


Soil C, N, P, K and Enzymes Stoichiometry of an Endangered Tree Species, Parashorea chinensis of Different Stand Ages Unveiled Soil Nutrient Limitation Factors


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling Site
2.2. Experimental Design and Soil Sampling
2.3. Soil Physicochemical Analyses
2.4. Soil Microbial Biomass and Enzyme Activities Assays
2.5. Data Statistics and Analysis
3. Results
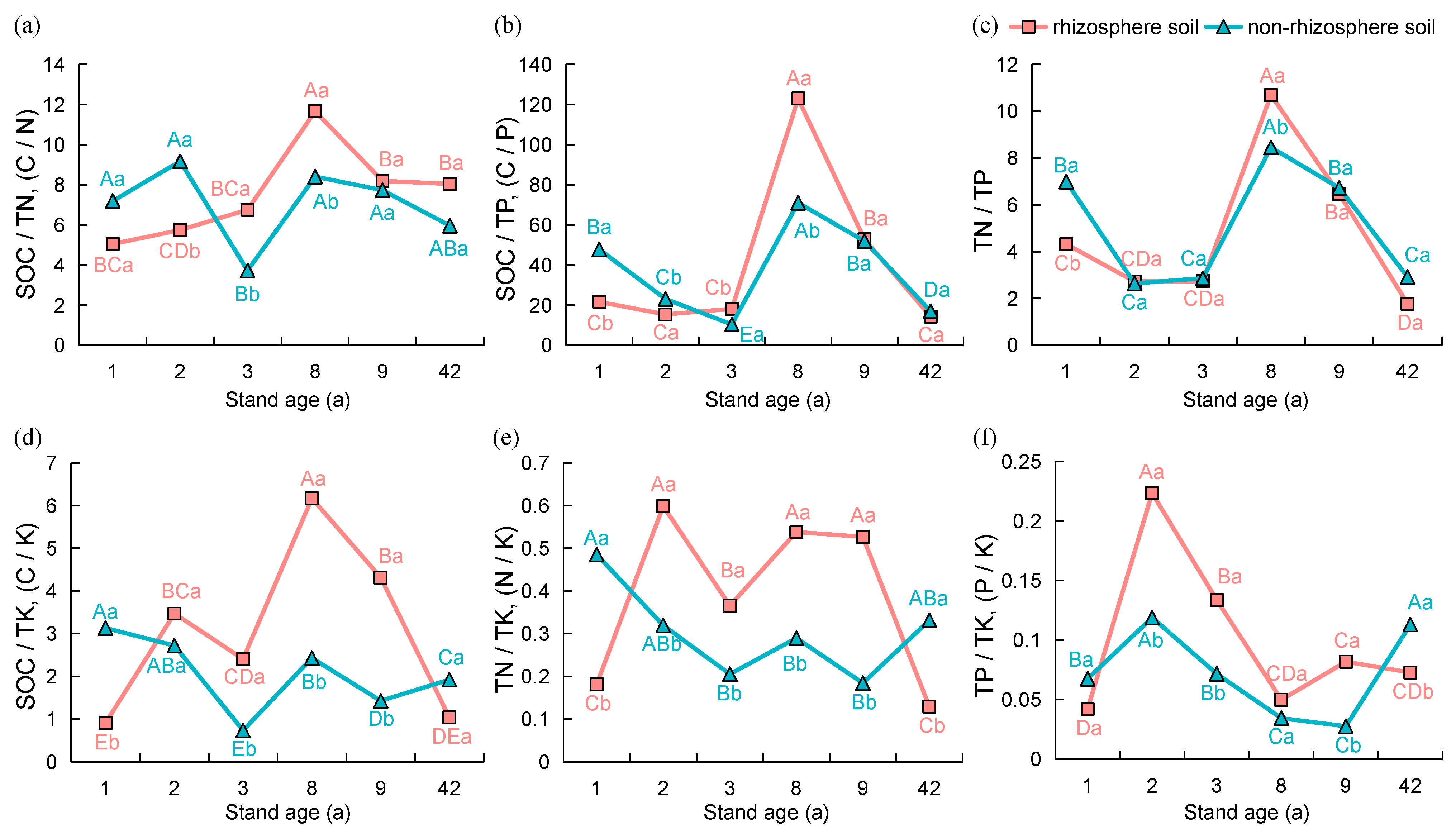
3.1. Physicochemical Properties and Nutrient Stoichiometric Ratios of Rhizosphere and Non-Rhizosphere Soils under Different Age Stands
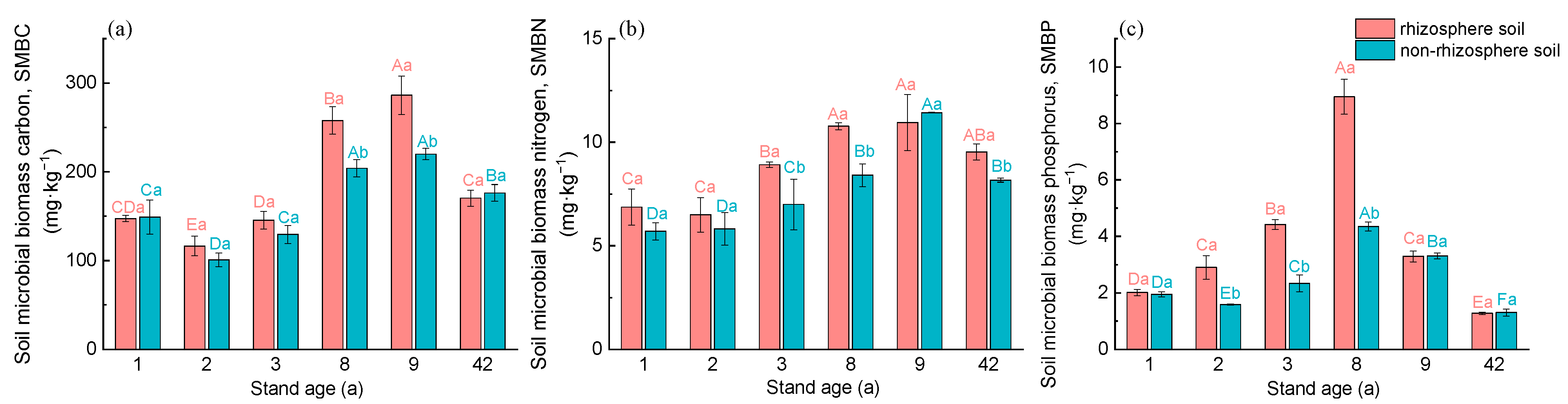
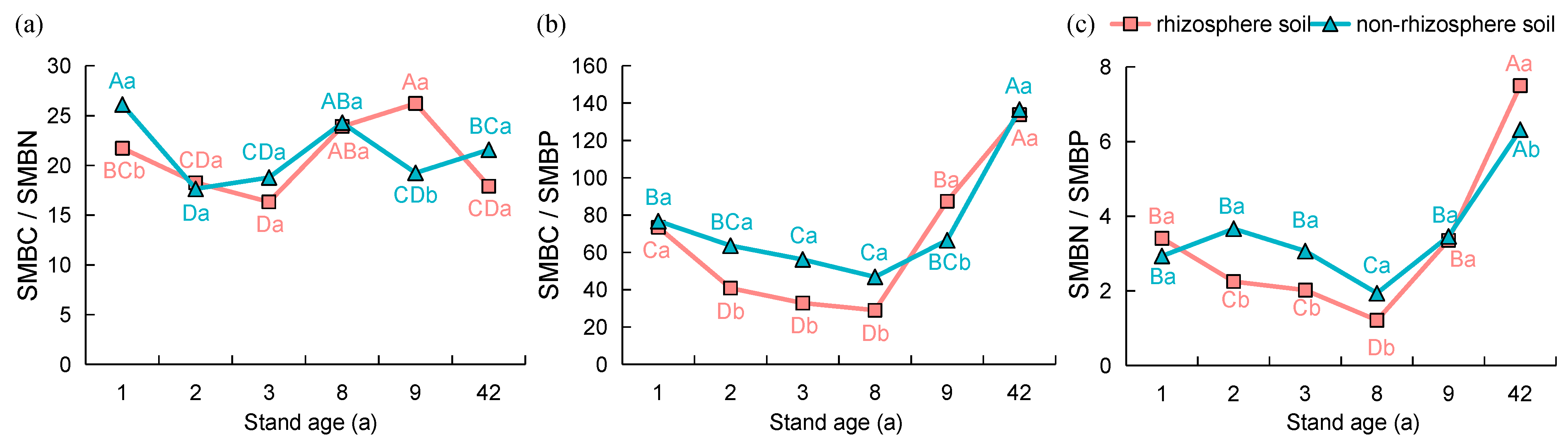
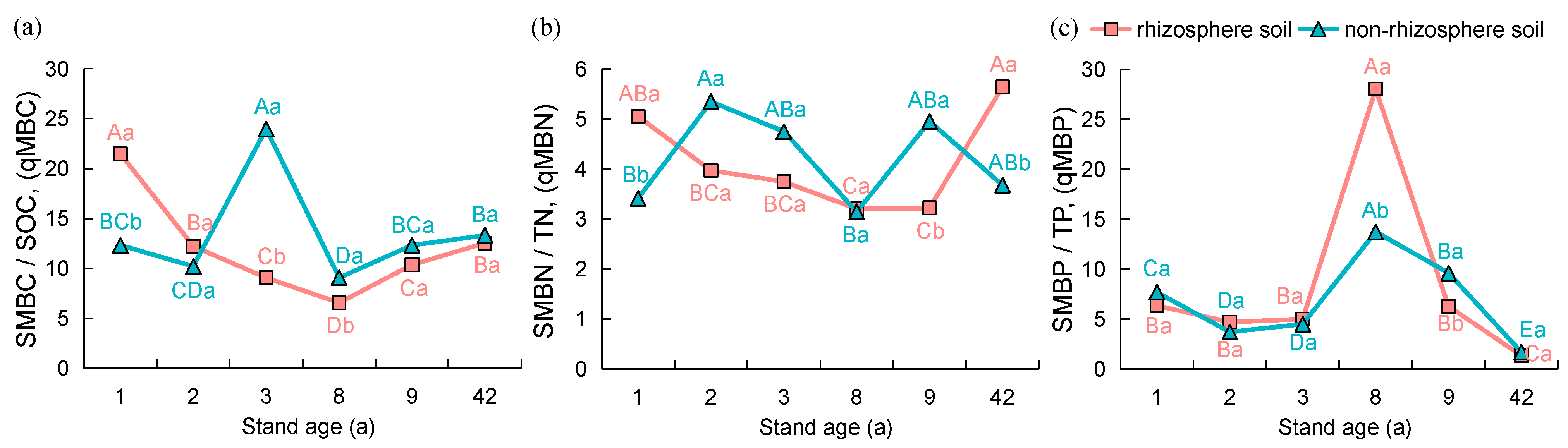
3.2. Microbial Biomass and Stoichiometric Ratios in the Rhizosphere and Non-Rhizosphere Soils under Different Age Stands
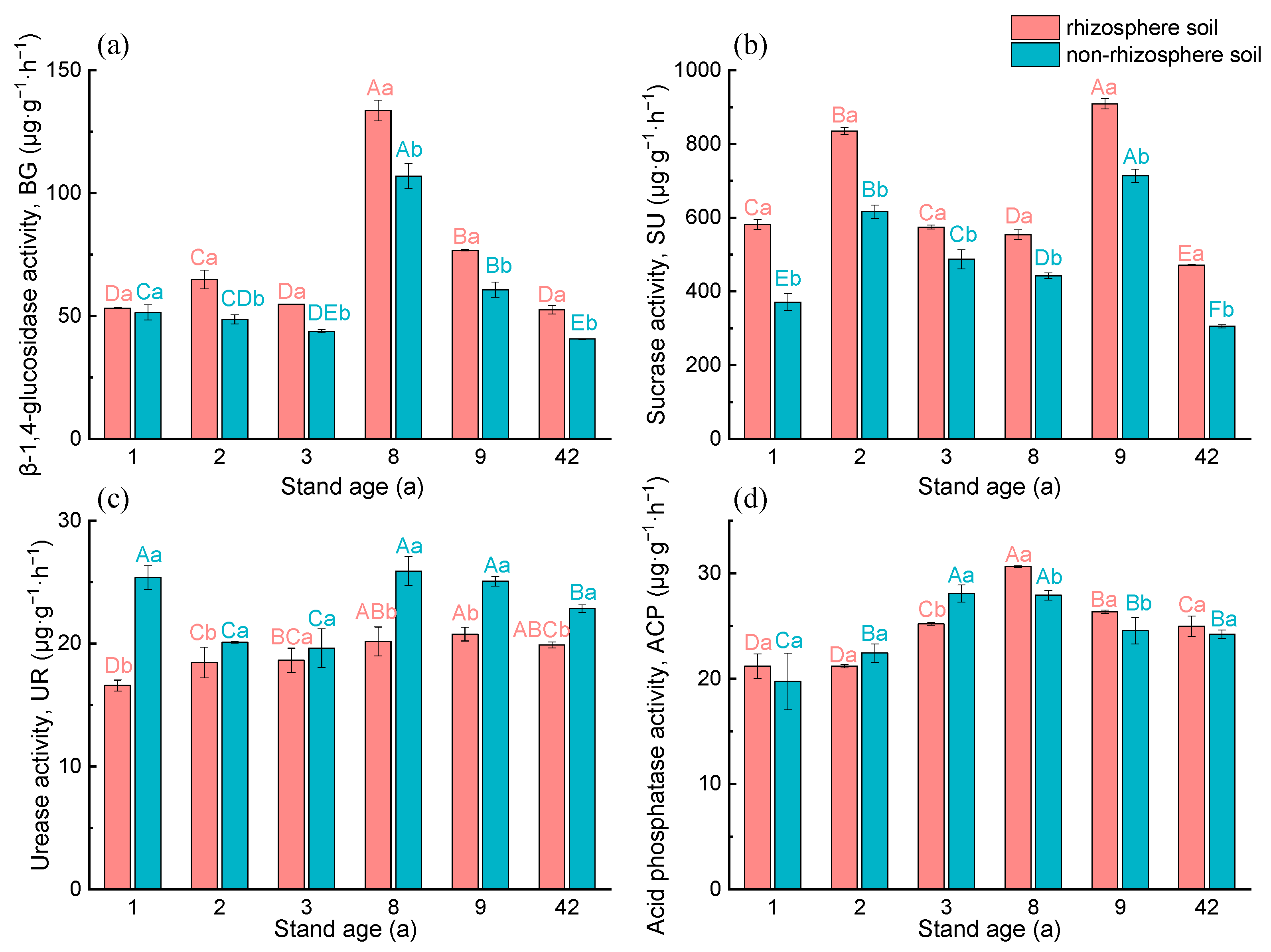
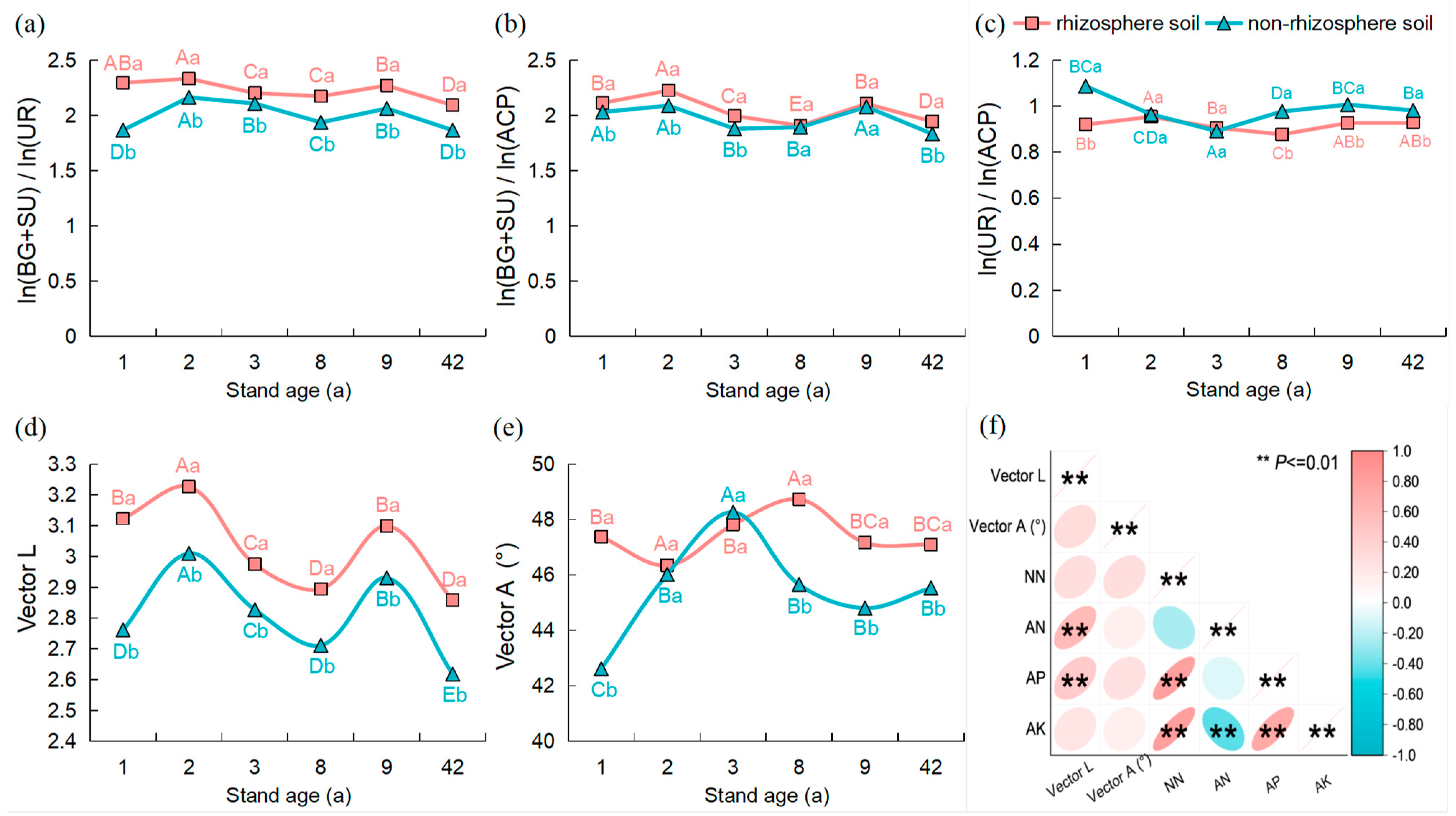
3.3. Enzyme Activity and Enzyme Stoichiometry Ratios in the Rhizosphere and Non-Rhizosphere Soils under Different Age Stands
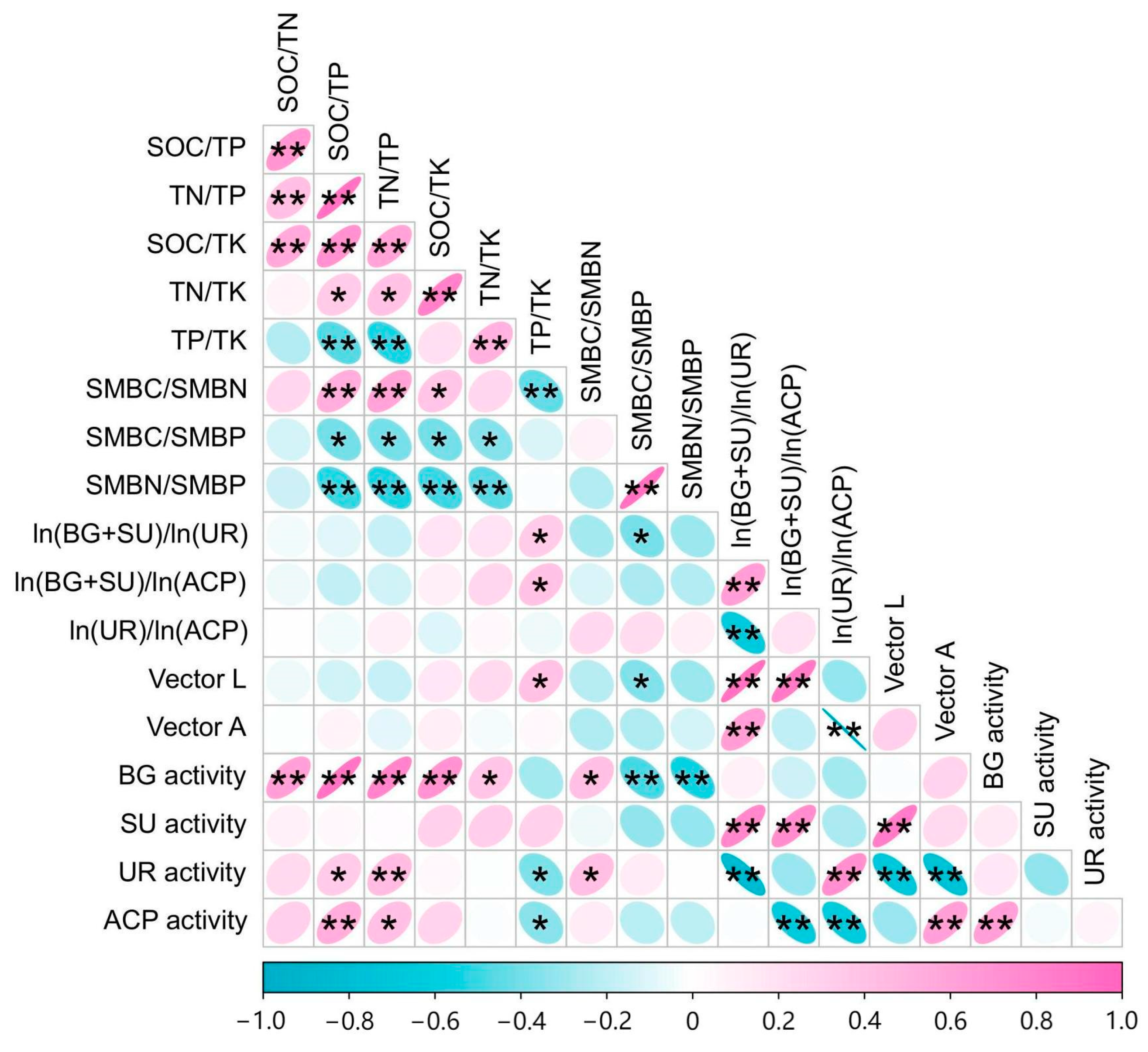
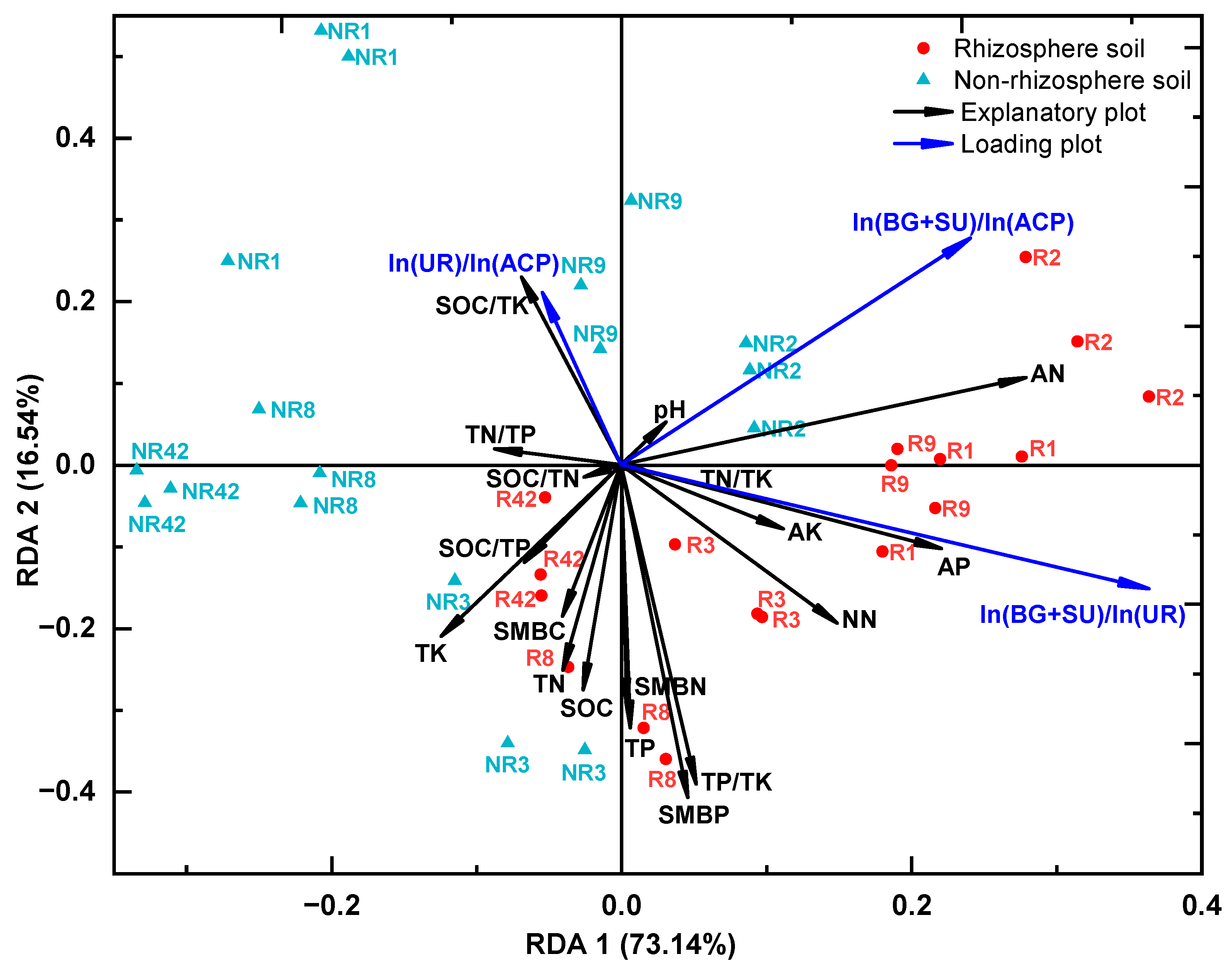
3.4. Relationships between Soil Nutrients, Microbial Biomass, and Enzyme Activity and Their Stoichiometric Ratios in the Rhizosphere and Non-Rhizosphere Soils
4. Discussion
4.1. Ecological Stoichiometry of Soil Nutrients, Microbial Biomass, and Enzyme Activity in Rhizosphere and Non-Rhizosphere Soils
4.2. Effect of Stand Age on Soil Nutrients, Microbial Biomass, and Enzyme Activity
4.3. Effect of Stand Age on Soil Nutrients, Microbial Biomass, and Extracellular Enzyme Stoichiometry Ratios
4.4. Correlation of C, N, and P Stoichiometric Ratios between “Nutrient-Microbial Biomass-Enzyme Activity” in Soils
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Das, S.C.; Alam, M.S.; Hossain, M.A. Diversity and structural composition of species in dipterocarp forests: A study from fasiakhali wildlife sanctuary, bangladesh. J. For. Res. 2018, 29, 1241–1249. [Google Scholar] [CrossRef]
- Guo, J.H. National class i protected plant Parashorea chinensis and its family. Biol. Teach. 2000, 43, 37–38. [Google Scholar]
- Qin, H.N.; Yang, Y.; Dong, S.Y.; He, Q.; Jia, Y.; Zhao, L.N.; Yu, S.X.; Liu, H.Y.; Liu, B. Threatened species list of China’s higher plants. Biodivers. Sci. 2017, 25, 696–744. [Google Scholar] [CrossRef] [Green Version]
- Ming, Y. The adjusted “list of national key protected wild plants” was officially announced. Green China 2021, 581, 74–79. [Google Scholar]
- Ismail, S.A.; Buser, A.; Shaanker, R.U.; Ravikanth, G.; Ghazoul, J.; Kettle, C.J. Development of polymorphic microsatellite markers for the critically endangered and endemic indian dipterocarp, Vateria indica L. (Dipterocarpaceae). Conserv. Genet. Resour. 2013, 5, 465–467. [Google Scholar] [CrossRef] [Green Version]
- Maycock, C.R.; Kettle, C.J.; Khoo, E.; Pereira, J.T.; Sugau, J.B.; Nilus, R.; Ong, R.C.; Amaludin, N.A.; Newman, M.F.; Burslem, D. A revised conservation assessment of dipterocarps in sabah. Biotropica 2012, 44, 649–657. [Google Scholar] [CrossRef] [Green Version]
- Nawi, L.; Suratman, M.N.; Siti, Z.M.T. Conservation of the Critically Endangered Tree Species Dipterocarpus Semivestitus in Malaysia; IEEE: Ney York, NY, USA, 2013; pp. 225–228. [Google Scholar]
