries and Health: A Review. Crit. Rev. Food Sci. Nutr. 2010, 51, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Hu, G.P.; Liu, Z.S.; Zhang, J.; Ma, L.; Tian, T.; Wang, H.; Chen, T.; Chen, Q.; He, W.; et al. Phenotyping in flower and main fruit traits of Chinese cherry Cerasus pseudocerasus (Lindl.) G.Don. Sci. Hortic. 2022, 296, 110920. [Google Scholar] [CrossRef]
- Karaat, F.E.; Gunduz, K.; Saracoglu, O.; Yildirim, H. Pomological and phytochemical evaluation of different cherry species: Mahaleb (Prunus mahaleb L.), wild sweet cherry (Prunus avium L.) and wild sour cherry (Prunus cerasus L.), sweet and sour cherry cultivars. Acta Sci. Pol.-Hortorum Cultus 2019, 18, 181–191. [Google Scholar] [CrossRef]
- Wang, Y.C.; Bao, W.K.; Wu, N. Shrub island effects on a high-altitude forest cutover in the eastern Tibetan Plateau. Ann. For. Sci. 2011, 68, 1127–1141. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, A. Cherry removal by seed-dispersing mammals: Mutualism through commensal association with frugivorous birds. Pol. J. Ecol. 2008, 56, 127–138. [Google Scholar]
- Ding, L.; Xu, L.J.; Chu, X.; Yang, L.; Zhu, H.L.; Huang, J.X. Dissimilarity analysis of microbial communities in the rhizosphere and tissues of diseased and healthy cherry trees (Cerasus pseudocerasus). Can. J. Plant Pathol. 2021, 43, 612–621. [Google Scholar] [CrossRef]
- Eken, B.U.; Kirdok, E.; Velioglu, E.; Ciftci, Y.O. Assessment of genetic variation of natural populations of wild cherry (Prunus avium L.) via SSR markers. Turk. J. Bot. 2022, 46, 14–25. [Google Scholar] [CrossRef]
- Hou, Q.D.; Li, S.; Shang, C.Q.; Wen, Z.; Cai, X.W.; Hong, Y.; Qiao, G. Genome-wide characterization of chalcone synthase genes in sweet cherry and functional characterization of CpCHS1 under drought stress. Front. Plant Sci. 2022, 13, 3054. [Google Scholar] [CrossRef]
- Shirasawa, K.; Esumi, T.; Itai, A.; Isobe, S. Cherry Blossom Forecast Based on Transcriptome of Floral Organs Approaching Blooming in the Flowering Cherry (Cerasus x yedoensis) Cultivar ‘Somei-Yoshino’. Front. Plant Sci. 2022, 13, 802203. [Google Scholar] [CrossRef]
- Shirasawa, K.; Esumi, T.; Hirakawa, H.; Tanaka, H.; Itai, A.; Ghelfi, A.; Nagasaki, H.; Isobe, S. Phased genome sequence of an interspecific hybrid flowering cherry, ‘Somei-Yoshino’ (Cerasus x yedoensis). DNA Res. 2019, 26, 379–389. [Google Scholar] [CrossRef] [PubMed]
- Tobler, W. A Computer Movie Simulating Urban Growth in the Detroit Region. Econ. Geogr. 1970, 46, 234–240. [Google Scholar] [CrossRef]
- Clarke, A.; Gaston, K.J. Climate, energy and diversity. Proc. R. Soc. B-Biol. Sci. 2006, 273, 2257–2266. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Song, Y.F.; Sylvester, S.P.; Wang, X.R. Comparative analysis of the complete plastid genomes in Prunus subgenus Cerasus (Rosaceae): Molecular structures and phylogenetic relationships. PLoS ONE 2022, 17, e0266535. [Google Scholar] [CrossRef]
- Hipp, A.L.; Manos, P.S.; González-Rodríguez, A.; Hahn, M.; Kaproth, M.; McVay, J.D.; Avalos, S.V.; Cavender-Bares, J. Sympatric parallel diversification of major oak clades in the Americas and the origins of Mexican species diversity. New Phytol. 2018, 217, 439–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moghadam, E.G.; Khalighi, A. Relationship between vigor of Iranian Prunus mahaleb L. selected dwarf rootstocks and some morphological characters. Sci. Hortic. 2007, 111, 209–212. [Google Scholar] [CrossRef]
