
Allometric Growth of Common Urban Tree Species in Qingdao City of Eastern China



Abstract
:1. Introduction
2. Materials and Methods
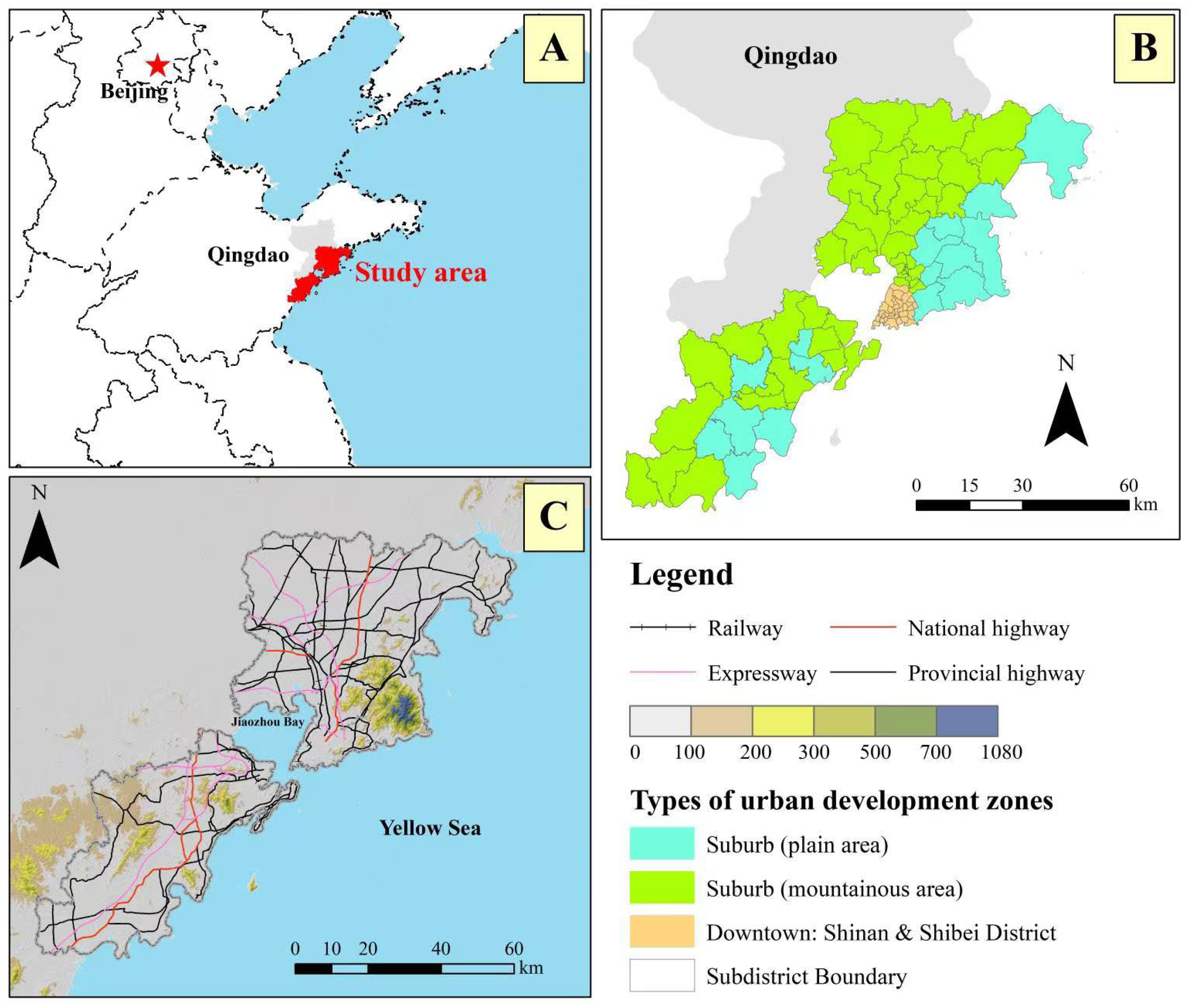
2.1. Site Description
2.2. Data Collection
2.3. Tree Allometric Growth Model
3. Results
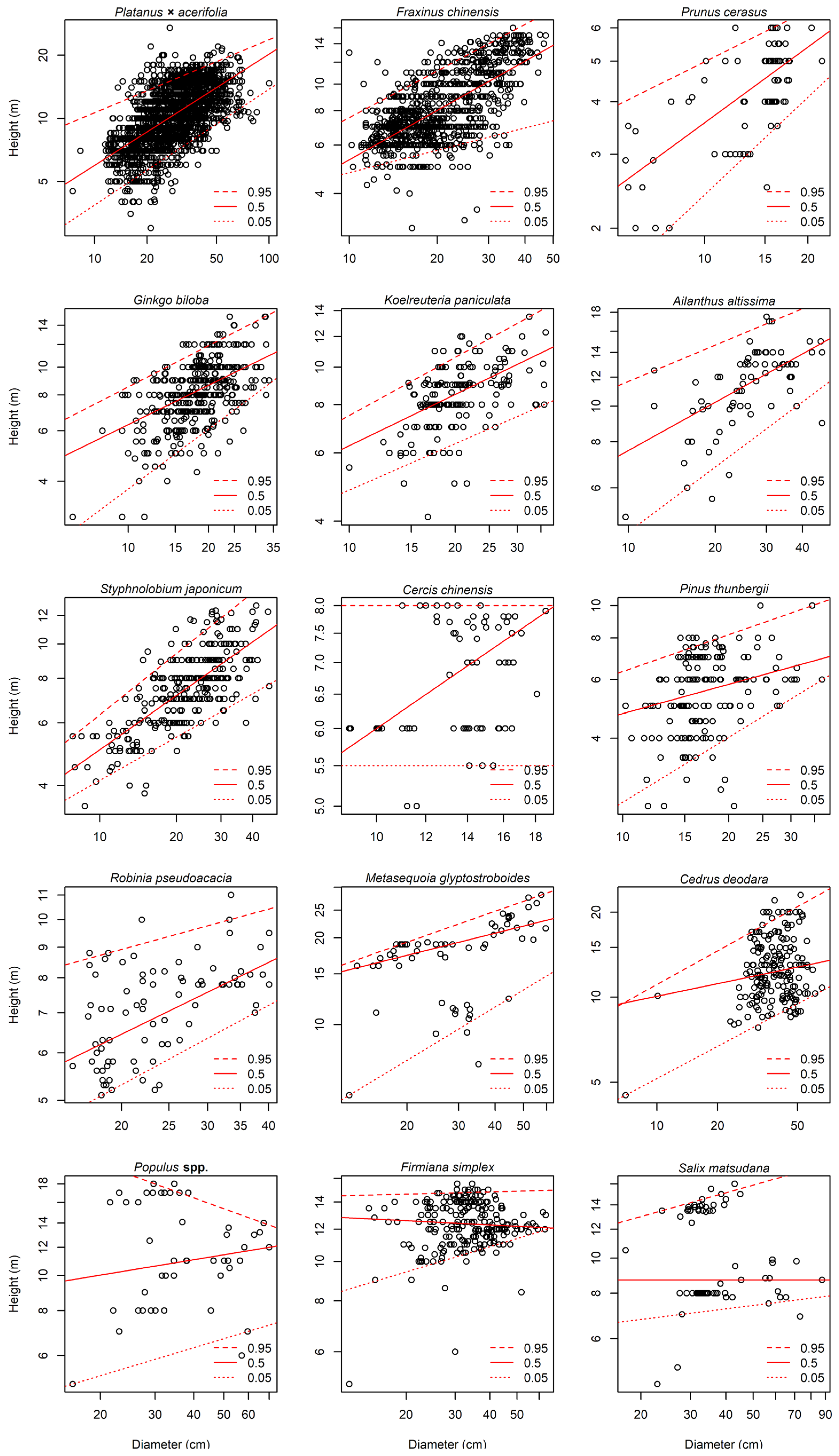
3.1. Allometric Growth Model of DBH and Tree Height
3.2. Allometric Growth Model of DBH and Crown Width
3.3. Allometric Relationships of Crown Projection Area and Crown Volume
4. Discussion
4.1. Allometric Models
4.2. Comparison of Allometric Growth of Different Tree Species
4.3. Factors Affecting Allometric Growth Relationship
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


References
- Gómez-Baggethun, E.; Barton, D.N. Classifying and Valuing Ecosystem Services for Urban Planning. Ecol. Econ. 2013, 86, 235–245. [Google Scholar] [CrossRef]
- Elmqvist, T.; Setälä, H.; Handel, S.; Van Der Ploeg, S.; Aronson, J.; Blignaut, J.N.; Gomez-Baggethun, E.; Nowak, D.; Kronenberg, J.; De Groot, R. Benefits of Restoring Ecosystem Services in Urban Areas. Curr. Opin. Environ. Sustain. 2015, 14, 101–108. [Google Scholar] [CrossRef] [Green Version]
- Esperon-Rodriguez, M.; Rymer, P.D.; Power, S.A.; Barton, D.N.; Cariñanos, P.; Dobbs, C.; Eleuterio, A.A.; Escobedo, F.J.; Hauer, R.; Hermy, M.; et al. Assessing Climate Risk to Support Urban Forests in a Changing Climate. Plants People Planet 2022, 4, 201–213. [Google Scholar] [CrossRef]
- Jim, C.Y.; Chen, W.Y. Ecosystem Services and Valuation of Urban Forests in China. Cities 2009, 26, 187–194. [Google Scholar] [CrossRef]
- Wang, X.; Yao, J.; Yu, S.; Miao, C.; Chen, W.; He, X. Street Trees in a Chinese Forest City: Structure, Benefits and Costs. Sustainability 2018, 10, 674. [Google Scholar] [CrossRef] [Green Version]
- Escobedo, F.J.; Kroeger, T.; Wagner, J.E. Urban Forests and Pollution Mitigation: Analyzing Ecosystem Services and Disservices. Environ. Pollut. 2011, 159, 2078–2087. [Google Scholar] [CrossRef]
- Roy, S.; Byrne, J.; Pickering, C. A Systematic Quantitative Review of Urban Tree Benefits, Costs, and Assessment Methods across Cities in Different Climatic Zones. Urban For. Urban Green. 2012, 11, 351–363. [Google Scholar] [CrossRef] [Green Version]
- Hardin, P.J.; Jensen, R.R. The Effect of Urban Leaf Area on Summertime Urban Surface Kinetic Temperatures: A Terre Haute Case Study. Urban For. Urban Green. 2007, 6, 63–72. [Google Scholar] [CrossRef]
- Escobedo, F.J.; Wagner, J.E.; Nowak, D.J.; De la Maza, C.L.; Rodriguez, M.; Crane, D.E. Analyzing the Cost Effectiveness of Santiago, Chile’s Policy of Using Urban Forests to Improve Air Quality. J. Environ. Manag. 2008, 86, 148–157. [Google Scholar] [CrossRef]