- Tian, Z.Z.; Zeng, P.; Lu, X.Y.; Zhou, T.G.; Han, Y.W.; Peng, Y.M.; Xiao, Y.X.; Zhou, B.T.; Liu, X.; Zhang, Y.T.; et al. Thirteen dipterocarpoideae genomes provide insights into their evolution and borneol biosynthesis. Plant Commun. 2022, 3, 100464. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, H.L.; Chen, Y.K.; Nizamani, M.M.; Zhou, Q.; Su, X.T. Analyses of community stability and inter-specific associations between a plant species with extremely small populations (hopea hainanensis) and its associated species. Front. Ecol. Evol. 2022, 10, 922829. [Google Scholar] [CrossRef]
- Filipiak, M.; Filipiak, Z.M. Application of ionomics and ecological stoichiometry in conservation biology: Nutrient demand and supply in a changing environment. Biol. Conserv. 2022, 272, 109622. [Google Scholar] [CrossRef]
- Shukla, A.; Mehrotra, R.C.; Guleria, J.S. Emergence and extinction of dipterocarpaceae in western india with reference to climate change: Fossil wood evidences. J. Earth Syst. Sci. 2013, 122, 1373–1386. [Google Scholar] [CrossRef] [Green Version]
- Velden, V.D.; Ferry Slik, J.W.; Hu, Y.H.; Lan, G.; Lin, L.; Deng, X.; Poorter, L. Monodominance of Parashorea chinensis on fertile soils in a chinese tropical rain forest. J. Trop. Ecol. 2014, 30, 311–322. [Google Scholar] [CrossRef]
- Meng, L.Z.; Zhang, J.L.; Cao, K.F.; Xu, Z.F. Diurnal changes of photosynthetic characteristics and chlorophyll fluorescence in canopy leaves of four diptocarp species under ex-situ conservation. Acta Phytoecol. Sin. 2005, 29, 976–984. [Google Scholar]
- Xiao, Y.X.; Liu, G.Y. Dipterocarpaceae plants ex-situ conservation and resources exploitation in Xishuangbanna tropical botanical garden (xtbg). Guihaia 2021, 41, 843–852. [Google Scholar]
- Su, L.; Du, H.; Zeng, F.P.; Peng, W.X.; Rizwan, M.; Nunez-Delgado, A.; Zhou, Y.Y.; Song, T.Q.; Wang, H. Soil and fine roots ecological stoichiometry in different vegetation restoration stages in a karst area, southwest China. J. Environ. Manag. 2019, 252, 109694. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, L.M.; Chen, J.; Feng, L.; Li, F.B.; Yu, L.F. Functional diversity of plant communities in relationship to leaf and soil stoichiometry in karst areas of south west China. Forests 2022, 13, 864. [Google Scholar] [CrossRef]
- Amazonas, N.T.; Martinelli, L.A.; Piccolo, M.D.; Rodrigues, R.R. Nitrogen dynamics during ecosystem development in tropical forest restoration. For. Ecol. Manag. 2011, 262, 1551–1557. [Google Scholar] [CrossRef]
- Guo, Z.M.; Zhang, X.Y.; Green, S.M.; Dungait, J.; Wen, X.F.; Quine, T.A. Soil enzyme activity and stoichiometry along a gradient of vegetation restoration at the karst critical zone observatory in southwest China. Land Degrad. Dev. 2019, 30, 1916–1927. [Google Scholar] [CrossRef]
- Waal, D.B.V.D.; Elser, J.J.; Martiny, A.C.; Sterner, R.W.; Cotner, J.B. Editorial: Progress in ecological stoichiometry. Front. Microbiol. 2018, 9, 1957. [Google Scholar] [CrossRef] [Green Version]
- Sterner, R.W.; Elser, J.J. Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere; Princeton University Press: Princeton, NJ, USA, 2002; pp. 439–464. [Google Scholar]
