
Examining the Effect of Beech Bark Disease on Radial Growth in American Beech

Abstract
:1. Introduction
2. Methods
2.1. Site Description
2.2. Monitoring of Beech Bark Disease and Radial Growth
2.3. Model Development
2.4. Model Estimation, Specification, Validation, and Convergence
3. Results
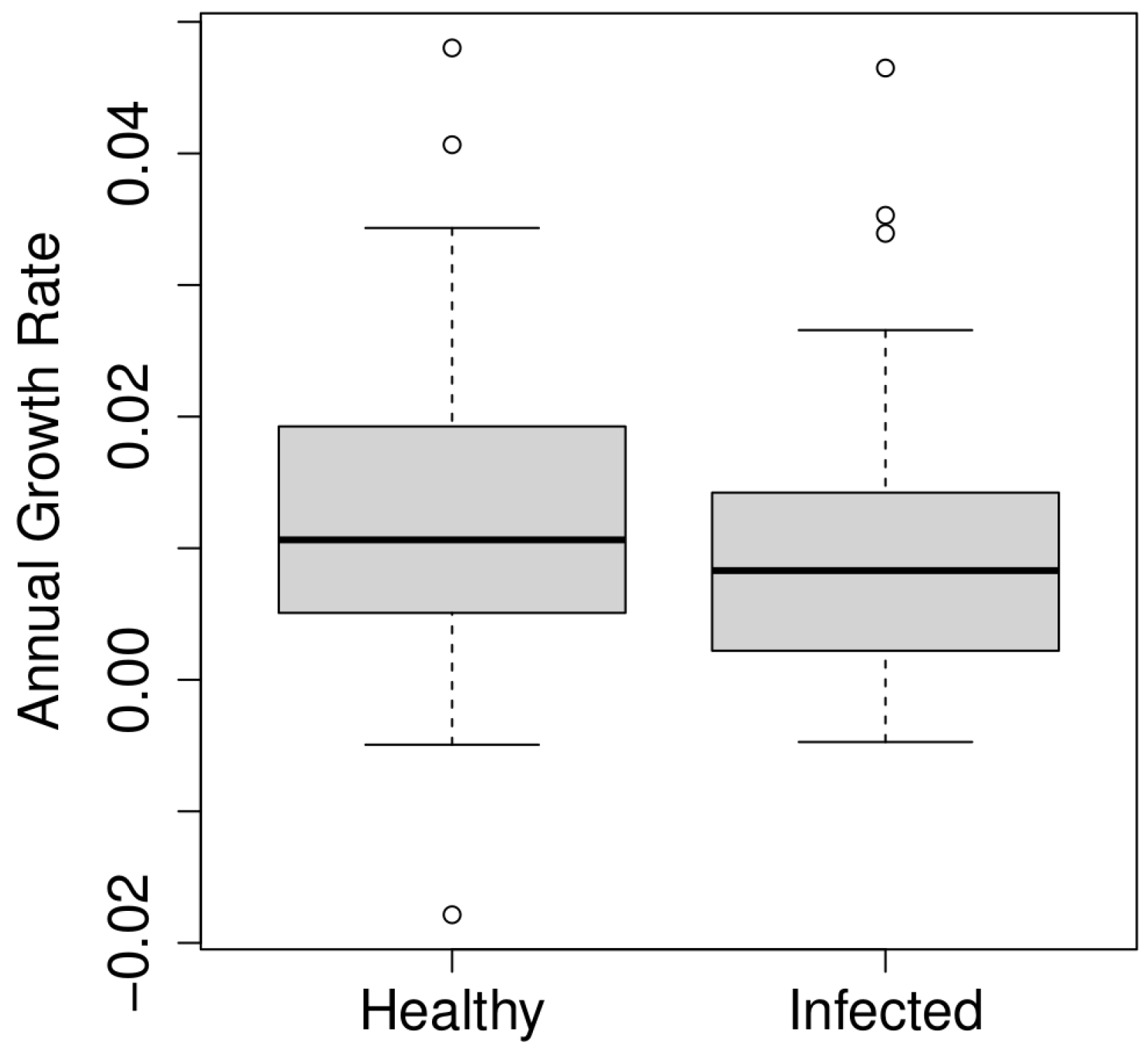
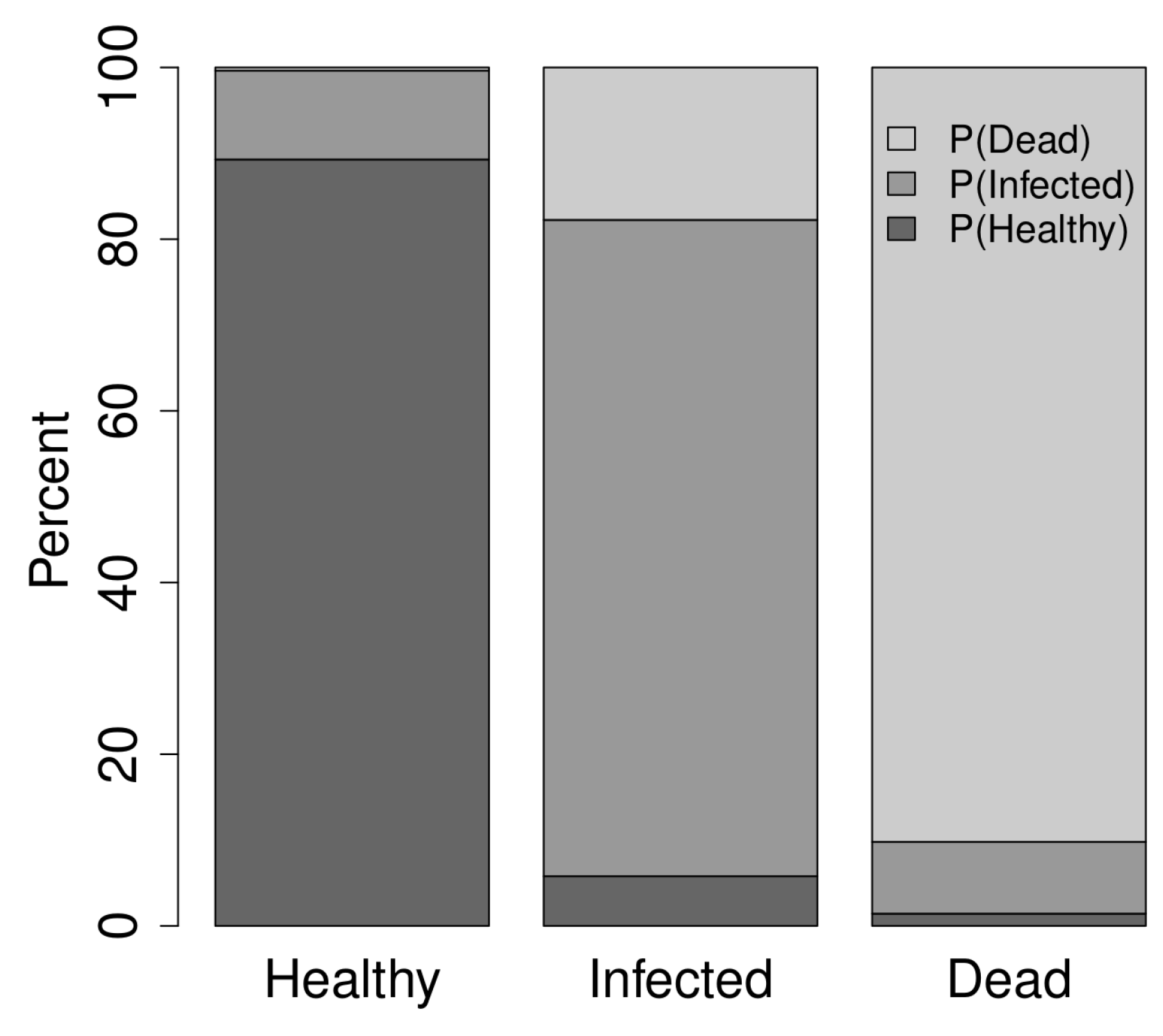
3.1. Overview of Data
3.2. Model Validation, Specification, and Convergence
3.3. Parameter Estimates
3.4. Observed Error Estimates
4. Discussion
Observer Error and Missing Data
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Klopfenstein, N.B.; Juzwik, J.; Ostry, M.E.; Kim, M.S.; Zambino, P.J.; Venette, R.C.; Richardson, B.A.; Lundquist, J.E.; Lodge, D.J.; Glaeser, J.A.; et al. Invasive forest pathogens: Summary of issues, critical needs, and future goals for forest service research and development. In A Dynamic Invasive Species Research Vision: Opportunities and Priorities 2009-29; Gen. Tech. Rep. WO-79/83; Dix, M.E., Britton, K., Eds.; US Department of Agriculture, Forest Service, Research and Development: Washington, DC, USA, 2010; Volume 79, pp. 23–33. [Google Scholar]
- Fisher, M.C.; Gow, N.A.; Gurr, S.J. Introduction: Tackling emerging fungal threats to animal health, food security and ecosystem resilience. Philos. Trans. R. Soc. Biol. Sci. 2016, 371, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Santini, A.; Ghelardini, L.; De Pace, C.; Desprez-Loustau, M.L.; Capretti, P.; Chandelier, A.; Cech, T.; Chira, D.; Diamandis, S.; Gaitniekis, T.; et al. Biogeographical patterns and determinants of invasion by forest pathogens in Europe. New Phytol. 2013, 197, 238–250. [Google Scholar] [CrossRef] [PubMed]
- Santini, A.; Battisti, A. Complex insect–pathogen interactions in tree pandemics. Front. Physiol. 2019, 10, 550. [Google Scholar] [CrossRef] [PubMed]