- Chaparro, L.; Terradas, J. Ecological Services of Urban Forest in Barcelona; Institut Municipal de Parcs i Jardins Ajuntament de Barcelona, Àrea de Medi Ambient: Bellaterra, Spain, 2009. [Google Scholar]
- Fang, C.-F.; Ling, D.-L. Investigation of the Noise Reduction Provided by Tree Belts. Landsc. Urban Plan. 2003, 63, 187–195. [Google Scholar] [CrossRef]
- Ma, B.; Hauer, R.J.; Xu, C.; Li, W. Visualizing Evaluation Model of Human Perceptions and Characteristic Indicators of Landscape Visual Quality in Urban Green Spaces by Using Nomograms. Urban For. Urban Green. 2021, 65, 127314. [Google Scholar] [CrossRef]
- Bosch-Cano, F.; Bernard, N.; Sudre, B.; Gillet, F.; Thibaudon, M.; Richard, H.; Badot, P.-M.; Ruffaldi, P. Human Exposure to Allergenic Pollens: A Comparison between Urban and Rural Areas. Environ. Res. 2011, 111, 619–625. [Google Scholar] [CrossRef] [Green Version]
- Calfapietra, C.; Fares, S.; Manes, F.; Morani, A.; Sgrigna, G.; Loreto, F. Role of Biogenic Volatile Organic Compounds (BVOC) Emitted by Urban Trees on Ozone Concentration in Cities: A Review. Environ. Pollut. 2013, 183, 71–80. [Google Scholar] [CrossRef]
- Klein, R.W.; Koeser, A.K.; Hauer, R.J.; Hansen, G.; Escobedo, F.J. Risk Assessment and Risk Perception of Trees: A Review of Literature Relating to Arboriculture and Urban Forestry. Arboric. Urban For. 2019, 45, 26–38. [Google Scholar] [CrossRef]
- Nowak, D.J. Institutionalizing Urban Forestry as a “Biotechnology” to Improve Environmental Quality. Urban For. Urban Green. 2006, 5, 93–100. [Google Scholar] [CrossRef]
- Vogt, J.; Fischer, B.C.; Hauer, R.J. Urban Forestry and Arboriculture as Interdisciplinary Environmental Science: Importance and Incorporation of Other Disciplines. J. Environ. Stud. Sci. 2016, 6, 371–386. [Google Scholar] [CrossRef]
- Huxley, J.S.; Teissier, G. Terminology of Relative Growth. Nature 1936, 137, 780–781. [Google Scholar] [CrossRef]
- Pretzsch, H.; Dieler, J. Evidence of Variant Intra-and Interspecific Scaling of Tree Crown Structure and Relevance for Allometric Theory. Oecologia 2012, 169, 637–649. [Google Scholar] [CrossRef] [Green Version]
- Fu, L.; Sharma, R.P.; Hao, K.; Tang, S. A Generalized Interregional Nonlinear Mixed-Effects Crown Width Model for Prince Rupprecht Larch in Northern China. For. Ecol. Manag. 2017, 389, 364–373. [Google Scholar] [CrossRef]