- Qi, K.B.; Pang, X.Y.; Yang, B.; Bao, W.K. Soil carbon, nitrogen and phosphorus ecological stoichiometry shifts with tree species in subalpine plantations. PeerJ 2020, 8, e9702. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.H.; Ji, H.W.; Sun, N.X.; Tao, H.M.; Du, B.M.; Hui, D.F.; Liu, C.J. Imbalanced plant stoichiometry at contrasting geologic-derived phosphorus sites in subtropics: The role of microelements and plant functional group. Plant Soil 2018, 430, 113–125. [Google Scholar] [CrossRef]
- Wang, L.J.; Wang, P.; Sheng, M.Y.; Tian, J. Ecological stoichiometry and environmental influencing factors of soil nutrients in the karst rocky desertification ecosystem, southwest China. Glob. Ecol. Conserv. 2018, 16, e449. [Google Scholar] [CrossRef]
- Hu, X.Y.; Duan, A.G.; Zhang, J.G.; Du, H.L.; Zhang, X.Q.; Guo, W.F.; Sun, J.J. Stoichiometry of carbon, nitrogen, and phosphorus of chinese fir plantations in daqing mountain, guangxi. Acta Ecol. Sin. 2020, 40, 1207–1218. [Google Scholar]
- Güsewell, S. N: P ratios in terrestrial plants: Variation and functional significance. New Phytol. 2004, 164, 243–266. [Google Scholar] [CrossRef]
- Wu, P.; Zhou, H.; Cui, Y.C.; Zhao, W.J.; Hou, Y.J.; Tan, C.J.; Yang, G.N.; Ding, F.J. Stoichiometric characteristics of leaf, litter and soil during vegetation succession in maolan national nature reserve, guizhou, China. Sustainability 2022, 14, 16517. [Google Scholar] [CrossRef]
- Finn, D.; Kopittke, P.M.; Dennis, P.G.; Dalal, R.C. Microbial energy and matter transformation in agricultural soils. Soil Biol. Biochem. 2017, 111, 176–192. [Google Scholar] [CrossRef] [Green Version]
- Sinsabaugh, R.L.; Shah, J. Ecoenzymatic stoichiometry and ecological theory. Annu. Rev. Ecol. Evol. Syst. 2012, 43, 313–343. [Google Scholar] [CrossRef] [Green Version]
- Schimel, J.P.; Weintraub, M.N. The implications of exoenzyme activity on microbial carbon and nitrogen limitation in soil: A theoretical model. Soil Biol. Biochem. 2003, 35, 549–563. [Google Scholar] [CrossRef]
- Yu, Y.; Zheng, W.; Zhong, X.; Ying, B. Stoichiometric characteristics of carbon, nitrogen and phosphorus in zanthoxylum planispinum var. Dintanensis plantation of different ages. Agron. J. 2020, 113, 685–695. [Google Scholar]
- Wang, X.F.; Li, J.L.; Xing, G.T.; Mai, S.W.; Liu, W.J.; Jiang, Y.M.; Xu, W.X.; Yang, Q.; Yang, H.; Lu, J.L.; et al. Soil organic carbon distribution, enzyme activities, and the temperature sensitivity of a tropical rainforest in wuzhishan, hainan island. Forests 2022, 13, 1943. [Google Scholar] [CrossRef]
- Hill, B.H.; Elonen, C.M.; Jicha, T.M.; Cotter, A.M.; Trebitz, A.S.; Danz, N.P. Sediment microbial enzyme activity as an indicator of nutrient limitation in great lakes coastal wetlands. Freshw. Biol. 2006, 51, 1670–1683. [Google Scholar] [CrossRef]
- Chen, H.N.; Xiang, Y.; Yao, Z.X.; Zhang, Q.; Li, H.; Cheng, M. Stability of C:N:P stoichiometry in the plant-soil continuum along age classes in natural pinus tabuliformis carr. Forests of the eastern loess plateau, China. Forests 2023, 14, 44. [Google Scholar] [CrossRef]