- Fleming, P.A.; Wentzel, J.J.; Dundas, S.J.; Kreplins, T.L.; Craig, M.D.; Hardy, G.E.S.J. Global meta-analysis of tree decline impacts on fauna. Biol. Rev. 2021, 96, 1744–1768. [Google Scholar] [CrossRef] [PubMed]
- Webster, C.R.; Dickinson, Y.L.; Burton, J.I.; Frelich, L.E.; Jenkins, M.A.; Kern, C.C.; Raymond, P.; Saunders, M.R.; Walters, M.B.; Willis, J.L. Promoting and maintaining diversity in contemporary hardwood forests: Confronting contemporary drivers of change and the loss of ecological memory. For. Ecol. Manag. 2018, 421, 98–108. [Google Scholar] [CrossRef]
- Prospero, S.; Botella, L.; Santini, A.; Robin, C. Biological control of emerging forest diseases: How can we move from dreams to reality? For. Ecol. Manag. 2021, 496, 119377. [Google Scholar] [CrossRef]
- Lamarche, J.; Potvin, A.; Pelletier, G.; Stewart, D.; Feau, N.; Alayon, D.I.; Dale, A.L.; Coelho, A.; Uzunovic, A.; Bilodeau, G.J.; et al. Molecular detection of 10 of the most unwanted alien forest pathogens in Canada using real-time PCR. PLoS ONE 2015, 10, e0134265. [Google Scholar] [CrossRef]
- Cale, J.A.; Garrison-Johnston, M.T.; Teale, S.A.; Castello, J.D. Beech bark disease in North America: Over a century of research revisited. For. Ecol. Manag. 2017, 394, 86–103. [Google Scholar] [CrossRef]
- Ehrlich, J. The beech bark disease: A Nectria disease of Fagus, following Cryptococcus fagi (Baer.). Can. J. Res. 1934, 10, 593–692. [Google Scholar] [CrossRef]
- Houston, D.R.; O’Brien, J. Beech Bark Disease; U.S. Department of Agriculture Forest Service: Washington, DC, USA, 2003. [Google Scholar]
- Stephanson, C.A.; Coe, N.R. Impacts of beech bark disease and climate change on American beech. Forests 2017, 8, 155. [Google Scholar] [CrossRef] [Green Version]
- Tubbs, C.H.; Houston, D.R. Fagus grandifolia Ehrh. American beech. Silvics N. Am. 1990, 2, 325. [Google Scholar]