- Pretzsch, H.; Biber, P.; Uhl, E.; Dahlhausen, J.; Rötzer, T.; Caldentey, J.; Koike, T.; Van Con, T.; Chavanne, A.; Seifert, T.; et al. Crown Size and Growing Space Requirement of Common Tree Species in Urban Centres, Parks, and Forests. Urban For. Urban Green. 2015, 14, 466–479. [Google Scholar] [CrossRef] [Green Version]
- Pretzsch, H.; Matthew, C.; Dieler, J. Allometry of Tree Crown Structure. Relevance for Space Occupation at the Individual Plant Level and for Self-Thinning at the Stand Level. In Growth and Defence in Plants; Springer: Berlin/Heidelberg, Germany, 2012; pp. 287–310. [Google Scholar]
- Randrup, T.; McPherson, E.; Costello, L. A Review of Tree Root Conflicts with Sidewalks, Curbs, and Roads. Urban Ecosyst. 2001, 5, 209–225. [Google Scholar] [CrossRef]
- Harrison, P.A.; Dunford, R.; Barton, D.N.; Kelemen, E.; Martín-López, B.; Norton, L.; Termansen, M.; Saarikoski, H.; Hendriks, K.; Gómez-Baggethun, E.; et al. Selecting Methods for Ecosystem Service Assessment: A Decision Tree Approach. Ecosyst. Serv. 2018, 29, 481–498. [Google Scholar] [CrossRef] [Green Version]
- Tyrväinen, L. Economic Valuation of Urban Forest Benefits in Finland. J. Environ. Manag. 2001, 62, 75–92. [Google Scholar] [CrossRef] [PubMed]
- Berland, A. Urban Tree Growth Models for Two Nearby Cities Show Notable Differences. Urban Ecosyst. 2020, 23, 1253–1261. [Google Scholar] [CrossRef]
- Hauer, R.J.; Hanou, I.S.; Sivyer, D. Planning for Active Management of Future Invasive Pests Affecting Urban Forests: The Ecological and Economic Effects of Varying Dutch Elm Disease Management Practices for Street Trees in Milwaukee, WI USA. Urban Ecosyst. 2020, 23, 1005–1022. [Google Scholar] [CrossRef]
- Vannatta, A.; Hauer, R.; Schuettpelz, N. Economic Analysis of Emerald Ash Borer (Coleoptera: Buprestidae) Management Options. J. Econ. Éntomol. 2012, 105, 196–206. [Google Scholar] [CrossRef]
- Pretzsch, H.; Biber, P.; Uhl, E.; Dahlhausen, J.; Schütze, G.; Perkins, D.; Rötzer, T.; Caldentey, J.; Koike, T.; van Con, T.; et al. Climate Change Accelerates Growth of Urban Trees in Metropolises Worldwide. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Monteiro, M.V.; Doick, K.J.; Handley, P. Allometric Relationships for Urban Trees in Great Britain. Urban For. Urban Green. 2016, 19, 223–236. [Google Scholar] [CrossRef]
- Timilsina, N.; Beck, J.L.; Eames, M.S.; Hauer, R.; Werner, L. A Comparison of Local and General Models of Leaf Area and Biomass of Urban Trees in USA. Urban For. Urban Green. 2017, 24, 157–163. [Google Scholar] [CrossRef] [Green Version]
- Day, S.D.; Eric Wiseman, P.; Dickinson, S.B.; Roger Harris, J. Tree Root Ecology in the Urban Environment and Implications for a Sustainable Rhizosphere. Arboric. Urban For. 2010, 36, 193. [Google Scholar] [CrossRef]