- Liu, C.H.; Ma, J.Y.; Qu, T.T.; Xue, Z.J.; Li, X.Y.; Chen, Q.; Wang, N.; Zhou, Z.C.; An, S.S. Extracellular enzyme activity and stoichiometry reveal nutrient dynamics during microbially-mediated plant residue transformation. Forests 2023, 14, 34. [Google Scholar] [CrossRef]
- Wang, J.P.; Wu, Y.H.; Li, J.J.; He, Q.Q.; Bing, H.J. Soil enzyme stoichiometry is tightly linked to microbial community composition in successional ecosystems after glacier retreat. Soil Biol. Biochem. 2021, 162, 108429. [Google Scholar] [CrossRef]
- Redel, Y.; Rubio, R.; Godoy, R.; Borie, F. Phosphorus fractions and phosphatase activity in an andisol under different forest ecosystems. Geoderma 2006, 145, 216–221. [Google Scholar] [CrossRef]
- Deforest, J. Effects of elevated pH and phosphorus fertilizer on soil C, N and P enzyme stoichiometry in an acidic mixed mesophytic deciduous forest. Soil Biol. Biochem. 2020, 150, 107996. [Google Scholar] [CrossRef]
- Moorhead, D.L.; Sinsabaugh, R.L.; Hill, B.H.; Weintraub, M.N. Vector analysis of ecoenzyme activities reveal constraints on coupled C, N and P dynamics. Soil Biol. Biochem. 2016, 93, 1–7. [Google Scholar] [CrossRef]
- Srivastava, S.C.; Singh, J.S. Microbial c, n and p in dry tropical forest soils: Effects of alternate land-uses and nutrient flux. Soil Biol. Biochem. 1991, 23, 117–124. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Lauber, C.L.; Weintraub, M.N.; Ahmed, B.; Allison, S.D.; Crenshaw, C.; Contosta, A.R.; Cusack, D.; Frey, S.; Gallo, M.E.; et al. Stoichiometry of soil enzyme activity at global scale. Ecol. Lett. 2008, 11, 1252–1264. [Google Scholar] [CrossRef] [PubMed]
- Bi, B.Y.; Wang, Y.; Wang, K.; Zhang, H.; Fei, H.Y.; Pan, R.P.; Han, F.P. Changes in microbial metabolic C- and N-limitations in the rhizosphere and bulk soils along afforestation chronosequence in desertified ecosystems. J. Environ. Manag. 2022, 303, 114215. [Google Scholar] [CrossRef]
- Peng, X.Q.; Wang, W. Stoichiometry of soil extracellular enzyme activity along a climatic transect in temperate grasslands of northern China. Soil Biol. Biochem. 2016, 98, 74–84. [Google Scholar] [CrossRef]
- Li, Y.; Niu, S.L.; Yu, G.R. Aggravated phosphorus limitation on biomass production under increasing nitrogen loading: A meta-analysis. Glob. Chang. Biol. 2016, 22, 934–943. [Google Scholar] [CrossRef] [PubMed]
- Vitousek, P.M.; Porder, S.; Houlton, B.Z.; Chadwick, O.A. Terrestrial phosphorus limitation: Mechanisms, implications, and nitrogen–phosphorus interactions. Ecol. Appl. 2010, 20, 5–15. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.H.; Wang, C.K.; Jin, Y. Stoichiometric responses of soil microflora to nutrient additions for two temperate forest soils. Biol. Fertil. Soils 2017, 53, 397–406. [Google Scholar] [CrossRef]
- Clausing, S.; Pena, R.; Song, B.; Muller, K.; Mayer-Gruner, P.; Marhan, S.; Grafe, M.; Schulz, S.; Krueger, J.; Lang, F.; et al. Carbohydrate depletion in roots impedes phosphorus nutrition in young forest trees. New Phytol. 2021, 229, 2611–2624. [Google Scholar] [CrossRef]
- Bao, S.D. Soil Agrochemical Analysis; China Agricultural Press: Beijing, China, 2000. [Google Scholar]
