
Use of an Arboretum and DNA Barcoding for the Detection and Identification of Leaf-Mining Insects on Alien Woody Plants




Abstract
:1. Introduction
2. Materials and Methods
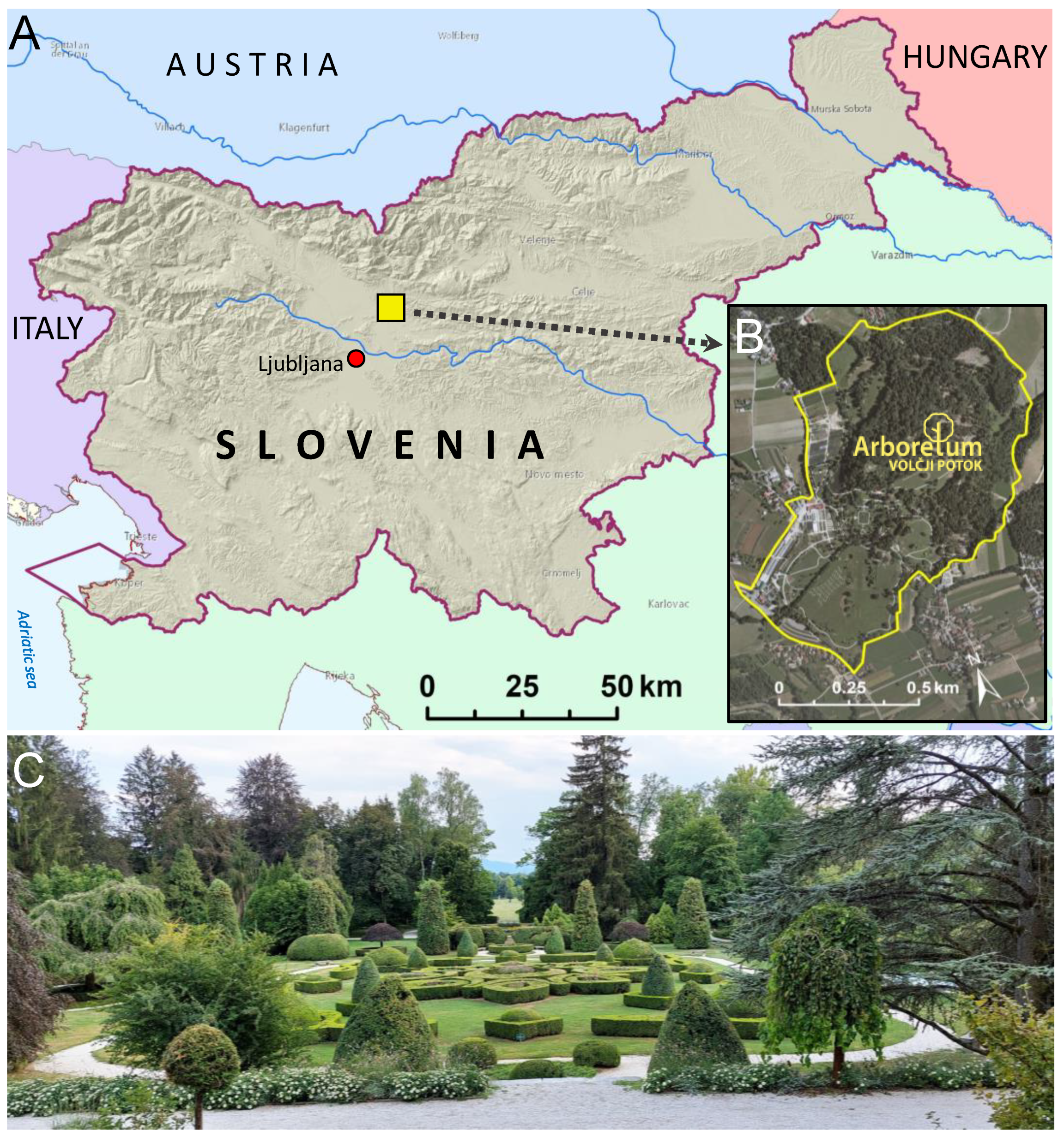
2.1. Surveyed Arboretum