- Bujdosó, G.; Hrotkó, K. Cherries: Botany, Production and Uses; Quero-Garcia, J., Iezzoni, A., Pulawska, J., Lang, G., Eds.; Cherry production; CABI International: Wallingford, UK, 2017; pp. 1–13. [Google Scholar]
- Shiell, K.J.; St-Pierre, R.G.; Zatylny, A.M. Timing, magnitude and causes of flower and immature fruit loss in pin cherry and choke cherry. Can. J. Plant Sci. 2002, 82, 157–164. [Google Scholar] [CrossRef] [Green Version]
- St-Pierre, R.G.; Zatylny, A.M.; Tulloch, H.P. Evaluation of growth, yield, and fruit size of chokecherry, pincherry, highbush cranberry, and black currant cultivars in Saskatchewan. Can. J. Plant Sci. 2005, 85, 659–664. [Google Scholar] [CrossRef]
- Lurie, S.; Crisosto, C.H. Chilling injury in peach and nectarine. Postharvest Biol. Technol. 2005, 37, 195–208. [Google Scholar] [CrossRef]
- Qian, H. A comparison of the taxonomic richness of temperate plants in East Asia and North America. Am. J. Bot. 2002, 89, 1818–1825. [Google Scholar] [CrossRef]
- Qian, H. A comparison of generic endemism of vascular plants between East Asia and North America. Int. J. Plant Sci. 2001, 162, 191–199. [Google Scholar] [CrossRef]
- Qian, H.; Jin, Y.; Ricklefs, R.E. Phylogenetic diversity anomaly in angiosperms between eastern Asia and eastern North America. Proc. Natl. Acad. Sci. USA 2017, 114, 11452–11457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, X.; Qian, H.; Sui, X.H.; Zhang, M.H.; Mao, L.F.; Svenning, J.C.; Ricklefs, R.E.; He, F.L.; Hurlbert, A. Effects of climate and topography on the diversity anomaly of plants disjunctly distributed in eastern Asia and eastern North America. Glob. Ecol. Biogeogr. 2021, 30, 2029–2042. [Google Scholar] [CrossRef]
- Zuo, Y.J.; Wen, J.; Zhou, S.L. Intercontinental and intracontinental biogeography of the eastern Asian-Eastern North American disjunct Panax (the ginseng genus, Araliaceae), emphasizing its diversification processes in eastern Asia. Mol. Phylogenetics Evol. 2017, 117, 60–74. [Google Scholar] [CrossRef]
- Molnar, P.; Boos, W.R.; Battisti, D.S. Orographic Controls on Climate and Paleoclimate of Asia: Thermal and Mechanical Roles for the Tibetan Plateau. Annu. Rev. Earth Planet. Sci. 2010, 38, 77–102. [Google Scholar] [CrossRef] [Green Version]
- Xing, Y.W.; Ree, R.H. Uplift-driven diversification in the Hengduan Mountains, a temperate biodiversity hotspot. Proc. Natl. Acad. Sci. USA 2017, 114, E3444–E3451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, J.; Zhang, J.Q.; Nie, Z.L.; Zhong, Y.; Sun, H. Evolutionary diversificatons of plants on the Qinghai-Tibetan Plateau. Front. Genet. 2014, 5, 4. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.H.; Ye, J.F.; Liu, B.; Mao, L.F.; Smith, S.A.; Barrett, R.L.; Soltis, P.S.; Soltis, D.E.; Chen, Z.D.; Lu, L.M. Temporal and spatial comparisons of angiosperm diversity between eastern Asia and North America. Natl. Sci. Rev. 2022, 9, nwab199. [Google Scholar] [CrossRef] [PubMed]
- Norris, J.R.; Betancourt, J.L.; Jackson, S.T. Late Holocene expansion of ponderosa pine (Pinus ponderosa) in the Central Rocky Mountains, USA. J. Biogeogr. 2016, 43, 778–790. [Google Scholar] [CrossRef] [Green Version]