- Garneau, D.E.; Lawler, M.E.; Rumpf, A.S.; Weyburne, E.S.; Cuppernull, T.M.; Boe, A.G. Potential effects of beech bark disease on small mammals and invertebrates in northeastern US forests. Northeast. Nat. 2012, 19, 391–410. [Google Scholar] [CrossRef]
- McCullough, D.G.; Heyd, R.L.; O’Brien, J.G. Biology and Management of Beech Bark Disease; Extension Bulletin E-2746; Michigan State University: East Lansing, MI, USA, 2001; Available online: https://www.canr.msu.edu/uploads/files/e2746.pdf (accessed on 3 January 2023).
- Nevins, M.T.; D’Amato, A.W.; Foster, J.R. Future forest composition under a changing climate and adaptive forest management in southeastern Vermont, USA. For. Ecol. Manag. 2021, 479, 118527. [Google Scholar] [CrossRef]
- Burns, R.M.; Honkala, B. Summary of tree characteristics. Silvics N. Am. 1990, 2, 846–849. [Google Scholar]
- Halman, J.M.; Schaberg, P.G.; Hawley, G.J.; Hansen, C.F.; Fahey, T.J. Differential impacts of calcium and aluminum treatments on sugar maple and American beech growth dynamics. Can. J. For. Res. 2015, 45, 52–59. [Google Scholar] [CrossRef]
- Hamelin, P.L. VT ANR Management Guidelines for Optimizing Mast Yields in Beech Mast Production Areas; Vermont Fish & Wildlife Department: Waterbury, VT, USA, 2011. [Google Scholar]
- Dracup, E.C.; MacLean, D.A. Partial harvest to reduce occurrence of American beech affected by beech bark disease: 10 year results. For. Int. J. For. Res. 2018, 91, 73–82. [Google Scholar] [CrossRef]
- Bernard, A.; Gélinas, N.; Duchateau, E.; Durocher, C.; Achim, A. American beech in value-added hardwood products: Assessing consumer preferences. BioResources 2018, 13, 6893–6910. [Google Scholar] [CrossRef]
- Gómez-Cortecero, A.; Saville, R.J.; Scheper, R.W.; Bowen, J.K.; Agripino De Medeiros, H.; Kingsnorth, J.; Xu, X.; Harrison, R.J. Variation in host and pathogen in the Neonectria/Malus interaction; toward an understanding of the genetic basis of resistance to European canker. Front. Plant Sci. 2016, 7, 1365. [Google Scholar] [CrossRef]