- Kammann, C.I.; Schmidt, H.; Messerschmidt, N.; Linsel, S.; Steffens, D.; Müller, C.; Koyro, H.; Conte, P.; Joseph, S.; Stephen, J. Plant growth improvement mediated by nitrate capture in co-composted biochar. Sci. Rep. 2015, 5, 11080. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.G.; Dong, X.X.; Yao, W.X.; Han, C.; Sun, S.; Zhao, C.M. C, N, P, K stoichiometric characteristics of the “leaf-root-litter-soil” system in dryland plantations. Ecol. Indic. 2022, 143, 109371. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Yu, W.; Brookes, P.C.; Qiang, M.; Zhou, H.; Xu, Y.; Shen, S. Extraction of soil nitrogen by chloroform fumigation—A new index for the evaluation of soil nitrogen supply. Soil Biol. Biochem. 2011, 43, 2423–2426. [Google Scholar] [CrossRef]
- Fanin, N.; Fromin, N.; Buatois, B.; Httenschwiler, S. An experimental test of the hypothesis of non-homeostatic consumer stoichiometry in a plant litter-microbe system (letter). Ecol. Lett. 2013, 16, 764–772. [Google Scholar] [CrossRef] [PubMed]
- Brookes, P.C.; Landman, A.; Pruden, G.; Jenkinson, D.S. Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil—Sciencedirect. Soil Biol. Biochem. 1985, 17, 837–842. [Google Scholar] [CrossRef]
- Gao, M.L.; Song, W.H.; Zhou, Q.; Ma, X.J.; Chen, X.Y. Interactive effect of oxytetracycline and lead on soil enzymatic activity and microbial biomass. Environ. Toxicol. Pharmacol. 2013, 36, 667–674. [Google Scholar] [CrossRef] [PubMed]
- Guan, S.Y. Soil Enzyme and Its Research Methods; China Agriculture Press: Beijing, China, 1986; pp. 321–376. [Google Scholar]
- Sinsabaugh, R.L.; Hill, B.H.; Shah, J. Ecoenzymatic stoichiometry of microbial organic nutrient acquisition in soil and sediment (vol 462, pg 795, 2009). Nature 2010, 468, 122. [Google Scholar] [CrossRef] [Green Version]
- Hill, B.H.; Elonen, C.M.; Jicha, T.M.; Kolka, R.K.; Lehto, L.R.L.P.; Sebestyen, S.D.; Seifert-Monson, L.R. Ecoenzymatic stoichiometry and microbial processing of organic matter in northern bogs and fens reveals a common p-limitation between peatland types. Biogeochemistry 2014, 120, 203–224. [Google Scholar] [CrossRef]
- Moorhead, D.L.; Rinkes, Z.L.; Sinsabaugh, R.L.; Weintraub, M.N. Dynamic relationships between microbial biomass, respiration, inorganic nutrients and enzyme activities: Informing enzyme-based decomposition models. Front. Microbiol. 2013, 4, 223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.R.; Peng, D.L.; Tan, Z.J.; Guo, J.P.; Zhang, Y.X.; Liu, H.L. Do stand density and month regulate soil enzymes and the stoichiometry of differently aged larix principis-rupprechtii plantations? Catena 2023, 220, 106683. [Google Scholar] [CrossRef]
- Cleveland, C.C.; Liptzin, D. C:n:p stoichiometry in soil: Is there a “redfield ratio” for the microbial biomass? Biogeochemistry 2007, 85, 235–252. [Google Scholar] [CrossRef]
- Tian, H.Q.; Chen, G.S.; Zhang, C.; Melillo, J.M.; Hall, C.A.S. Pattern and variation of c:n:p ratios in China’s soils: A synthesis of observational data. Biogeochemistry 2010, 98, 139–151. [Google Scholar] [CrossRef]
- Li, W.N.; Huang, Z.Y.; Zhao, C.M.; Yang, M. Characteristics of soil microbial biomass C, N and nutrients in young plantations of Parashorea chinensis. J. Beijing For. Univ. 2020, 42, 51–62. [Google Scholar]