- Troxel, B.; Piana, M.; Ashton, M.S.; Murphy-Dunning, C. Relationships between Bole and Crown Size for Young Urban Trees in the Northeastern USA. Urban For. Urban Green. 2013, 12, 144–153. [Google Scholar] [CrossRef]
- Yoon, T.K.; Park, C.-W.; Lee, S.J.; Ko, S.; Kim, K.N.; Son, Y.; Lee, K.H.; Oh, S.; Lee, W.-K.; Son, Y. Allometric Equations for Estimating the Aboveground Volume of Five Common Urban Street Tree Species in Daegu, Korea. Urban For. Urban Green. 2013, 12, 344–349. [Google Scholar] [CrossRef]
- McHale, M.; Burke, I.; Lefsky, M.; Peper, P.; McPherson, E. Urban Forest Biomass Estimates: Is It Important to Use Allometric Relationships Developed Specifically for Urban Trees? Urban Ecosyst. 2009, 12, 95–113. [Google Scholar] [CrossRef]
- Moser, A.; Rötzer, T.; Pauleit, S.; Pretzsch, H. Structure and Ecosystem Services of Small-Leaved Lime (Tilia cordata Mill.) and Black Locust (Robinia pseudoacacia L.) in Urban Environments. Urban For. Urban Green. 2015, 14, 1110–1121. [Google Scholar] [CrossRef]
- Zang, H.; Lei, X.; Zeng, W. Height-Diameter Equations for Larch Plantations in Northern and Northeastern China: A Comparison of the Mixed-Effects, Quantile Regression and Generalized Additive Models. For. Int. J. For. Res. 2016, 89, 434–445. [Google Scholar] [CrossRef] [Green Version]
- McPherson, E.G.; van Doorn, N.S.; Peper, P.J. Urban Tree Database and Allometric Equations; General Technical Report PSW-GTR-253; US Department of Agriculture, Forest Service, Pacific Southwest Research Station: Albany, CA, USA, 2016; 86p.
- Shoda, T.; Imanishi, J.; Shibata, S. Growth Characteristics and Growth Equations of the Diameter at Breast Height Using Tree Ring Measurements of Street Trees in Kyoto City, Japan. Urban For. Urban Green. 2020, 49, 126627. [Google Scholar] [CrossRef]
- Yang, J.; Li, X.; Li, S.; Liang, H.; Lu, H. The Woody Plant Diversity and Landscape Pattern of Fine-Resolution Urban Forest along a Distance Gradient from Points of Interest in Qingdao. Ecol. Indic. 2021, 122, 107326. [Google Scholar] [CrossRef]
- Preuhsler, T. Ertragskundliche Merkmale Oberbayerischer Bergmischwald-Verjüngungsbestände Auf Kalkalpinen Standorten Im Forstamt Kreuth. Forstwiss. Centralblatt 1981, 100, 313–345. [Google Scholar] [CrossRef]
- Koenker, R.; Portnoy, S.; Ng, P.T.; Melly, B.; Zeileis, A.; Grosjean, P.; Saad, Y.; Chernozhukov, V.; Fernandez-Val, I. Quantreg: Quantile Regression. In R Package; Version 5.05; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: http://CRAN.R-project.org/package=quantreg (accessed on 22 July 2022).
