
Climate-Adapted Potential Vegetation—A European Multiclass Model Estimating the Future Potential of Natural Vegetation


Abstract
:1. Introduction
2. Materials and Methods
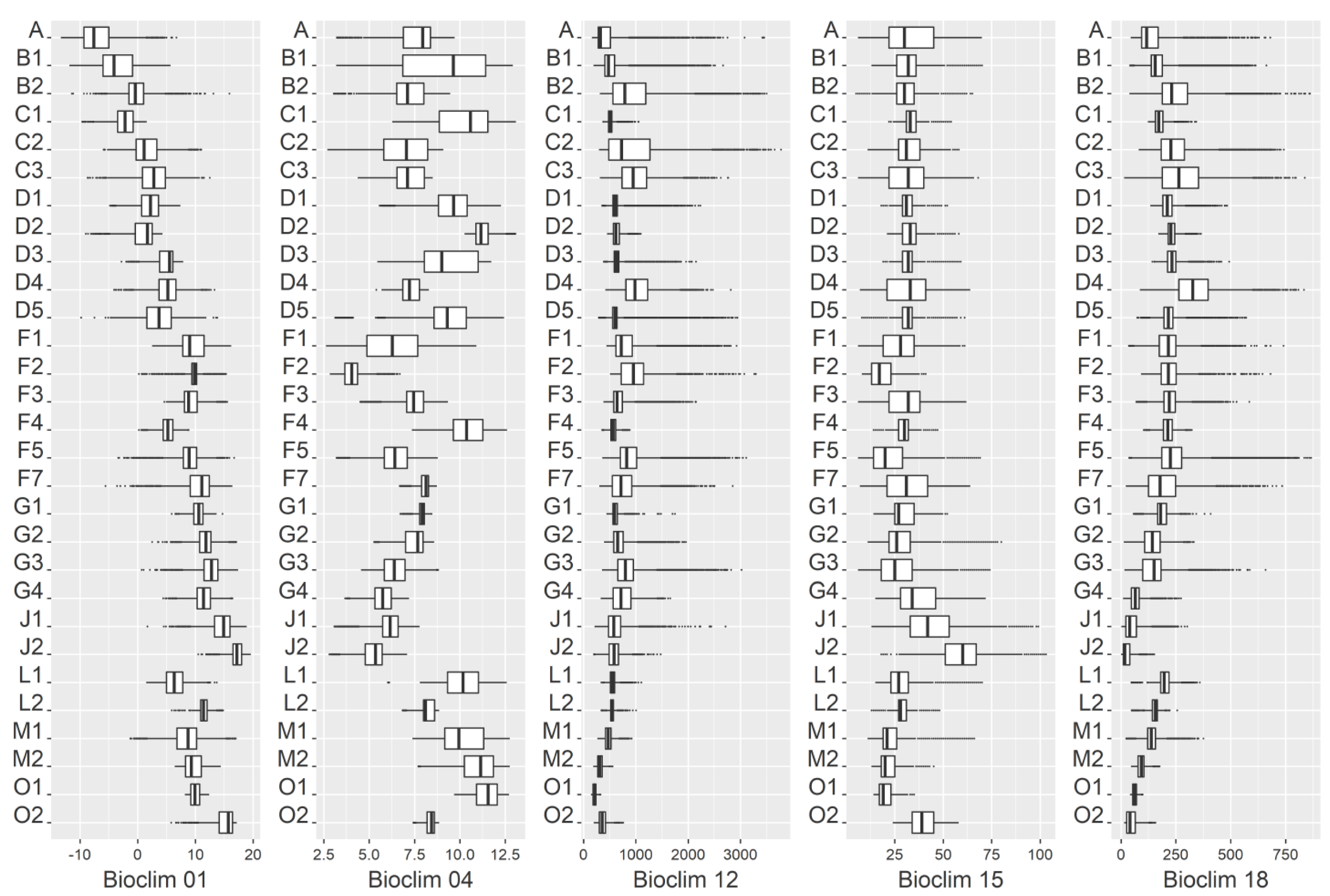
2.1. Data and Data Preparation
2.2. Modeling
3. Results
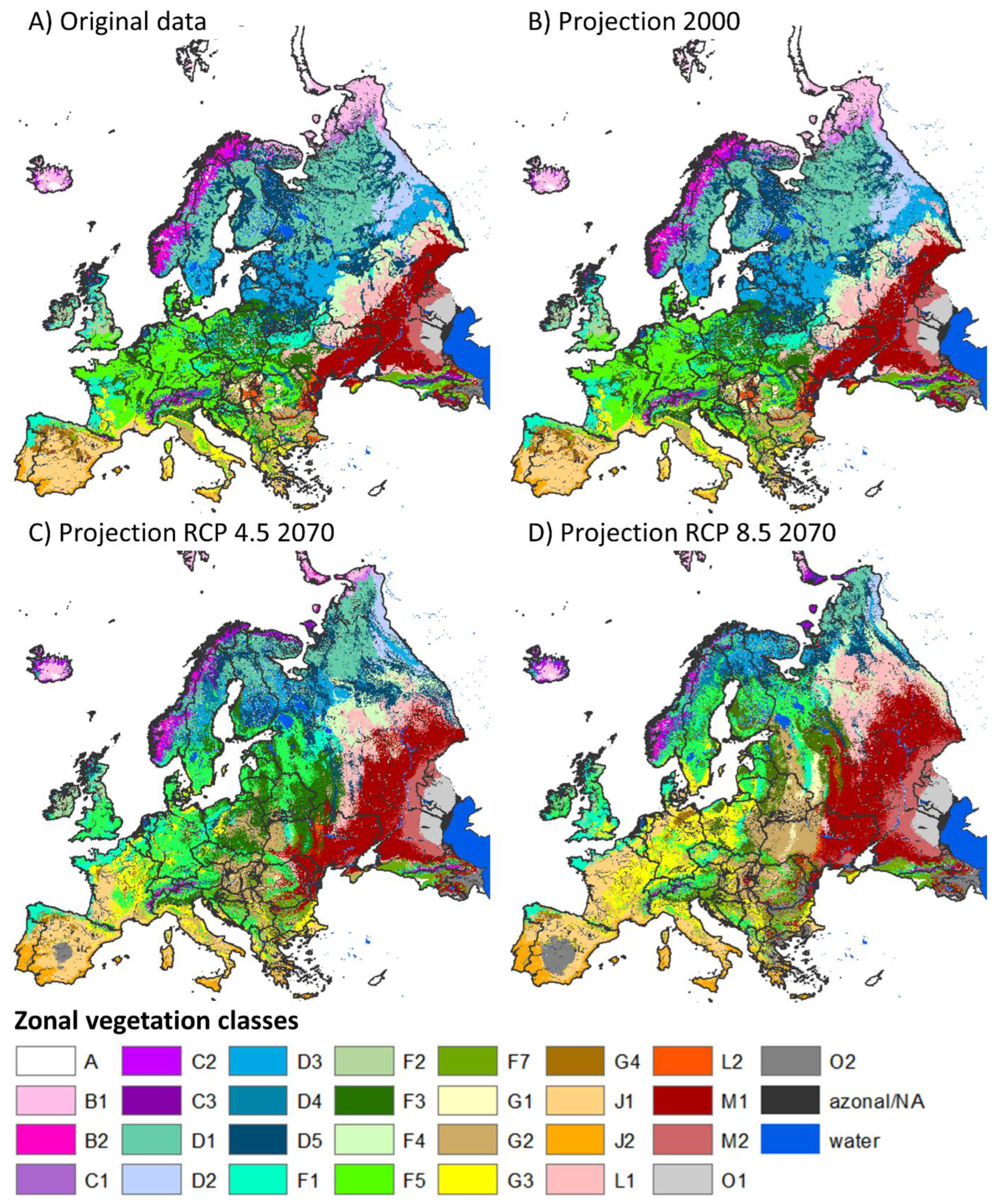
3.1. Model-Based Representation of the Current PNV Map
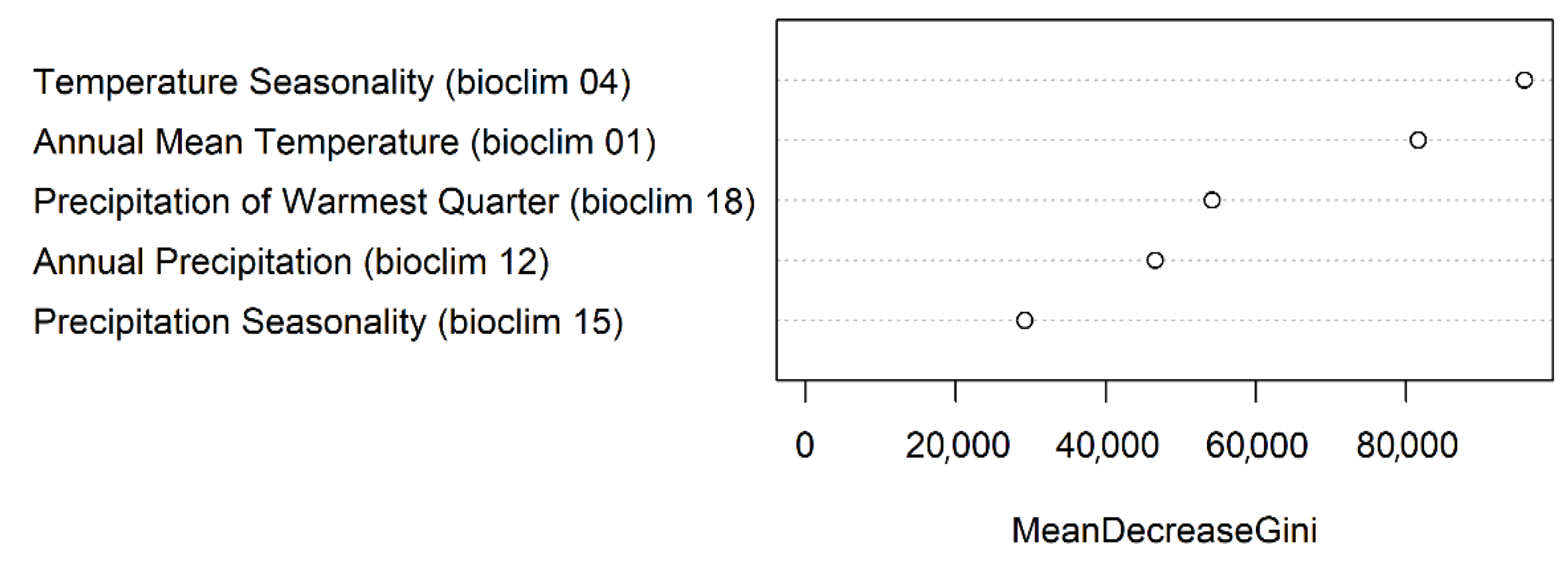
3.2. Variable Importance
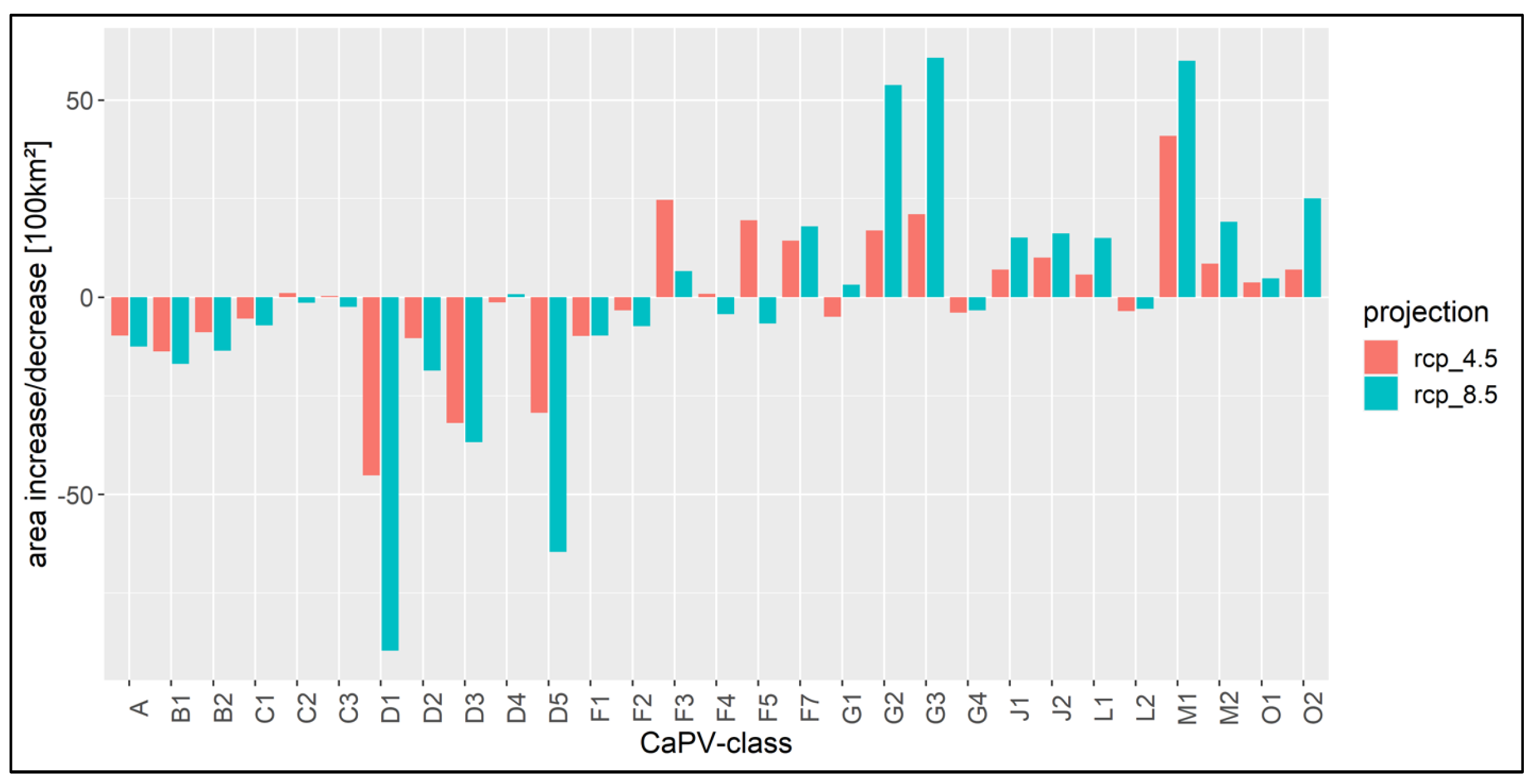
3.3. Model Projections of the Climate-Adapted Potential Vegetation (CaPV)
3.3.1. Vegetation Shifts
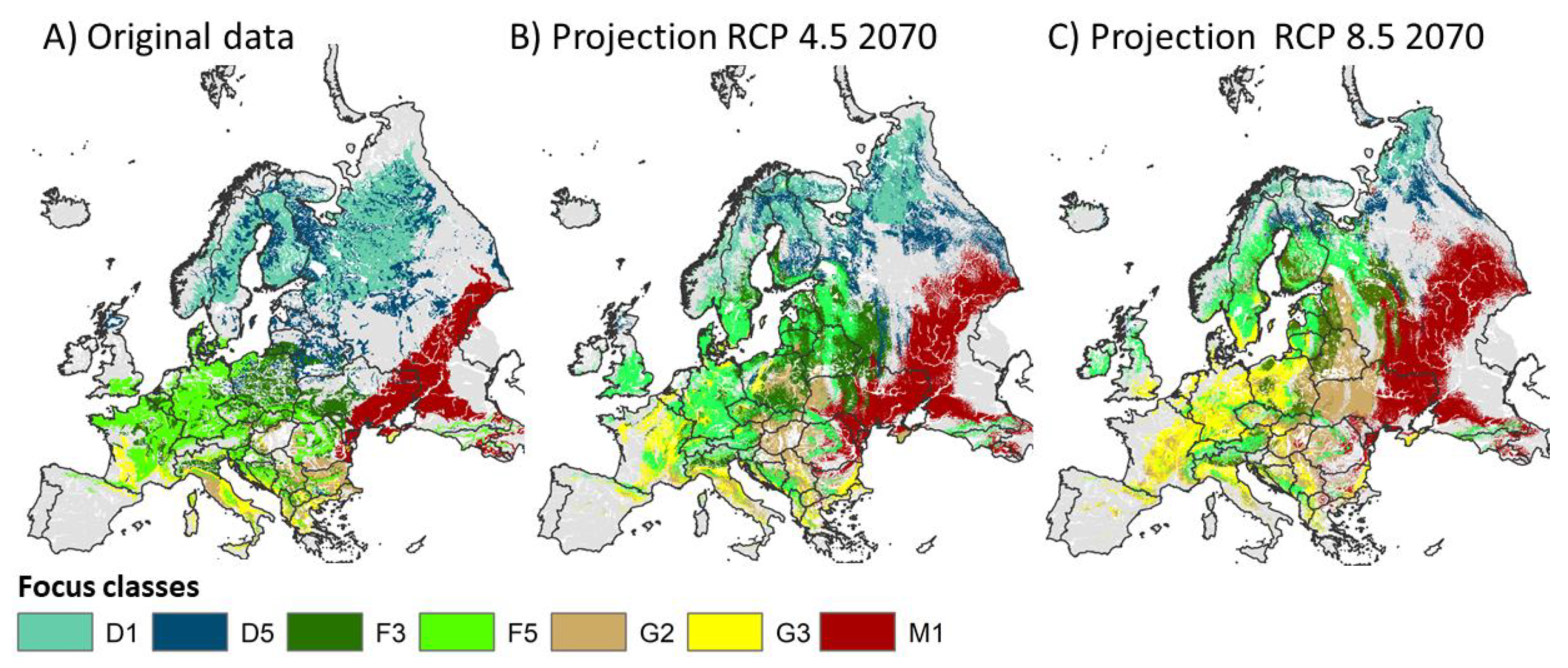
3.3.2. Focus Maps
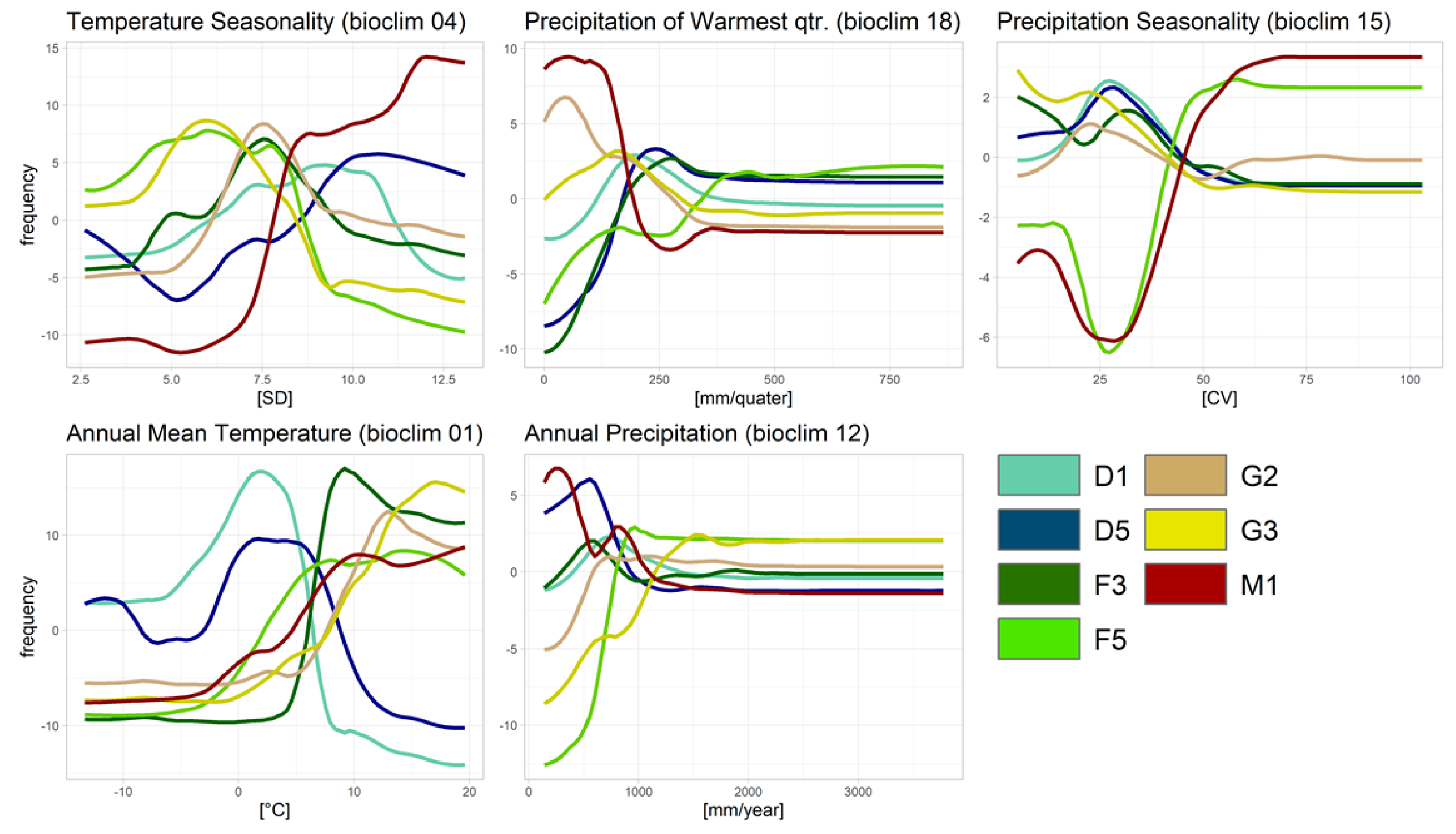
3.3.3. Partial Dependence Plots
4. Discussion
4.1. Shifts in Vegetation Potentials
4.2. Considerations for Biodiversity and Conservation
4.3. Management Considerations
4.4. Methodological Considerations
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
- A.1 = Arctic polar deserts;
- A.2 = Subnival-nival vegetation of high mountains in the boreal and nemoral zone;
- B.1 = Arctic tundras;
- B.2 = Alpine vegetation (Alpine grasslands, low creeping shrub, dwarf shrub and shrub vegetation) in the boreal, nemoral and Mediterranean zone;