- Mason, M.E.; Koch, J.L.; Krasowski, M.; Loo, J. Comparisons of protein profiles of beech bark disease resistant and susceptible American beech (Fagus grandifolia). Proteome Sci. 2013, 11, 1–21. [Google Scholar] [CrossRef]
- Seitler, K.; Coe, N.R. cDNA primers for quantitative analysis of protective compounds implicated in beech bark disease resistance in American beech, Fagus grandifolia. Conserv. Genet. Resour. 2015, 7, 689–691. [Google Scholar] [CrossRef]
- Kasson, M.; Livingston, W. Relationships among beech bark disease, climate, radial growth response and mortality of American beech in northern Maine, USA. For. Pathol. 2012, 42, 199–212. [Google Scholar] [CrossRef]
- Houston, D.R. American beech resistance to Cryptococcus fagisuga. In Sponsored by the USDA Forest Service, Northeastern Forest Experiment Station, Proceedings of the IUFRO Beech Bark Disease Working Party Conference, Hamden, CT, USA, 26 September–8 October 1982; Gen. Tech. Rep. WO-37; US Department of Agriculture, Forest Service: Washington, DC, USA, 1983; Volume 37, pp. 38–42. [Google Scholar]
- Hane, E.N. Indirect effects of beech bark disease on sugar maple seedling survival. Can. J. For. Res. 2003, 33, 807–813. [Google Scholar] [CrossRef]
- Petrillo, H.A.; Witter, J.A. Regeneration of American beech (Fagus grandifolia Ehrh.) in Michigan: Interactions of beech bark disease and management practices. In Beech Bark Disease: Proceedings of the Beech Bark Disease Symposium; Gen. Tech. Report NE-331; USDA, Forest Service: Washington, DC, USA, 2005; pp. 142–145. Available online: https://www.fs.usda.gov/ne/newtown_square/publications/technical_reports/pdfs/2005/331papers/petrillo331-3.pdf (accessed on 3 January 2023).