- Koenker, R.; Hallock, K.F. Quantile Regression. J. Econ. Perspect. 2001, 15, 143–156. [Google Scholar] [CrossRef]
- Rust, S. Analysis of Regional Variation of Height Growth and Slenderness in Populations of Six Urban Tree Species Using a Quantile Regression Approach. Urban For. Urban Green. 2014, 13, 336–343. [Google Scholar] [CrossRef]
- Vogt, J.; Hauer, R.J.; Fischer, B.C. The Costs of Maintaining and Not Maintaining the Urban Forest: A Review of the Urban Forestry and Arboriculture Literature. Arboric. Urban For. 2015, 41, 293–323. [Google Scholar] [CrossRef]
- Peper, P.J.; McPherson, E.G.; Mori, S.M. Equations for Predicting Diameter, Height, Crown Width, and Leaf Area of San Joaquin Valley Street Trees. Arboric. Urban For. 2001, 27, 306–317. [Google Scholar] [CrossRef]
- Zheng, J.; Zang, H.; Yin, S.; Sun, N.; Zhu, P.; Han, Y.; Kang, H.; Liu, C. Modeling Height-Diameter Relationship for Artificial Monoculture Metasequoia Glyptostroboides in Sub-Tropic Coastal Megacity Shanghai, China. Urban For. Urban Green. 2018, 34, 226–232. [Google Scholar] [CrossRef]
- Foli, E.; Alder, D.; Miller, H.; Swaine, M. Modelling Growing Space Requirements for Some Tropical Forest Tree Species. For. Ecol. Manag. 2003, 173, 79–88. [Google Scholar] [CrossRef]
- Schmucker, J.; Uhl, E.; Steckel, M.; Pretzsch, H. Crown Allometry and Growing Space Requirements of Four Rare Domestic Tree Species Compared to Oak and Beech: Implications for Adaptive Forest Management. Eur. J. For. Res. 2022, 141, 587–604. [Google Scholar] [CrossRef]
- Zhang, W.-P.; Zhao, L.; Larjavaara, M.; Morris, E.C.; Sterck, F.J.; Wang, G.-X. Height-Diameter Allometric Relationships for Seedlings and Trees across China. Acta Oecologica 2020, 108, 103621. [Google Scholar] [CrossRef]
- Devisscher, T.; Ordóñez-Barona, C.; Dobbs, C.; Baptista, M.D.; Navarro, N.M.; Aguilar, L.A.O.; Perez, J.F.C.; Mancebo, Y.R.; Escobedo, F.J. Urban Forest Management and Governance in Latin America and the Caribbean: A Baseline Study of Stakeholder Views. Urban For. Urban Green. 2022, 67, 127441. [Google Scholar] [CrossRef]
- John Barker, E.; Andy Kenney, W. Urban Forest Management in Small Ontario Municipalities. For. Chron. 2012, 88, 118–123. [Google Scholar] [CrossRef]
- Stoffberg, G.; Van Rooyen, M.; Van der Linde, M.; Groeneveld, H. Predicting the Growth in Tree Height and Crown Size of Three Street Tree Species in the City of Tshwane, South Africa. Urban For. Urban Green. 2008, 7, 259–264. [Google Scholar] [CrossRef]
- Chreptun, C. Kronenstruktureigenschaften von Linden Und Robinien in München: Anwendungen Des Terrestrischen Laserscannings. PhD Thesis, Technische Universität München, München, Germany, 2015. [Google Scholar]
- West, G.B.; Enquist, B.J.; Brown, J.H. A General Quantitative Theory of Forest Structure and Dynamics. Proc. Natl. Acad. Sci. USA 2009, 106, 7040–7045. [Google Scholar] [CrossRef] [Green Version]
- Forrester, D.I.; Benneter, A.; Bouriaud, O.; Bauhus, J. Diversity and Competition Influence Tree Allometric Relationships–Developing Functions for Mixed-Species Forests. J. Ecol. 2017, 105, 761–774. [Google Scholar] [CrossRef] [Green Version]
- Franceschi, E.; Moser-Reischl, A.; Rahman, M.A.; Pauleit, S.; Pretzsch, H.; Rötzer, T. Crown Shapes of Urban Trees-Their Dependences on Tree Species, Tree Age and Local Environment, and Effects on Ecosystem Services. Forests 2022, 13, 748. [Google Scholar] [CrossRef]