- C.1 = Eastern boreal open woodlands (Betula pubescens subsp. czerepanovii, Picea obovate, Pinus sylvestris);
- C.2 = Western boreal and nemoral-montane birch forests (Betula pubescens s. l.), partly with pine forests (Pinus sylvestris);
- C.3 = Subalpine and oro-Mediterranean vegetation (forests, shrub and dwarf shrub communities in combination with grasslands and tall-forb communities);
- D.1 = Western boreal spruce forests (Picea abies, P. obovate, P. abies x P. obovate) partly with Pinus sylvestris, locally with birch (Betula pubescens s. l., B. pendula), alder (Alnus incana) or mixed forests;
- D.2 = Eastern boreal pine-spruce (Picea obovate, Pinus sibirica) and fir-spruce forests (Picea obovate, Abies sibirica), partly with Betula pubescens subsp. czerepanovii, Larix sibirica;
- D.3 = Hemiboreal spruce (Picea abies, P. abies, P. obovate, P. obovate) and fir-spruce forests (Picea obovate, P. abies x P. obovate, Abies sibirica) with broad-leaved trees (Quercus robur, Tilia condata, Ulmus glabra; Acer platanoides, etc.);
- D.4 = Montane to altimontane, partly submontane fir (Abies alba, A. nordmannia) and spruce forests (Picea abies, P. omorika, P. orientalis) in the nemoral zone;