- McLaughlin, J.; Greifenhagen, S. Beech bark disease in Ontario: A primer and management recommendations. For. Res.-Note-Ont. For. Res. Inst. 2012, 71, 1–8. [Google Scholar]
- Koch, J.L.; Carey, D.W.; Mason, M.E.; Nelson, C.D. Assessment of beech scale resistance in full-and half-sibling American beech families. Can. J. For. Res. 2010, 40, 265–272. [Google Scholar] [CrossRef]
- Koch, J.L.; Mason, M.E.; Carey, D.W. Screening for resistance to beech bark disease: Improvements and results from seedlings and grafted field selections. In Disease and Insect Resistance in Forest Trees, Proceedings of the Fourth International Workshop on the Genetics of Host-Parasite Interactions in Forestry; Gen. Tech. Rep. PSW-GTR-240; Sniezko, R.A., Yanchuk, A.D., Kliejunas, J.T., Palmieri, K.M., Alexander, J.M., Frankel, S.J., Eds.; Pacific Southwest Research Station, Forest Service, US Department of Agriculture: Albany, CA, USA, 2012; Volume 240, pp. 196–208. [Google Scholar]
- Bose, A.K.; Weiskittel, A.; Wagner, R.G. Occurrence, pattern of change, and factors associated with American beech-dominance in stands of the northeastern USA forest. For. Ecol. Manag. 2017, 392, 202–212. [Google Scholar] [CrossRef]
- Forrester, J.A.; McGee, G.G.; Mitchell, M.J. Effects of beech bark disease on aboveground biomass and species composition in a mature northern hardwood forest, 1985 to 2000. J. Torrey Bot. Soc. 2003, 130, 70–78. [Google Scholar] [CrossRef]
- Latty, E.F. Stand-level patterns and ecosystem consequences of beach bark disease. In Proceedings of the Beech Bark Disease Symposium, Saranac Lake, NY, USA, 16–18 June 2004; General Technical Report NE-331. USDA Forest Service, Northern Research Station: Newtown Square, PA, USA, 2005; pp. 36–42. [Google Scholar]
- Rumble, L.; Taylor, G.; Grinath, J.B.; Morris, A.B. Measuring spatial and temporal shifts in forest structure and composition in high elevation beech forests in response to beech bark disease in Great Smoky Mountains National Park. For. Ecol. Manag. 2020, 461, 117954. [Google Scholar] [CrossRef]
- Cale, J.A.; McNulty, S.A. Not dead yet: Beech trees can survive nearly three decades in the aftermath phase of a deadly forest disease complex. For. Ecol. Manag. 2018, 409, 372–377. [Google Scholar] [CrossRef]
- Gove, J.; Houston, D.R. Monitoring the growth of American beech affected by beech bark disease in Maine using the Kalman filter. Environ. Ecol. Stat. 1996, 3, 167–187. [Google Scholar] [CrossRef]
- Božić, M.; Đureta, F.; Goršić, E.; Vedriš, M. Influence of a measurer and measurement errors on measurement of a tree diameter. Šumarski List. 2020, 144, 233–244. [Google Scholar] [CrossRef]
- Dalla Corte, A.P.; Souza, D.V.; Rex, F.E.; Sanquetta, C.R.; Mohan, M.; Silva, C.A.; Zambrano, A.M.A.; Prata, G.; de Almeida, D.R.A.; Trautenmüller, J.W.; et al. Forest inventory with high-density UAV-Lidar: Machine learning approaches for predicting individual tree attributes. Comput. Electron. Agric. 2020, 179, 105815. [Google Scholar] [CrossRef]