- Purves, D.W.; Lichstein, J.W.; Pacala, S.W. Crown Plasticity and Competition for Canopy Space: A New Spatially Implicit Model Parameterized for 250 North American Tree Species. PLoS ONE 2007, 2, e870. [Google Scholar] [CrossRef] [PubMed]
- Sanders, J.; Grabosky, J.; Cowie, P. Establishing Maximum Size Expectations for Urban Trees with Regard to Designed Space. Arboric. Urban For. 2013, 39, 68–73. [Google Scholar] [CrossRef]
- Bolund, P.; Hunhammar, S. Ecosystem Services in Urban Areas. Ecol. Econ. 1999, 29, 293–301. [Google Scholar] [CrossRef]
- Cysneiros, V.C.; de Souza, F.C.; Gaui, T.D.; Pelissari, A.L.; Orso, G.A.; Machado, S.D.A.; de Carvalho, D.C.; Silveira-Filho, T.B. Integrating Climate, Soil and Stand Structure into Allometric Models: An Approach of Site-Effects on Tree Allometry in Atlantic Forest. Ecol. Indic. 2021, 127, 107794. [Google Scholar] [CrossRef]
- Sharma, R.P.; Vacek, Z.; Vacek, S. Individual Tree Crown Width Models for Norway Spruce and European Beech in Czech Republic. For. Ecol. Manag. 2016, 366, 208–220. [Google Scholar] [CrossRef]
- Coombes, A.; Martin, J.; Slater, D. Defining the Allometry of Stem and Crown Diameter of Urban Trees. Urban For. Urban Green. 2019, 44, 126421. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Valladares, F. Tolerance to Shade, Drought, and Waterlogging of Temperate Northern Hemisphere Trees and Shrubs. Ecol. Monogr. 2006, 76, 521–547. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | n | Tree DBH (cm) | Tree Height (m) | Crown Width (m) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Min | Max | Mean | Sd | Min | Max | Mean | Sd | Min | Max | Mean | Sd | ||
| Platanus × acerifolia | 2630 | 7.5 | 100.4 | 30.2 | 11.4 | 3.0 | 27.0 | 10.8 | 3.2 | 0.5 | 21.3 | 7.6 | 3.0 |
| Fraxinus chinensis | 1057 | 10.0 | 47.1 | 21.0 | 7.2 | 3.0 | 16.0 | 8.3 | 2.5 | 1.0 | 14.1 | 6.0 | 2.3 |
| Prunus cerasus | 85 | 5.9 | 22.0 | 14.2 | 3.5 | 2.0 | 6.0 | 4.4 | 1.1 | 0.5 | 6.0 | 2.9 | 0.9 |
| Ginkgo biloba | 418 | 6.2 | 33.8 | 18.4 | 4.6 | 3.0 | 15.0 | 8.4 | 2.0 | 1.0 | 9.2 | 3.7 | 1.5 |
| Koelreuteria paniculata | 179 | 10.0 | 36.1 | 20.7 | 5.0 | 4.1 | 13.5 | 8.5 | 1.6 | 2.0 | 9.8 | 5.2 | 1.7 |
| Ailanthus altissima | 73 | 9.8 | 46.8 | 27.4 | 8.3 | 5.0 | 17.5 | 11.7 | 2.6 | 2.5 | 10.5 | 6.6 | 1.8 |
| Styphnolobium japonicum | 287 | 7.8 | 46.4 | 23.4 | 7.6 | 3.5 | 12.8 | 7.7 | 1.9 | 1.5 | 13.5 | 6.7 | 1.9 |
| Cercis chinensis | 85 | 9.0 | 18.7 | 13.3 | 2.5 | 5.0 | 8.0 | 6.7 | 0.9 | 1.0 | 7.6 | 4.3 | 1.7 |
| Pinus thunbergii | 163 | 10.2 | 36.8 | 17.9 | 4.7 | 2.5 | 10.0 | 5.7 | 1.5 | 1.5 | 7.0 | 4.3 | 1.0 |
| Robinia pseudoacacia | 83 | 15.9 | 40.1 | 24.8 | 6.8 | 5.1 | 11.0 | 7.2 | 1.3 | 3.2 | 8.0 | 4.5 | 1.2 |
| Metasequoia glyptostroboides | 64 | 12.7 | 59.5 | 31.7 | 12.6 | 5.7 | 18.2 | 17.9 | 4.9 | 1.5 | 6.5 | 4.0 | 1.3 |
| Cedrus deodara | 184 | 7.0 | 66.3 | 39.5 | 9.0 | 4.5 | 23.0 | 12.9 | 3.3 | 3.2 | 27.7 | 8.6 | 2.7 |
| Populus spp. | 46 | 16.3 | 70.0 | 38.9 | 13.9 | 5.0 | 18.0 | 12.1 | 3.7 | 2.0 | 13.0 | 7.7 | 2.5 |
| Firmiana simplex | 221 | 12.5 | 63.4 | 34.4 | 9.3 | 5.0 | 15.5 | 12.4 | 1.6 | 3.0 | 14.8 | 8.4 | 2.3 |
| Salix matsudana | 73 | 17.7 | 87.6 | 38.2 | 12.5 | 4.5 | 16.0 | 10.3 | 3.0 | 3.0 | 10.3 | 6.5 | 1.9 |
| Species | a | Std. Err. (a) | t (a) | p (a) | α | Std. Err. (α) | t (α) | p (α) |
|---|---|---|---|---|---|---|---|---|
| Platanus × acerifolia | 0.6782 | 0.0252 | 26.9184 | 0.0000 | 0.3485 | 0.0152 | 22.9473 | 0.0000 |
| Fraxinus chinensis | 0.3157 | 0.0467 | 6.7553 | 0.0000 | 0.5624 | 0.0336 | 16.7310 | 0.0000 |
| Prunus cerasus | 0.2898 | 0.3288 | 0.8813 | 0.3807 | 0.4075 | 0.2744 | 1.4848 | 0.1414 |