- D.5 = Boreal and hemiboreal pine forests (Pinus sylvestris), partly with Betula pubescens s. l., Picea obovara, P. abies;
- D.6 = Montane to altimontane (subalpine) pine forests (Pinus peuce, P. sylvestris, P. kochiana) in the nemoral zone;
- E = Atlantic dwarf shrub heaths;
- F.1 = Species-poor acidophilous oak and mixed oak forests (Quercus robur, Q. petraea, Q. pyranaica, Pinus sylvestris, Betula pendula, B. pubescens, B. pubescens subsp. Celtiberica, Castanea sativa);
- F.2 = Mixed-oak–ash forests (Fraxinus excelsior, Quercus robur, Ulmus glabra, Quercus petraea);
- F.3 = Mixed-oak–hornbeam forests (Carpinus betulus, Quercus robur, Q. petraea, Tilia cordata);
- F.4 = Lime–pedunculate oak forests (Quercus robur, Tilia cordata, partly Acer platanoides, A. campestre, Ulmus glabra);
- F.5 = Beech and mixed beech forests (Fagus sylvatica, partly F. sylvatica subs. Moesiaca, Abies alba);
- F.6 = Oriental beech forests and hornbeam- Oriental beech forests (Fagus sylvatica subsp. Orientalis, Carpinus betulus);
- F.7 = Caucasian mixed hornbeam-oak forests (Quercus robur, Q. petraea, Q. iberica, Q. pedunculiflora, Q. macranthera, Carpinus betulus, C. orientalis, etc.);