- Pretzsch, H. The course of tree growth. Theory and reality. For. Ecol. Manag. 2020, 478, 118508. [Google Scholar] [CrossRef]
- Griffin, J.M.; Lovett, G.M.; Arthur, M.A.; Weathers, K.C. The distribution and severity of beech bark disease in the Catskill Mountains, NY. Can. J. For. Res. 2003, 33, 1754–1760. [Google Scholar] [CrossRef] [Green Version]
- Mulder, O.; Sleith, R.; Mulder, K.; Coe, N.R. A Bayesian analysis of topographic influences on the presence and severity of beech bark disease. For. Ecol. Manag. 2020, 472, 118198. [Google Scholar] [CrossRef]
- Hobbs, N.T.; Hooten, M.B. Bayesian Models; Princeton University Press: Princeton, NJ, USA, 2015. [Google Scholar]
- Wade, P.R. Bayesian methods in conservation biology. Conserv. Biol. 2000, 14, 1308–1316. [Google Scholar] [CrossRef]
- Wikle, C.K. Hierarchical Bayesian models for predicting the spread of ecological processes. Ecology 2003, 84, 1382–1394. [Google Scholar] [CrossRef]
- Farnsworth, M.L.; Hoeting, J.A.; Hobbs, N.T.; Miller, M.W. Linking chronic wasting disease to mule deer movement scales: A hierarchical Bayesian approach. Ecol. Appl. 2006, 16, 1026–1036. [Google Scholar] [CrossRef]
- Ellison, A.M. Bayesian inference in ecology. Ecol. Lett. 2004, 7, 509–520. [Google Scholar] [CrossRef]
- Shanahan, E.; Wright, W.J.; Irvine, K.M. Adaptive monitoring in action: Reconsidering design-based estimators reveals underestimation of whitebark pine disease prevalence in the Greater Yellowstone Ecosystem. J. Appl. Ecol. 2021, 58, 1079–1089. [Google Scholar] [CrossRef]
- Richardson, S.J.; Hurst, J.M.; Easdale, T.A.; Wiser, S.K.; Griffiths, A.D.; Allen, R.B. Diameter growth rates of beech (Nothofagus) trees around New Zealand. New Zealand J. For. 2011, 56, 3–11. [Google Scholar]
- Gelfand, A.E.; Smith, A.F. Sampling-based approaches to calculating marginal densities. J. Am. Stat. Assoc. 1990, 85, 398–409. [Google Scholar] [CrossRef]
- King, R.; Morgan, B.; Gimenez, O.; Brooks, S. Bayesian Analysis for Population Ecology; Chapman and Hall/CRC: Boca Raton, FL, USA, 2009. [Google Scholar]
- Gelman, A.; Rubin, D.B. Inference from iterative simulation using multiple sequences. Stat. Sci. 1992, 7, 457–472. [Google Scholar] [CrossRef]
- Conn, P.B.; Johnson, D.S.; Williams, P.J.; Melin, S.R.; Hooten, M.B. A guide to Bayesian model checking for ecologists. Ecol. Monogr. 2018, 88, 526–542. [Google Scholar] [CrossRef]
- Bayat, M.; Bettinger, P.; Heidari, S.; Hamidi, S.K.; Jaafari, A. A Combination of Biotic and Abiotic Factors and Diversity Determine Productivity in Natural Deciduous Forests. Forests 2021, 12, 1450. [Google Scholar] [CrossRef]
- Luo, D.; Huang, J.G.; Jiang, X.; Ma, Q.; Liang, H.; Guo, X.; Zhang, S. Effect of climate and competition on radial growth of Pinus massoniana and Schima superba in China’s subtropical monsoon mixed forest. Dendrochronologia 2017, 46, 24–34. [Google Scholar] [CrossRef]