- Waring, B.G.; Weintraub, S.R.; Sinsabaugh, R.L. Ecoenzymatic stoichiometry of microbial nutrient acquisition in tropical soils. Biogeochemistry 2014, 117, 101–113. [Google Scholar] [CrossRef]
- Xu, Z.; Yu, G.; Zhang, X.; He, N.; Wang, Q.; Wang, S.; Wang, R.; Zhao, N.; Jia, Y.; Wang, C. Soil enzyme activity and stoichiometry in forest ecosystems along the north-south transect in eastern China (nstec). Soil Biol. Biochem. 2017, 104, 152–163. [Google Scholar] [CrossRef]
- Wang, M.W.; Ji, L.; Shen, F.Y.; Meng, J.; Wang, J.L.; Shan, C.F.; Yang, L.X. Differential responses of soil extracellular enzyme activity and stoichiometric ratios under different slope aspects and slope positions in larix olgensis plantations. Forests 2022, 13, 845. [Google Scholar] [CrossRef]
- Cui, Y.X.; Bing, H.J.; Fang, L.C.; Jiang, M.; Shen, G.T.; Yu, J.L.; Wang, X.; Zhu, H.; Wu, Y.H.; Zhang, X.C. Extracellular enzyme stoichiometry reveals the carbon and phosphorus limitations of microbial metabolisms in the rhizosphere and bulk soils in alpine ecosystems. Plant Soil 2021, 458, 7–20. [Google Scholar] [CrossRef]
- Cleveland, C.C.; Schmidt, T. Phosphorus limitation of microbial processes in moist tropical forests: Evidence from short-term laboratory incubations and field studies. Ecosystems 2002, 5, 680–691. [Google Scholar] [CrossRef]
- Deng, J.; Chong, Y.J.; Zhang, D.; Ren, C.J.; Zhao, F.Z.; Zhang, X.X.; Han, X.H.; Yang, G.H. Temporal variations in soil enzyme activities and responses to land-use change in the loess plateau, China. Appl. Sci. 2019, 9, 3129. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Haack, S.E.; Lin, W.; Li, B.; Wu, L.; Fang, C.; Zhang, Z. Soil microbial community structure and metabolic activity of pinus elliottii plantations across different stand ages in a subtropical area. PLoS ONE 2015, 10, e135354. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Du, H.; Zeng, F.; Song, T.; Peng, W. Diminished rhizosphere and bulk soil microbial abundance and diversity across succession stages in karst area, southwest China. Appl. Soil Ecol. 2020, 158, 103799. [Google Scholar] [CrossRef]
- Zeng, Y.L.; Fang, X.; Xiang, W.H.; Deng, X.W.; Peng, C.H. Stoichiometric and nutrient resorption characteristics of dominant tree species in subtropical chinese forests. Ecol. Evol. 2017, 7, 11033–11043. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Xue, L.; Jiao, R. Stoichiometric imbalances and the dynamics of phosphatase activity and the abundance of phoc and phod genes with the development of cunninghamia lanceolata (lamb.) Hook plantations. Appl. Soil Ecol. 2022, 173, 104373. [Google Scholar] [CrossRef]
- Deng, L.; Guan, Z.P.S. Afforestation drives soil carbon and nitrogen changes in China. Land Degrad. Dev. 2016, 28, 151–165. [Google Scholar] [CrossRef] [Green Version]
- Ali, A.; Hussain, M.; Ali, S.; Akhtar, K.; Muhammad, M.W.; Zamir, A.; Ali, A.; Nizami, S.M.; Ahmad, B.; Harrison, M.T.; et al. Ecological stoichiometry in pinus massoniana l. Plantation: Increasing nutrient limitation in a 48-year chronosequence. Forests 2022, 13, 469. [Google Scholar] [CrossRef]