- G.1 = Subcontinental themophilous (mixed) pedunculate oak and sessile oak forests (Quercus robur, Q. petraea, Q. dalechampii, Q. polycarpa, Pinus sylvestris, Acer tataricum);
- G.2 = Sub-Mediterranean-subcontinental themophilous bitter oak and Balkan oak forests (Quercus cerris, Q. petraea, Q. frainetto, Q. dalechampii, Q. pedunculiflora, Q. pubescens, Q. virgiliana, Q. polycarpa, Q. hartwissiana, Carpinus orientalis, Fraxinus ornus);
- G.3 = Sub-Mediterranean and meso-supra-Mediterranean downy oak forests, as well as mixed forests (Quercus pubescens, Q. virgiliana, Q. trojana, Fraxinus ornus, Ostrya carpinifolia, Carpinus orientalis);
- G.4 = Iberian supra- and meso-Mediterranean Quercus pyrenaica, Q. faginea, Q. faginea subsp. broteroi and Q. canariensis forests;
- Gla = Glaceirs;
- H = Hygro-thermophilous mixed deciduous broad-leaved trees;
- J.1 = Meso- and supra-Mediterranean, as well as relict sclerophyllous forests (Quercus ilex, Q. ilex subsp. Rotundifolia, Q. coccifera, Q. suber, Pistacia lentiscus);
- J.2 = Thermo-Mediterranean sclerophyllous forests and xerophytic scrub (Quercus suber, Q. ilex subsp. Rotundifolia, Olea europaea, Ceratonia silique, Periploca angustifolia, Rhamnus lycioides);