- Huang, X.; Dai, D.; Xiang, Y.; Yan, Z.; Teng, M.; Wang, P.; Zhou, Z.; Zeng, L.; Xiao, W. Radial growth of Pinus massoniana is influenced by temperature, precipitation, and site conditions on the regional scale: A meta-analysis based on tree-ring width index. Ecol. Indic. 2021, 126, 107659. [Google Scholar] [CrossRef]
- Orwig, D.A.; Abrams, M.D. Variation in radial growth responses to drought among species, site, and canopy strata. Trees 1997, 11, 474–484. [Google Scholar] [CrossRef]
- Latte, N.; Perin, J.; Kint, V.; Lebourgeois, F.; Claessens, H. Major changes in growth rate and growth variability of beech (Fagus sylvatica L.) related to soil alteration and climate change in Belgium. Forests 2016, 7, 174. [Google Scholar] [CrossRef]
- Zweifel, R.; Zimmermann, L.; Zeugin, F.; Newbery, D.M. Intra-annual radial growth and water relations of trees: Implications towards a growth mechanism. J. Exp. Bot. 2006, 57, 1445–1459. [Google Scholar] [CrossRef]
- Čufar, K.; De Luis, M.; Prislan, P.; Gričar, J.; Črepinšek, Z.; Merela, M.; Kajfež-Bogataj, L. Do variations in leaf phenology affect radial growth variations in Fagus sylvatica? Int. J. Biometeorol. 2015, 59, 1127–1132. [Google Scholar] [CrossRef]
- Kimberley, M.; Hood, I.; Knowles, R. Impact of Swiss needle-cast on growth of Douglas-fir. Phytopathology 2011, 101, 583–593. [Google Scholar] [CrossRef]
- Bert, D.; Lasnier, J.B.; Capdevielle, X.; Dugravot, A.; Desprez-Loustau, M.L. Powdery mildew decreases the radial growth of oak trees with cumulative and delayed effects over years. PLoS ONE 2016, 11, e0155344. [Google Scholar] [CrossRef]
- Dietze, M.C.; Matthes, J.H. A general ecophysiological framework for modelling the impact of pests and pathogens on forest ecosystems. Ecol. Lett. 2014, 17, 1418–1426. [Google Scholar] [CrossRef]
- Elzinga, C.; Shearer, R.C.; Elzinga, G. Observer variation in tree diameter measurements. West. J. Appl. For. 2005, 20, 134–137. [Google Scholar] [CrossRef] [Green Version]
- Butt, N.; Slade, E.; Thompson, J.; Malhi, Y.; Riutta, T. Quantifying the sampling error in tree census measurements by volunteers and its effect on carbon stock estimates. Ecol. Appl. 2013, 23, 936–943. [Google Scholar] [CrossRef]
- Morrison, L.W. Observer error in vegetation surveys: A review. J. Plant Ecol. 2016, 9, 367–379. [Google Scholar] [CrossRef]
- Burg, S.; Rixen, C.; Stöckli, V.; Wipf, S. Observation bias and its causes in botanical surveys on high-alpine summits. J. Veg. Sci. 2015, 26, 191–200. [Google Scholar] [CrossRef]
- Barrett, T.; Maltomo, M. Missing data in forest ecology and management: Advances in quantitative methods [Preface]. For. Ecol. Manag. 2012, 572, 1–2. [Google Scholar] [CrossRef]
- Futschik, A.; Winkler, M.; Steinbauer, K.; Lamprecht, A.; Rumpf, S.B.; Barančok, P.; Palaj, A.; Gottfried, M.; Pauli, H. Disentangling observer error and climate change effects in long-term monitoring of alpine plant species composition and cover. J. Veg. Sci. 2020, 31, 14–25. [Google Scholar] [CrossRef]
- Pinkard, E.; Battaglia, M.; Roxburgh, S.; O’Grady, A. Estimating forest net primary production under changing climate: Adding pests into the equation. Tree Physiol. 2011, 31, 686–699. [Google Scholar] [CrossRef]