| Ginkgo biloba | 0.4534 | 0.0757 | 5.9876 | 0.0000 | 0.4780 | 0.0604 | 7.9115 | 0.0000 |
| Koelreuteria paniculata | 0.3761 | 0.1709 | 2.2012 | 0.0290 | 0.4990 | 0.1324 | 3.7678 | 0.0002 |
| Ailanthus altissima | 0.7393 | 0.3579 | 2.0656 | 0.0425 | 0.3280 | 0.2646 | 1.2310 | 0.2191 |
| Styphnolobium japonicum | 0.2312 | 0.1355 | 1.707 | 0.0889 | 0.5696 | 0.0965 | 5.9060 | 0.0000 |
| Cercis chinensis | 0.9031 | 0.0329 | 27.4725 | 0.0000 | 0.0000 | 0.0288 | 0.0000 | 1.0000 |
| Pinus thunbergii | 0.4335 | 0.1393 | 3.1125 | 0.0022 | 0.3684 | 0.1149 | 3.2077 | 0.0016 |
| Robinia pseudoacacia | 0.6588 | 0.1965 | 3.3536 | 0.0012 | 0.2242 | 0.1452 | 1.54404 | 0.1265 |
| Metasequoia glyptostroboides | 0.8228 | 0.0569 | 14.4566 | 0.0000 | 0.3566 | 0.0441 | 8.0891 | 0.0000 |
| Cedrus deodara | 0.6504 | 0.2020 | 3.2195 | 0.0015 | 0.3943 | 0.1265 | 3.1161 | 0.0021 |
| Populus spp. | 1.7146 | 0.0995 | 17.2282 | 0.0000 | −0.3111 | 0.0567 | −5.4874 | 0.0000 |
| Firmiana simplex | 1.1410 | 0.0390 | 29.2232 | 0.0000 | 0.0186 | 0.0256 | 0.7249 | 0.4693 |
| Salix matsudana | 0.8294 | 0.2680 | 3.0949 | 0.0028 | 0.2187 | 0.1771 | 1.2346 | 0.2211 |
| Species | a | Std. Err. (a) | t (a) | p (a) | α | Std. Err. (α) | t (α) | p (α) |
|---|---|---|---|---|---|---|---|---|
| Platanus × acerifolia | 0.0922 | 0.0256 | 3.5993 | 0.0003 | 0.6588 | 0.0140 | 46.9119 | 0.0000 |
| Fraxinus chinensis | −0.0204 | 0.0495 | −0.4117 | 0.6806 | 0.7253 | 0.0312 | 23.2426 | 0.0000 |
| Prunus cerasus | 0.6532 | 0.1558 | 4.1931 | 0.0001 | 0.0000 | 0.1424 | 0.0000 | 1.0000 |
| Ginkgo biloba | −0.1733 | 0.1763 | −0.9832 | 0.3261 | 0.7300 | 0.1362 | 5.3613 | 0.0000 |
| Koelreuteria paniculata | 0.0884 | 0.2146 | 0.4117 | 0.6811 | 0.5908 | 0.1618 | 3.6513 | 0.0003 |
| Ailanthus altissima | 0.5570 | 0.2564 | 2.1728 | 0.0331 | 0.2838 | 0.1669 | 1.7001 | 0.0935 |
| Styphnolobium japonicum | 0.1396 | 0.0616 | 2.2674 | 0.0241 | 0.5875 | 0.0486 | 12.0870 | 0.0000 |
| Cercis chinensis | −2.1006 | 0.1965 | −10.6923 | 0.0000 | 2.5117 | 0.1772 | 14.1786 | 0.0000 |
| Pinus thunbergii | 0.2942 | 0.0837 | 3.5140 | 0.0006 | 0.3632 | 0.0621 | 5.8534 | 0.0000 |
| Robinia pseudoacacia | −0.2550 | 0.1623 | −1.5708 | 0.1201 | 0.7357 | 0.1124 | 6.5477 | 0.0000 |
| Metasequoia glyptostroboides | 0.5476 | 0.2014 | 2.7187 | 0.0085 | 0.1445 | 0.1258 | 1.1488 | 0.2550 |
| Cedrus deodara | 1.0899 | 0.4167 | 2.6155 | 0.0097 | 0.0104 | 0.2593 | 0.0401 | 0.9681 |
| Populus spp. | 0.3172 | 0.5334 | 0.5947 | 0.5551 | 0.4618 | 0.3089 | 1.4951 | 0.1420 |
| Firmiana simplex | −0.0755 | 0.0667 | −1.1328 | 0.2586 | 0.7274 | 0.0480 | 15.1501 | 0.0000 |
| Salix matsudana | 0.5894 | 0.2315 | 2.5457 | 0.0131 | 0.2523 | 0.1355 | 1.8617 | 0.0668 |
| Species | a | Std. Err. (a) | t (a) | p (a) | α | Std. Err. (α) | t (α) | p (α) |
|---|---|---|---|---|---|---|---|---|
| Platanus × acerifolia | 0.0794 | 0.0512 | 1.5508 | 0.1211 | 1.3175 | 0.0281 | 46.9086 | 0.0000 |
| Fraxinus chinensis | −0.1457 | 0.0991 | −1.4705 | 0.1417 | 1.4506 | 0.06241 | 23.2425 | 0.0000 |
| Prunus cerasus | 1.2015 | 0.3116 | 3.8564 | 0.0002 | 0.0000 | 0.2849 | 0.0000 | 1.0000 |
| Ginkgo biloba | −0.4516 | 0.3526 | −1.2807 | 0.2010 | 1.4601 | 0.2723 | 5.3613 | 0.0000 |
| Koelreuteria paniculata | 0.0718 | 0.4292 | 0.1673 | 0.8673 | 1.1816 | 0.3236 | 3.6513 | 0.0003 |
| Ailanthus altissima | 1.0091 | 0.5127 | 1.9681 | 0.0530 | 0.5676 | 0.3338 | 1.7001 | 0.0935 |
| Styphnolobium japonicum | 0.1743 | 0.1232 | 1.4156 | 0.1580 | 1.1750 | 0.0972 | 12.0870 | 0.0000 |