- K.1 = Pine forests and pine woodlands (Pinus sylvestris, P. nigra agg., P. heldreichii, P. halepensis, P. brutia, P. pityusa);
- K.2 = Meso- and supra-Mediterranean fir forests (Abies pinsapo, A. cephalpnica);
- K.3 = Juniper and cypress open woodlands and scrub (Juniperus thurifera, J. excelsa, J. foetidissima, J. polycarpos, Cupressus sempervirens);
- L.1 = Subcontinental meadow steppes and steppe-like dry grassland (Festuca rupicola, F. valesciaca, Stipa tirsa, S. pennata, Poa aangustifolia, Agrostis vinealis) alternating with pendunculate oak forests (Quercus robur);
- L.2 = Sub-Mediterranean-subcontinental herb-grass steppes, partly meadow steppes (Festuca valesciaca, Stipa spp., Bothriochola ischaemum, Chrysopogon gryllus) alternating with oak forests (Quercus pubescens, Q. robur, Q. pendunculiflora) with Acer tataricum
- M.1 = True steppes (Stipa pennata, S. trisa, S. dasyphylla, S. ucrainica, Festuca valesiaca, Koeleria macrantha);
- M.2 = Desert steppes (Stipa lessingiana, S. sareptana, Festuca valesiaca, Artemisia spp.)
- N = Oroxerophytic vegetation (thorn-cushion communities, tomillares, mountain steppes, partly scrub);
- O.1 = Northern lowland dwarf semishrub deserts;
- O.2 = Southern lowland-colline dwarf semishrub deserts with ephemeroids.