- Zinck, J.W.; Rajora, O.P. Post-glacial phylogeography and evolution of a wide-ranging highly-exploited keystone forest tree, eastern white pine (Pinus strobus) in North America: Single refugium, multiple routes. BMC Evol. Biol. 2016, 16, 56. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Abbreviations | Environmental Variables |
|---|---|---|
| Environmental energy availability | MAT | Mean annual temperature (°C) |
| MTWQ | Mean temperature of the coldest quarter (°C) | |
| MTCQ | Mean temperature of the warmest quarter (°C) | |
| PET | Potential evapotranspiration (mm) | |
| PETmin | Minimum monthly potential evapotranspiration (mm) | |
| Water availability | MAP | Mean annual precipitation (mm) |
| AET | Actual evapotranspiration (mm) | |
| WD | Water deficit (mm) | |
| PWQ | Precipitation of warmest quarter (mm) | |
| PCQ | Precipitation of coldest quarter (mm) | |
| Climate seasonality | ART | Annual range of temperature (°C) |
| TSN | Temperature seasonality (standard deviation of monthly mean temperatures × 100) | |
| PSN | Precipitation seasonality (standard deviation of monthly precipitation) | |
| Habitat heterogeneity | RELE | Range of elevation (m) within each grid cell |
| RMAT | Range of mean annual temperature (°C) within each grid cell | |
| RMAP | Range of mean annual precipitation (mm) within each grid cell | |
| Soil properties | AWC | Available water storage capacity |
| RBD | Reference bulk density (kg/dm3) | |
| USDA | The relative proportion of different grain sizes of mineral particles in soil | |
| OC | Organic carbon (% weight) | |
| CEC | Cation exchange capacity (cmol/kg) | |
| TEB | Total exchange bases (cmol/kg) | |
| pH | Hydrogen ion concentration | |
| ECE | Electrical conductivity (dS/m) |
| Min | Max | Mean | Median | SD | Optimal Range | ||
|---|---|---|---|---|---|---|---|
| Lower Limit | Upper Limit | ||||||
| Richness | 1 | 24 | 2.82 | 1 | 3.61 | ||
| MAT/°C | −7.06 | 26.30 | 8.58 | 8.35 | 6.82 | 6.13 | 15.14 |
| MTWQ/°C | 6.81 | 33.16 | 19.14 | 18.87 | 4.91 | 13.47 | 24.41 |
| MTCQ/°C | −33.82 | 15.78 | −8.56 | −7.44 | 10.55 | −9.85 | 1.17 |
| PET/mm | 0.89 | 2722.32 | 1148.62 | 1106.17 | 400.82 | 950.81 | 1211.56 |
| PETmin/mm | 0.01 | 8.35 | 34.30 | 26.89 | 27.21 | 38.73 | 74.71 |
| MAP/mm | 167.47 | 2540.83 | 848.70 | 698.83 | 456.25 | 869.59 | 1307.80 |
| AET/mm | 250.55 | 3649.93 | 934.94 | 792.18 | 524.55 | 634.04 | 811.49 |
| PWQ/mm | 0.49 | 1435.15 | 285.58 | 240.03 | 210.53 | 483.20 | 571.81 |
| PCQ/mm | 5.85 | 987.23 | 149.85 | 116.88 | 124.45 | 23.09 | 72.85 |
| ART/°C | 15.95 | 57.69 | 34.63 | 34.29 | 8.67 | 24.13 | 28.64 |
| TSN | 202.56 | 1658.14 | 881.07 | 851.74 | 287.92 | 511.73 | 771.02 |
| PSN | 11.83 | 127.63 | 50.22 | 46.02 | 26.48 | 62.79 | 91.69 |
| RELE/m | 86.00 | 8614.00 | 1982.30 | 1737.00 | 1562.75 | 2630.00 | 6691.00 |
| RMAT/°C | 1.75 | 44.98 | 11.22 | 9.88 | 7.92 | 12.99 | 32.08 |
| RMAP/mm | 99.00 | 8490.00 | 881.63 | 566.00 | 954.36 | 434.00 | 3093.00 |
| Globe | Eastern Asia | Europe | North America | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Group | Variables | Coefficients | adj. R2/% | Variables | Coefficients | adj. R2/% | Variables | Coefficients | adj. R2/% | Variables | Coefficients | adj. R2/% |
| Energy Availability | PET | −0.5501 *** | 15.06 | MTCQ | 0.5791 *** | 20.66 | MAT | 0.9606 *** | 17.2 | MAT | −1.561 *** | 33.44 |