| Cercis chinensis | −4.3061 | 0.3929 | −10.9593 | 0.0000 | 5.0234 | 0.3543 | 14.1786 | 0.0000 |
| Pinus thunbergii | 0.4834 | 0.1674 | 2.8874 | 0.0044 | 0.7265 | 0.1241 | 5.8534 | 0.0000 |
| Robinia pseudoacacia | −0.6149 | 0.3247 | −1.8940 | 0.0618 | 1.4715 | 0.2247 | 6.5477 | 0.0000 |
| Metasequoia glyptostroboides | 0.9903 | 0.4028 | 2.4583 | 0.0168 | 0.2889 | 0.2515 | 1.1488 | 0.2550 |
| Cedrus deodara | 2.0749 | 0.8335 | 2.4896 | 0.0137 | 0.0208 | 0.5186 | 0.0401 | 0.9681 |
| Populus spp. | 0.5295 | 1.0668 | 0.4964 | 0.6221 | 0.9236 | 0.6178 | 1.4951 | 0.1420 |
| Firmiana simplex | −0.2559 | 0.1333 | −1.9198 | 0.0562 | 1.4547 | 0.0960 | 15.1501 | 0.0000 |
| Salix matsudana | 1.0739 | 0.4631 | 2.3191 | 0.0233 | 0.5046 | 0.2710 | 1.8617 | 0.0668 |
| Species | a | Std. Err. (a) | t (a) | p (a) | α | Std. Err. (α) | t (α) | p (α) |
|---|---|---|---|---|---|---|---|---|
| Platanus × acerifolia | 0.4342 | 0.1349 | 3.2196 | 0.0013 | 1.7755 | 0.0834 | 21.2980 | 0.0000 |
| Fraxinus chinensis | −0.5079 | 0.2019 | −2.5152 | 0.0120 | 2.3664 | 0.1372 | 17.2487 | 0.0000 |
| Prunus cerasus | 1.0937 | 0.5769 | 1.8958 | 0.0615 | 0.6477 | 0.4929 | 1.3141 | 0.1925 |
| Ginkgo biloba | −0.6534 | 0.3223 | −2.0272 | 0.0433 | 2.2636 | 0.2384 | 9.4956 | 0.0000 |
| Koelreuteria paniculata | −0.4227 | 0.4498 | −0.9397 | 0.3486 | 2.1490 | 0.3157 | 6.8064 | 0.0000 |
| Ailanthus altissima | 1.5308 | 0.4448 | 3.4416 | 0.0010 | 0.8810 | 0.2793 | 3.1545 | 0.0024 |
| Styphnolobium japonicum | 0.2802 | 0.1951 | 1.4363 | 0.1520 | 1.6605 | 0.1218 | 13.6326 | 0.0000 |
| Cercis chinensis | −3.4541 | 0.6360 | −5.4313 | 0.0000 | 4.9176 | 0.5434 | 9.0502 | 0.0000 |
| Pinus thunbergii | 0.2878 | 0.2320 | 1.2405 | 0.2166 | 1.3851 | 0.1709 | 8.1035 | 0.0000 |
| Robinia pseudoacacia | −1.0522 | 0.5781 | −1.8200 | 0.0725 | 2.2688 | 0.4225 | 5.3698 | 0.0000 |
| Metasequoia glyptostroboides | 1.1750 | 0.2344 | 5.0120 | 0.0000 | 0.9454 | 0.1416 | 6.6779 | 0.0000 |
| Cedrus deodara | 2.6802 | 1.0283 | 2.6064 | 0.0099 | 0.3385 | 0.6155 | 0.5499 | 0.5831 |
| Populus spp. | 1.5983 | 0.2577 | 6.2031 | 0.0000 | 0.8401 | 0.1493 | 5.6266 | 0.0000 |
| Firmiana simplex | 0.7884 | 0.3483 | 2.2639 | 0.0246 | 1.4006 | 0.2350 | 6.0384 | 0.0000 |
| Salix matsudana | 1.9686 | 0.4972 | 3.9589 | 0.0002 | 0.6290 | 0.2888 | 2.1779 | 0.0327 |
| Species | Shade Tolerance Index | Drought Tolerance Index | Waterlogging Tolerance Index |
|---|---|---|---|
| Platanus × acerifolia | 3 | 3.5 | 2 |
| Ginkgo biloba | 1.34 ± 0.33 | 3.99 ± 0.47 | 1.13 ± 0.27 |
| Pinus thunbergii | 1.35 | 4.03 ± 0.18 | 1.02 ± 0.02 |
| Cedrus deodara | 2.59 ± 0.41 | 3.85 ± 0.15 | 1.02 ± 0.02 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.; Zhang, M.; Zhang, J.; Lu, H.; Hauer, R.J. Allometric Growth of Common Urban Tree Species in Qingdao City of Eastern China. Forests 2023, 14, 472. https://doi.org/10.3390/f14030472
Yang J, Zhang M, Zhang J, Lu H, Hauer RJ. Allometric Growth of Common Urban Tree Species in Qingdao City of Eastern China. Forests. 2023; 14(3):472. https://doi.org/10.3390/f14030472
Chicago/Turabian StyleYang, Jinming, Mengfan Zhang, Jian Zhang, Huicui Lu, and Richard J. Hauer. 2023. "Allometric Growth of Common Urban Tree Species in Qingdao City of Eastern China" Forests 14, no. 3: 472. https://doi.org/10.3390/f14030472
APA StyleYang, J., Zhang, M., Zhang, J., Lu, H., & Hauer, R. J. (2023). Allometric Growth of Common Urban Tree Species in Qingdao City of Eastern China. Forests, 14(3), 472. https://doi.org/10.3390/f14030472