Appendix B

References
- Hannah, L.; Midgley, G.; Lovejoy, T.; Bond, W.; Bush, M.; Lovett, J.; Scott, D.; Woodward, F. Conservation of biodiversity in a changing climate. Conserv. Biol. 2002, 16, 264–268. [Google Scholar]
- Hengl, T.; Walsh, M.G.; Sanderman, J.; Wheeler, I.; Harrison, S.P.; Prentice, I.C. Global mapping of potential natural vegetation: An assessment of machine learning algorithms for estimating land potential. PeerJ 2018, 6, e5457. [Google Scholar]
- Tüxen, R. Die heutige potentielle natürliche Vegetation als Gegenstand der Vegetationskartierung. Angewandte Pflanzensoziologie 1956, 13, 4–42. [Google Scholar]
- Chiarucci, A.; Araújo, M.B.; Decocq, G.; Beierkuhnlein, C.; Fernández-Palacios, J.M. The concept of potential natural vegetation: An epitaph? J. Veg. Sci. 2010, 21, 1172–1178. [Google Scholar]
- Brang, P.; Spathelf, P.; Larsen, J.B.; Bauhus, J.; Boncčìna, A.; Chauvin, C.; Drössler, L.; García-Güemes, C.; Heiri, C.; Kerr, G. Suitability of close-to-nature silviculture for adapting temperate European forests to climate change. For. Int. J. For. Res. 2014, 87, 492–503. [Google Scholar] [CrossRef] [Green Version]
- Reif, A.; Walentowski, H. The assessment of naturalness and its role for nature conservation and forestry in Europe. Wald. Landsch. Und Nat. 2008, 6, 63–76. [Google Scholar]
- Spielmann, M.; Bücking, W.; Quadt, V.; Krumm, F. Integration of Nature Protection in Forest Policy in Baden-Württemberg (Germany); Integzrate Country Report; EFICENT-OEF: Freiburg, Germany, 2013. [Google Scholar]
- Loidi, J.; Fernández-González, F. Potential natural vegetation: Reburying or reboring? J. Veg. Sci. 2012, 23, 596–604. [Google Scholar] [CrossRef]
- Prach, K.; Tichý, L.; Lencová, K.; Adámek, M.; Koutecký, T.; Sádlo, J.; Bartošová, A.; Novák, J.; Kovář, P.; Jírová, A. Does succession run towards potential natural vegetation? An analysis across seres. J. Veg. Sci. 2016, 27, 515–523. [Google Scholar]
- Bohn, U.; Gollub, G.; Hettwer, C.; Neuhäuslová, Z.; Raus, T.; Schlüter, H.; Weber, H.; Hennekens, S. Map of the natural vegetation of Europe. Scale 2000, 1, 500. [Google Scholar]
- Hickler, T.; Vohland, K.; Feehan, J.; Miller, P.A.; Smith, B.; Costa, L.; Giesecke, T.; Fronzek, S.; Carter, T.R.; Cramer, W. Projecting the future distribution of European potential natural vegetation zones with a generalized, tree species-based dynamic vegetation model. Glob. Ecol. Biogeogr. 2012, 21, 50–63. [Google Scholar]
- Bakkenes, M.; Alkemade, J.; Ihle, F.; Leemans, R.; Latour, J. Assessing effects of forecasted climate change on the diversity and distribution of European higher plants for 2050. Glob. Chang. Biol. 2002, 8, 390–407. [Google Scholar]
- Gonzalez, P.; Neilson, R.P.; Lenihan, J.M.; Drapek, R.J. Global patterns in the vulnerability of ecosystems to vegetation shifts due to climate change. Glob. Ecol. Biogeogr. 2010, 19, 755–768. [Google Scholar] [CrossRef]
- Simpson, M.G. Plant Systematics; Academic Press: Cambridge, MA, USA, 2019. [Google Scholar]
- Ellenberg, H. Tentative physiognomic-ecological classification of plant formations of the earth. Ber. geobot. Inst. ETH Stiftg. 1967, 37, 21–55. [Google Scholar]
- Karger, D.N.; Conrad, O.; Böhner, J.; Kawohl, T.; Kreft, H.; Wilber Soria-Auza, R.; Zimmermann, N.; Linder, H.P.; Kessler, M. Climatologies at high resolution for the earth’s land surface areas. arXiv 2016, arXiv:1607.00217. [Google Scholar]
- Taylor, K.E.; Stouffer, R.J.; Meehl, G.A. An overview of CMIP5 and the experiment design. Bull. Am. Meteorol. Soc. 2012, 93, 485–498. [Google Scholar] [CrossRef] [Green Version]
- Blázquez, J.; Nunez, M.N. Analysis of uncertainties in future climate projections for South America: Comparison of WCRP-CMIP3 and WCRP-CMIP5 models. Clim. Dyn. 2013, 41, 1039–1056. [Google Scholar]
- Lutz, A.F.; ter Maat, H.W.; Biemans, H.; Shrestha, A.B.; Wester, P.; Immerzeel, W.W. Selecting representative climate models for climate change impact studies: An advanced envelope-based selection approach. Int. J. Climatol. 2016, 36, 3988–4005. [Google Scholar]
- Guisan, A.; Thuiller, W.; Zimmermann, N.E. Habitat Suitability and Distribution Models: With Applications in R; Cambridge University Press: Cambridge, MA, USA, 2017. [Google Scholar]
- Christensen, O.; Christensen, J. Intensification of extreme European summer precipitation in a warmer climate. Glob. Planet. Chang. 2004, 44, 107–117. [Google Scholar]
- RStudio-Team. RStudio: Integrated Development Environment for R; RStudio, PBC: Boston, MA, USA, 2020. [Google Scholar]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar]
- Gall, J.; Razavi, N.; Van Gool, L. An introduction to random forests for multi-class object detection. In Outdoor and Large-Scale Real-World Scene Analysis; Springer: Berlin/Heidelberg, Germany, 2012; pp. 243–263. [Google Scholar]
- Liaw, A.; Wiener, M. Classification and regression by randomForest. R News 2002, 2, 18–22. [Google Scholar]
- Cutler, D.R.; Edwards, T.C., Jr.; Beard, K.H.; Cutler, A.; Hess, K.T.; Gibson, J.; Lawler, J.J. Random forests for classification in ecology. Ecology 2007, 88, 2783–2792. [Google Scholar] [CrossRef] [PubMed]
- Friedman, J.; Hastie, T.; Tibshirani, R. The Elements of Statistical Learning; Vol. 1 Springer Series in Statistics; Springer: New York, NY, USA, 2001. [Google Scholar]