- Dong, H.Y.; Ge, J.F.; Sun, K.; Wang, B.Z.; Xue, J.M. Change in root-associated fungal communities affects soil enzymatic activities during pinus massoniana forest development in subtropical China. For. Ecol. Manag. 2020, 482, 118817. [Google Scholar] [CrossRef]
- Wang, C.Q.; Jiao, R.Z. Adaptive pathways of microorganisms to cope with the shift from p- to n-limitation in subtropical plantations. Front. Microbiol. 2022, 13, 870667. [Google Scholar] [CrossRef]
- Han, W.Y.; Kemmitt, S.J.; Brookes, P.C. Soil microbial biomass and activity in chinese tea gardens of varying stand age and productivity. Soil Biol. Biochem. 2007, 39, 1468–1478. [Google Scholar] [CrossRef]
- Sanders, J.R. The effect of ph and organic matter of some soils on the ionic activities and concentrations of trace elements in soil solutions. J. Sci. Food Agric. 1983, 34, 52–55. [Google Scholar]
- Nannipieri, P.; Giagnoni, L.; Renella, G.; Puglisi, E.; Ceccanti, B.; Masciandaro, G.; Fornasier, F.; Moscatelli, M.C.; Marinari, S. Soil enzymology: Classical and molecular approaches. Biol. Fertil. Soils Coop. J. Int. Soc. Soil Sci. 2012, 48, 743–762. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, H.Y.; Lv, A.H. The variation of ecological stoichiometry characteristics of carbon, nitrogen and phosphorus in root system of myrica rubra and its soil microbial biomass with different stand ages. Ecol. Sci. 2022, 41, 84–90. [Google Scholar]
- Pan, J.W.; Guo, Q.Q.; Li, H.E.; Luo, S.Q.; Zhang, Y.Q.; Yao, S.; Fan, X.; Sun, X.G.; Qi, Y.J. Dynamics of soil nutrients, microbial community structure, enzymatic activity, and their relationships along a chronosequence of pinus massoniana plantations. Forests 2021, 12, 376. [Google Scholar] [CrossRef]
- Mooshammer, M.; Wanek, W.; Zechmeister-Boltenstern, S.; Richter, A. Stoichiometric imbalances between terrestrial decomposer communities and their resources: Mechanisms and implications of microbial adaptations to their resources. Front. Microbiol. 2014, 5, 22. [Google Scholar] [CrossRef] [Green Version]
- Jiao, P.Y.; Guo, W.; Chen, Z.L.; Liu, X.; Hu, Y.L.; Wang, Y.Z. Soil enzyme stoichiometric characteristics of pinus massoniana plantations at different stand ages in mid-subtropical areas. Environ. Sci. 2022, 43, 1059–1068. [Google Scholar]
- Osono, T.; Takeda, H. Potassium, calcium, and magnesium dynamics during litter decomposition in a cool temperate forest. J. For. Res. 2004, 9, 23–31. [Google Scholar] [CrossRef]
- Luo, H.Q.; Yu, J.L.; Li, R.X.; Gu, J.D.; Luo, L.; Zhang, Y.Y.; He, Y.; Xiao, Y.L.; Deng, S.H.; Zhang, Y.Z.; et al. Microbial biomass C:N:P as a better indicator than soil and ecoenzymatic c:n:p for microbial nutrient limitation and c dynamics in zoige plateau peatland soils. Int. Biodeterior. Biodegrad. 2022, 175, 105492. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stand Age | Soil Type | PH | Soil Organic Matter, SOM (g·kg−1) | Total Nitrogen, TN (g·kg−1) | Total Phosphorus, TP (g·kg−1) | Total Potassium, TK (g·kg−1) | Nitrate Nitrogen, NN (mg·kg−1) | Ammonium Nitrogen, AN (mg·kg−1) | Available Phosphorus, AP (mg·kg−1) | Available Potassium, AK (mg·kg−1) |
|---|---|---|---|---|---|---|---|---|---|---|
| 1a | rhizosphere | 4.58 ± 0.36 Ba | 11.98 ± 0.09 Fb | 1.84 ± 0.63 Ca | 0.32 ± 0.00 Da | 3.59 ± 0.14 Ca | 5.93 ± 0.06 Da | 6.61 ± 0.63 Ba | 3.44 ± 0.09 Da | 52.49 ± 4.76 Ea |