| PETmin | 0.6606 *** | PET | −0.647 *** | PETmin | −0.9306 *** | MTCQ | 1.739 *** | |||||
| Water Availability | PWQ | 0.4176 *** | 18.07 | PCQ | 0.2055 * | 3.33 | MAP | 0.4799 ** | 8.303 | PWQ | −0.3228 *** | 19.31 |
| PCQ | −0.06357 | PCQ | −0.5705 *** | PCQ | 0.3884 *** | |||||||
| Climate Seasonality | ART | −0.2589 *** | 12.6 | PSN | −0.3501 *** | 11.44 | TSN | −0.1827 * | 11.53 | TSN | −0.4314 *** | 18.04 |
| PSN | 0.2524 *** | PSN | −0.3031 *** | |||||||||
| Habitat heterogeneity | RELE | 0.3942 *** | 11.82 | RELE | 0.3432 ** | 13.82 | RMAT | 0.02042 | 2.422 | RELE | 0.3614 *** | 34.95 |
| RMAP | −0.106 | RMAP | −0.2756 ** | RMAP | 0.3051 *** | |||||||
| Soil Properties | AWC | −0.2775 *** | 9.51 | AWC | −0.4068 *** | 17.2 | ECE | −0.2647 ** | 10.56 | CEC | 1.43 ** | 8.575 |
| RBD | −0.3013 *** | ECE | −0.3796 *** | pH | 0.2899 ** | OC | −1.226 *** | |||||
| TEB | −0.5197 * | |||||||||||
| Integrated Model | MAT | 0.4973 *** | 41.92 | PSN | −0.4908 *** | 49.09 | MAT | 0.9697 *** | 41.57 | TSN | −0.3282 *** | 46.74 |
| PET | −0.4985 *** | RELE | 0.5464 *** | PETmin | −0.3074 | RELE | 0.5731 *** | |||||
| PWQ | 0.3547 *** | RMAP | −0.2916 *** | PCQ | −0.4467 *** | PETmin2 | −0.3609 *** | |||||
| RELE | 0.6054 *** | AWC | −0.435 *** | PWQ | 0.6929 *** | ECE | −0.2127 *** | |||||
| RMAP | −0.4847 *** | |||||||||||
| Northeast of Eastern Asia | Southwest of Eastern Asia | |||||
|---|---|---|---|---|---|---|
| Group | Variables | Coefficients | adj. R2/% | Variables | Coefficients | adj. R2/% |
| Energy Availability | MTCQ | 0.7711 *** | 58.11 | MAT | 0.4706 *** | 45.75 |
| PET | −0.933 *** | |||||
| Water Availability | AET | 0.5359 *** | 61.23 | AET | −1.393 *** | 41.94 |
| PCQ | 0.3455 * | WD | −1.211 *** | |||
| Climate Seasonality | TSN | −0.7013 *** | 47.48 | PSN | −0.4478 *** | 18.98 |
| Habitat heterogeneity | RELE | 0.2682 * | 66.93 | RELE | 1.727 *** | 19.06 |
| RMAP | 0.6717 *** | RMAT | −1.621 *** | |||
| Soil Properties | AWC | −0.5726 *** | 50.26 | CEC | 0.7867 *** | 20.99 |
| TEB | −0.8255 *** | USDA | −0.7061 *** | |||
| Integrated Model | RELE | 0.4514 *** | 84.33 | AET | −0.5839 *** | 62.81 |
| MAT | 0.6081 *** | PSN | −0.6329 *** | |||
| PCQ | 0.2175 * | RELE | 0.5193 *** | |||
| RMAP | −0.2326 ** | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, C.-L.; Xie, C.-P.; Idrees, M.; Yi, X.-G.; Wang, X.-R.; Li, M. The Species Richness-Environment Relationship for Cherries (Prunus subgenus Cerasus) across the Northern Hemisphere. Forests 2023, 14, 193. https://doi.org/10.3390/f14020193
Fu C-L, Xie C-P, Idrees M, Yi X-G, Wang X-R, Li M. The Species Richness-Environment Relationship for Cherries (Prunus subgenus Cerasus) across the Northern Hemisphere. Forests. 2023; 14(2):193. https://doi.org/10.3390/f14020193
Chicago/Turabian StyleFu, Chen-Long, Chun-Ping Xie, Muhammad Idrees, Xian-Gui Yi, Xian-Rong Wang, and Meng Li. 2023. "The Species Richness-Environment Relationship for Cherries (Prunus subgenus Cerasus) across the Northern Hemisphere" Forests 14, no. 2: 193. https://doi.org/10.3390/f14020193
APA StyleFu, C. -L., Xie, C. -P., Idrees, M., Yi, X. -G., Wang, X. -R., & Li, M. (2023). The Species Richness-Environment Relationship for Cherries (Prunus subgenus Cerasus) across the Northern Hemisphere. Forests, 14(2), 193. https://doi.org/10.3390/f14020193