- Bourel, M.; Segura, A. Multiclass classification methods in ecology. Ecol. Indic. 2018, 85, 1012–1021. [Google Scholar] [CrossRef]
- Crisci, C.; Ghattas, B.; Perera, G. A review of supervised machine learning algorithms and their applications to ecological data. Ecol. Model. 2012, 240, 113–122. [Google Scholar] [CrossRef]
- Strobl, C.; Boulesteix, A.-L.; Kneib, T.; Augustin, T.; Zeileis, A. Conditional variable importance for random forests. BMC Bioinform. 2008, 9, 307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hair, J.F.; Black, W.C.; Babin, B.J.; Anderson, R.E.; Tatham, R.L. Multivariate data analysis 6th Edition. J. Abnorm. Psychol. 2006, 87, 49–74. [Google Scholar]
- Biau, G.; Scornet, E. A random forest guided tour. Test 2016, 25, 197–227. [Google Scholar]
- Breiman, L.; Friedman, J.H.; Olshen, R.A.; Stone, C.J. Classification and Regression Trees; Routledge: London, UK, 2017. [Google Scholar]
- Reed, J.F., III. Homogeneity of kappa statistics in multiple samples. Comput. Methods Programs Biomed. 2000, 63, 43–46. [Google Scholar] [CrossRef]
- Falk, W.; Hempelmann, N. Species favourability shift in europe due to climate change: A case study for Fagus sylvatica L. and Picea abies (L.) Karst. based on an ensemble of climate models. J. Climatol. 2013, 2013, 787250. [Google Scholar] [CrossRef]
- Thurm, E.A.; Hernandez, L.; Baltensweiler, A.; Ayan, S.; Rasztovits, E.; Bielak, K.; Zlatanov, T.M.; Hladnik, D.; Balic, B.; Freudenschuss, A. Alternative tree species under climate warming in managed European forests. For. Ecol. Manag. 2018, 430, 485–497. [Google Scholar]
- Schlyter, P.; Stjernquist, I.; Bärring, L.; Jönsson, A.M.; Nilsson, C. Assessment of the impacts of climate change and weather extremes on boreal forests in northern Europe, focusing on Norway spruce. Clim. Res. 2006, 31, 75–84. [Google Scholar] [CrossRef]
- Solberg, S. Summer drought: A driver for crown condition and mortality of Norway spruce in Norway. For. Pathol. 2004, 34, 93–104. [Google Scholar] [CrossRef]
- Woodward, F.I.; Williams, B. Climate and plant distribution at global and local scales. Vegetatio 1987, 69, 189–197. [Google Scholar] [CrossRef]
- Penuelas, J.; Boada, M. A global change-induced biome shift in the Montseny mountains (NE Spain). Glob. Chang. Biol. 2003, 9, 131–140. [Google Scholar] [CrossRef] [Green Version]
- Jump, A.S.; Hunt, J.M.; Penuelas, J. Rapid climate change-related growth decline at the southern range edge of Fagus sylvatica. Glob. Chang. Biol. 2006, 12, 2163–2174. [Google Scholar] [CrossRef] [Green Version]
- Rigling, A.; Bigler, C.; Eilmann, B.; Feldmeyer-Christe, E.; Gimmi, U.; Ginzler, C.; Graf, U.; Mayer, P.; Vacchiano, G.; Weber, P. Driving factors of a vegetation shift from Scots pine to pubescent oak in dry Alpine forests. Glob. Chang. Biol. 2013, 19, 229–240. [Google Scholar] [CrossRef]
- Vacchiano, G.; Motta, R. An improved species distribution model for Scots pine and downy oak under future climate change in the NW Italian Alps. Ann. For. Sci. 2015, 72, 321–334. [Google Scholar] [CrossRef] [Green Version]
- Pasta, S.; De Rigo, D.; Caudullo, G. Quercus pubescens in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; Publications Office of the European Union: Brussels, Belgium, 2016; pp. 156–157. [Google Scholar]
- Ruiz-Labourdette, D.; Schmitz, M.F.; Pineda, F.D. Changes in tree species composition in Mediterranean mountains under climate change: Indicators for conservation planning. Ecol. Indic. 2013, 24, 310–323. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Zouber, A.; Lasanta, T.; Pueyo, Y. Dryness is accelerating degradation of vulnerable shrublands in semiarid Mediterranean environments. Ecol. Monogr. 2012, 82, 407–428. [Google Scholar] [CrossRef] [Green Version]
- Cheval, S.; Dumitrescu, A.; Birsan, M.-V. Variability of the aridity in the South-Eastern Europe over 1961–2050. Catena 2017, 151, 74–86. [Google Scholar] [CrossRef]
- Kertész, A.; Mika, J. Aridification—Climate change in South-Eastern Europe. Phys. Chem. Earth Part A Solid Earth Geod. 1999, 24, 913–920. [Google Scholar] [CrossRef]
- Svenning, J.C.; Sandel, B. Disequilibrium vegetation dynamics under future climate change. Am. J. Bot. 2013, 100, 1266–1286. [Google Scholar] [CrossRef] [PubMed]
- Ohlemüller, R.; Anderson, B.J.; Araújo, M.B.; Butchart, S.H.; Kudrna, O.; Ridgely, R.S.; Thomas, C.D. The coincidence of climatic and species rarity: High risk to small-range species from climate change. Biol. Lett. 2008, 4, 568–572. [Google Scholar] [CrossRef] [Green Version]
- Araújo, M.B.; Alagador, D.; Cabeza, M.; Nogués-Bravo, D.; Thuiller, W. Climate change threatens European conservation areas. Ecol. Lett. 2011, 14, 484–492. [Google Scholar] [CrossRef] [Green Version]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.T. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Carnicer, J.; Coll, M.; Ninyerola, M.; Pons, X.; Sanchez, G.; Penuelas, J. Widespread crown condition decline, food web disruption, and amplified tree mortality with increased climate change-type drought. Proc. Natl. Acad. Sci. USA 2011, 108, 1474–1478. [Google Scholar] [CrossRef] [Green Version]
- Senf, C.; Buras, A.; Zang, C.S.; Rammig, A.; Seidl, R. Excess forest mortality is consistently linked to drought across Europe. Nat. Commun. 2020, 11, 6200. [Google Scholar] [CrossRef]