- Elphick, C.S. How you count counts: The importance of methods research in applied ecology. J. Appl. Ecol. 2008, 45, 1313–1320. [Google Scholar] [CrossRef]
- Ewing, C.J.; Hausman, C.E.; Pogacnik, J.; Slot, J.; Bonello, P. Beech leaf disease: An emerging forest epidemic. For. Pathol. 2019, 49, 12488. [Google Scholar] [CrossRef]
- Coe, N.R.; Mulder, O.; Worland, J.; Mulder, K. Beech Bark Disease Data 2005–2019. 2022. Available online: https://doi.org/10.5281/zenodo.6348119 (accessed on 3 January 2023).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Symbol | Definition |
|---|---|---|
| Overall growth rate coefficient | Mean growth rate coefficient (unitless) across all years. | |
| Average growth rate coefficient by year | Growth rate coefficient in year k. | |
| Growth rate exponent | Exponent for growth model. | |
| BBD coefficient | Proportion of average growth achieved by infected trees relative to healthy trees. | |
| Within year variability | Variability in growth rates within a year. | |
| Between year variability | Variability in growth coefficients () between years. | |
| Annual growth by tree by year | Growth of tree i in year k. | |
| Latent DBH | Diameter at breast height in cm for tree i in year k. | |
| Observer variability for DBH | Variability in observer error when measuring DBH. | |
| Observed DBH | Observed DBH for tree i in year j. | |
| Probability of infection by year | Probability of a healthy tree becoming infected during year k. | |
| Probability of death | Probability of an infected tree dying within the coming year. | |
| Latent infection status | Infection status (healthy, infected, or dead) of tree i in year k. | |
| Infection status observation probability matrix | 3 × 3 matrix. = probability of observing infection status n given latent status m where status ranges from healthy, infected, dead. | |
| Observed Infection Status | Observed infection status of tree i in year j. |
| Mean | 0.494 | 0.610 | 0.595 | 0.864 | |||
| Sd | |||||||
| 2.5% | 0.371 | 0.504 | 0.561 | 0.648 | |||
| Median | 0.490 | 0.614 | 0.594 | 0.880 | |||
| 97.5% | 0.640 | 0.700 | 0.630 | 0.993 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coe, N.; Mulder, O.; Mulder, K. Examining the Effect of Beech Bark Disease on Radial Growth in American Beech. Forests 2023, 14, 312. https://doi.org/10.3390/f14020312
Coe N, Mulder O, Mulder K. Examining the Effect of Beech Bark Disease on Radial Growth in American Beech. Forests. 2023; 14(2):312. https://doi.org/10.3390/f14020312
Chicago/Turabian StyleCoe, Natalie, Obadiah Mulder, and Kenneth Mulder. 2023. "Examining the Effect of Beech Bark Disease on Radial Growth in American Beech" Forests 14, no. 2: 312. https://doi.org/10.3390/f14020312
APA StyleCoe, N., Mulder, O., & Mulder, K. (2023). Examining the Effect of Beech Bark Disease on Radial Growth in American Beech. Forests, 14(2), 312. https://doi.org/10.3390/f14020312