| non rhizosphere | 3.74 ± 0.03 Ab | 21.24 ± 1.07 Da | 1.37 ± 0.16 Ba | 0.26 ± 0.04 Ea | 3.98 ± 0.76 Db | 4.58 ± 0.30 Db | 6.26 ± 0.95 Ba | 2.23 ± 0.10 Bb | 55.24 ± 0.79 Ca | |
| 2a | rhizosphere | 3.40 ± 0.10 Aa | 16.65 ± 0.49 Ea | 1.72 ± 0.36 Ca | 0.63 ± 0.05 Ba | 2.91 ± 0.78 Ca | 11.23 ± 0.62 Ca | 11.75 ± 1.14 Aa | 4.67 ± 0.46 Ca | 51.20 ± 2.64 Ea |
| non rhizosphere | 3.29 ± 0.03 Ca | 17.23 ± 0.43 Ea | 1.15 ± 0.35 Ba | 0.43 ± 0.03 Cb | 3.67 ± 0.36 Da | 8.92 ± 0.66 Cb | 8.42 ± 0.27 Ab | 1.17 ± 0.09 Db | 53.12 ± 0.95 Ca | |
| 3a | rhizosphere | 3.49 ± 0.09 Ba | 27.97 ± 0.44 Ca | 2.43 ± 0.43 Ba | 0.89 ± 0.08 Aa | 6.70 ± 0.53 Ba | 13.16 ± 0.20 Ba | 6.43 ± 1.19 Bb | 5.10 ± 0.02 BCa | 75.76 ± 3.77 Ca |
| non rhizosphere | 3.27 ± 0.18 Ca | 9.43 ± 0.28 Fb | 1.50 ± 0.33 Bb | 0.53 ± 0.03 Bb | 7.48 ± 1.12 Ca | 8.51 ± 1.24 Cb | 7.66 ± 0.51 Aa | 1.06 ± 0.26 Db | 37.68 ± 0.72 Eb | |
| 8a | rhizosphere | 3.61 ± 0.11 Ba | 68.52 ± 1.41 Aa | 3.40 ± 0.38 Aa | 0.32 ± 0.03 Da | 6.58 ± 1.34 Bb | 13.58 ± 1.12 Ba | 5.40 ± 0.56 BCa | 5.48 ± 0.18 Ba | 87.76 ± 2.72 Ba |
| non rhizosphere | 3.54 ± 0.01 Ba | 39.36 ± 2.16 Ab | 2.69 ± 0.15 Ab | 0.32 ± 0.02 Da | 9.31 ± 0.34 Ba | 11.28 ± 0.07 Bb | 4.37 ± 0.63 Ca | 4.50 ± 0.19 Ab | 46.68 ± 1.40 Db | |
| 9a | rhizosphere | 3.47 ± 0.01 Ba | 48.27 ± 1.85 Ba | 3.40 ± 0.27 Aa | 0.53 ± 0.03 Ca | 6.54 ± 1.03 Bb | 20.44 ± 1.76 Aa | 4.74 ± 0.09 Ca | 7.38 ± 0.38 Aa | 108.78 ± 0.77 Aa |
| non rhizosphere | 3.54 ± 0.01 Ba | 31.11 ± 0.50 Bb | 2.31 ± 0.09 ABb | 0.34 ± 0.01 Db | 12.58 ± 0.85 Aa | 15.76 ± 0.44 Ab | 3.30 ± 0.38 Db | 4.40 ± 0.37 Ab | 87.49 ± 5.72 Ab | |
| 42a | rhizosphere | 3.69 ± 0.16 Ba | 23.71 ± 0.23 Da | 1.70 ± 0.13 Ca | 0.96 ± 0.05 Aa | 13.2 ± 0.92 Aa | 10.43 ± 0.44 Ca | 3.11 ± 0.10 Da | 2.65 ± 0.15 Ea | 65.82 ± 1.37 Da |
| non rhizosphere | 3.72 ± 0.13 Aa | 23.08 ± 0.29 Ca | 2.27 ± 0.43 ABa | 0.78 ± 0.01 Ab | 6.92 ± 0.38 Cb | 9.11 ± 0.29 Cb | 3.26 ± 0.08 Da | 1.56 ± 0.06 Cb | 61.42 ± 1.02 Ba |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, W.; Ali, I.; Han, X.; Ullah, S.; Yang, M. Soil C, N, P, K and Enzymes Stoichiometry of an Endangered Tree Species, Parashorea chinensis of Different Stand Ages Unveiled Soil Nutrient Limitation Factors. Forests 2023, 14, 624. https://doi.org/10.3390/f14030624
Li W, Ali I, Han X, Ullah S, Yang M. Soil C, N, P, K and Enzymes Stoichiometry of an Endangered Tree Species, Parashorea chinensis of Different Stand Ages Unveiled Soil Nutrient Limitation Factors. Forests. 2023; 14(3):624. https://doi.org/10.3390/f14030624
Chicago/Turabian StyleLi, Wannian, Izhar Ali, Xiaomei Han, Saif Ullah, and Mei Yang. 2023. "Soil C, N, P, K and Enzymes Stoichiometry of an Endangered Tree Species, Parashorea chinensis of Different Stand Ages Unveiled Soil Nutrient Limitation Factors" Forests 14, no. 3: 624. https://doi.org/10.3390/f14030624
APA StyleLi, W., Ali, I., Han, X., Ullah, S., & Yang, M. (2023). Soil C, N, P, K and Enzymes Stoichiometry of an Endangered Tree Species, Parashorea chinensis of Different Stand Ages Unveiled Soil Nutrient Limitation Factors. Forests, 14(3), 624. https://doi.org/10.3390/f14030624