- Taccoen, A.; Piedallu, C.; Seynave, I.; Perez, V.; Gégout-Petit, A.; Nageleisen, L.-M.; Bontemps, J.-D.; Gégout, J.-C. Background mortality drivers of European tree species: Climate change matters. Proc. R. Soc. B 2019, 286, 20190386. [Google Scholar] [CrossRef] [Green Version]
- Feurdean, A.; Bhagwat, S.A.; Willis, K.J.; Birks, H.J.B.; Lischke, H.; Hickler, T. Tree migration-rates: Narrowing the gap between inferred post-glacial rates and projected rates. PLoS ONE 2013, 8, e71797. [Google Scholar] [CrossRef]
- McKenney, D.W.; Pedlar, J.H.; Lawrence, K.; Campbell, K.; Hutchinson, M.F. Potential impacts of climate change on the distribution of North American trees. BioScience 2007, 57, 939–948. [Google Scholar] [CrossRef]
- Hanewinkel, M.; Cullmann, D.A.; Michiels, H.-G.; Kändler, G. Converting probabilistic tree species range shift projections into meaningful classes for management. J. Environ. Manag. 2014, 134, 153–165. [Google Scholar] [CrossRef]
- Elith, J.; Kearney, M.; Phillips, S. The art of modelling range-shifting species. Methods Ecol. Evol. 2010, 1, 330–342. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vegetation | CaPV Class | n | Sensitivity | Specificity | Balanced Accuracy |
|---|---|---|---|---|---|
| Polar deserts, subnival-nival vegetation of high mountains and glaciers | A | 3192 | 0.69 | 1 | 0.85 |
| Arctic tundras | B1 | 19,280 | 0.91 | 1 | 0.95 |
| Alpine vegetation | B2 | 11,194 | 0.67 | 0.99 | 0.83 |
| Eastern boreal open woodlands | C1 | 4396 | 0.59 | 1 | 0.79 |
| Western boreal and nemoral-montane Betula forests | C2 | 8350 | 0.58 | 0.99 | 0.79 |
| Subalpine and oro-Mediterranean vegetation | C3 | 4769 | 0.46 | 1 | 0.73 |
| Western boreal Picea forests | D1 | 68,887 | 0.86 | 0.98 | 0.92 |
| Eastern boreal Pinus-Picea and Abies-Picea forests | D2 | 14,678 | 0.82 | 1 | 0.91 |
| Hemiboreal Picea and Abies-Picea forests | D3 | 35,219 | 0.83 | 0.99 | 0.91 |
| Montane to altimontane, partly submontane Abies and Picea forests | D4 | 5051 | 0.44 | 0.99 | 0.71 |
| Boreal and hemiboreal Pinus forests (D5) + Montane to altimontane (subalpine) Pinus forests (D6) | D5 | 61,311 | 0.65 | 0.96 | 0.81 |
| Species-poor acidophilous Quercus and mixed Quercus forests | F1 | 27,788 | 0.67 | 0.98 | 0.82 |
| Mixed Quercus-Fraxinus forests | F2 | 7953 | 0.85 | 1 | 0.92 |
| Mixed Quercus-Carpinus forests | F3 | 35,102 | 0.71 | 0.98 | 0.85 |
| Tilia-Q.robur forests | F4 | 17,559 | 0.68 | 0.99 | 0.84 |
| Fagus and mixed Fagus forests (F5) + Fagus orientalis forests and Carpinus-Fagus orientalis forests (F6) | F5 | 60,531 | 0.85 | 0.98 | 0.91 |
| Caucasian mixed Carpinus-Quercus forests | F7 | 4444 | 0.69 | 1 | 0.85 |
| Subcontinental thermophilous (mixed) Q. robur L. and Q. petraea Liebl. forests | G1 | 3931 | 0.6 | 1 | 0.8 |
| Sub-Mediterranean-subcontinental thermophilous Q. cerris L. and Q. frainetto Ten. forests | G2 | 14,453 | 0.75 | 0.99 | 0.87 |
| Sub-Mediterranean and meso-supra-Mediterranean Q. pubescens Willd. forests | G3 | 13,386 | 0.63 | 0.99 | 0.81 |
| Iberian supra- and meso-Mediterranean Q. pyrenaica Willd., Q. faginea Lam., Q. faginea subsp. broteroi Cout. and Q. canariensis Willd. forests | G4 | 5068 | 0.68 | 1 | 0.84 |
| Meso- and supra-Mediterranean, as well as relict sclerophyllous forests | J1 | 25,968 | 0.89 | 0.99 | 0.94 |
| Thermo-Mediterranean sclerophyllous forests and xerophytic scrub | J2 | 6579 | 0.82 | 1 | 0.91 |
| Subcontinental meadow steppes and steppe-like dry grassland alternating with Q. robur forests | L1 | 23,214 | 0.74 | 0.99 | 0.87 |
| Sub-Mediterranean-subcontinental herb-grass steppes, partly meadow steppes alternating with oak forests | L2 | 3094 | 0.79 | 1 | 0.89 |
| True steppes | M1 | 45,652 | 0.94 | 0.99 | 0.97 |
| Desert steppes | M2 | 9106 | 0.92 | 1 | 0.96 |
| Northern lowland dwarf semishrub deserts | O1 | 7596 | 0.98 | 1 | 0.99 |
| Southern lowland-colline dwarf semishrub deserts with ephemeroids | O2 | 2298 | 0.94 | 1 | 0.97 |
| All classes | Mean | 18,967 | 0.75 | 0.99 | 0.87 |
| Parameter | Specification | Min | Max | Mean | SD |
|---|---|---|---|---|---|
| Bioclim 01 | Annual Mean Temperature (°C) | −13.3 | 19.6 | 6.5 | 4.9 |
| Bioclim 04 | Temperature Seasonality (SD (monthly means)) | 2.6 | 13.1 | 8.4 | 2.1 |
| Bioclim 12 | Annual Precipitation (mm/year) | 142 | 3773 | 667.6 | 273.1 |
| Bioclim 15 | Precipitation Seasonality (coefficient of variation) | 5 | 104 | 29.8 | 10.3 |
| Bioclim 18 | Precipitation of Warmest Quarter (mm/quarter) | 0 | 866 | 193.3 | 79.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hinze, J.; Albrecht, A.; Michiels, H.-G. Climate-Adapted Potential Vegetation—A European Multiclass Model Estimating the Future Potential of Natural Vegetation. Forests 2023, 14, 239. https://doi.org/10.3390/f14020239
Hinze J, Albrecht A, Michiels H-G. Climate-Adapted Potential Vegetation—A European Multiclass Model Estimating the Future Potential of Natural Vegetation. Forests. 2023; 14(2):239. https://doi.org/10.3390/f14020239
Chicago/Turabian StyleHinze, Jonas, Axel Albrecht, and Hans-Gerhard Michiels. 2023. "Climate-Adapted Potential Vegetation—A European Multiclass Model Estimating the Future Potential of Natural Vegetation" Forests 14, no. 2: 239. https://doi.org/10.3390/f14020239
APA StyleHinze, J., Albrecht, A., & Michiels, H. -G. (2023). Climate-Adapted Potential Vegetation—A European Multiclass Model Estimating the Future Potential of Natural Vegetation. Forests, 14(2), 239. https://doi.org/10.3390/f14020239