2.2. Woody Plants
2.3. Leaf Miner Sampling
2.4. Leaf Miner Identification
2.5. DNA Barcoding
2.6. Analysis of Trophic Associations
3. Results
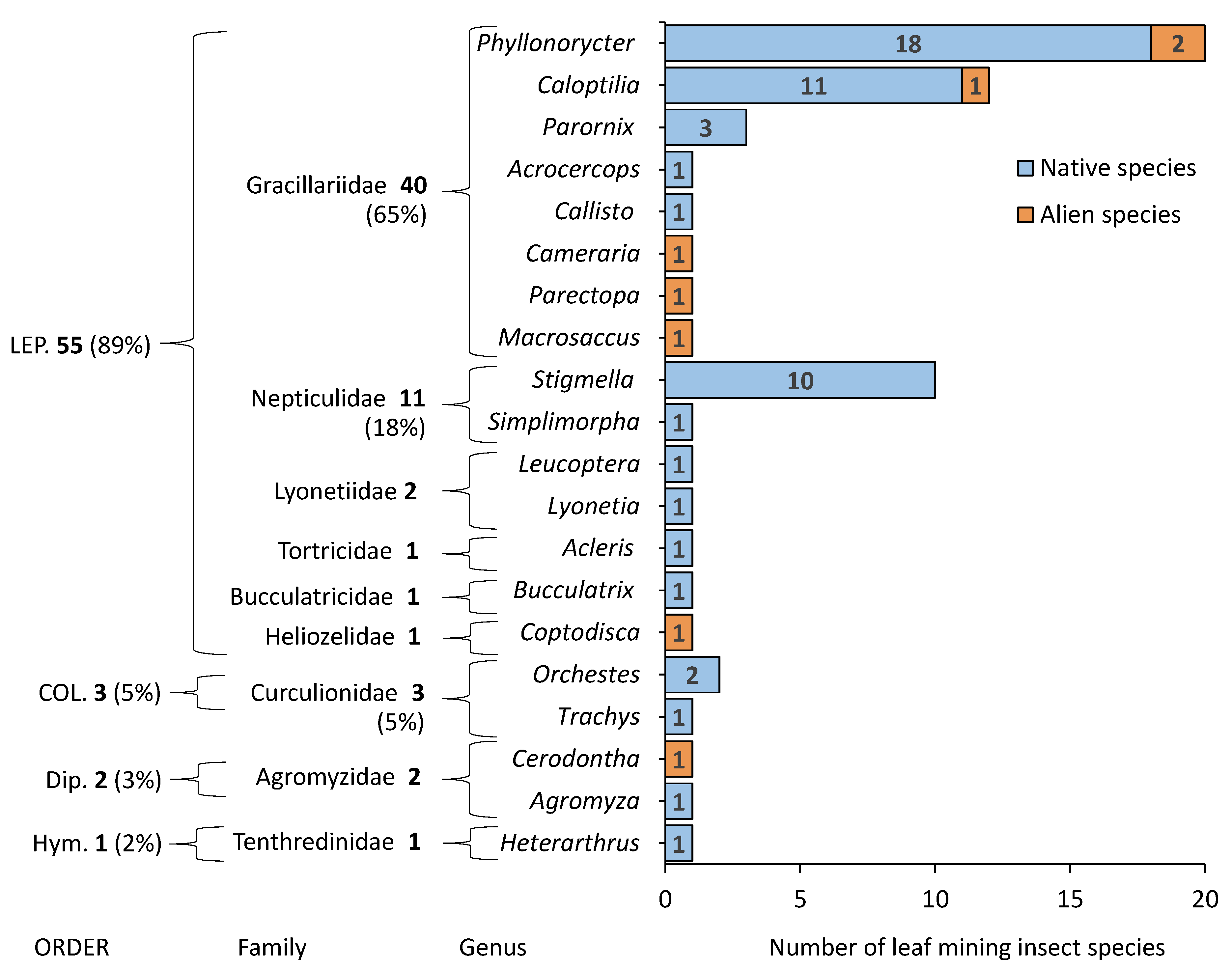
3.1. Leaf Miner Diversity on Woody Plants in the Arboretum
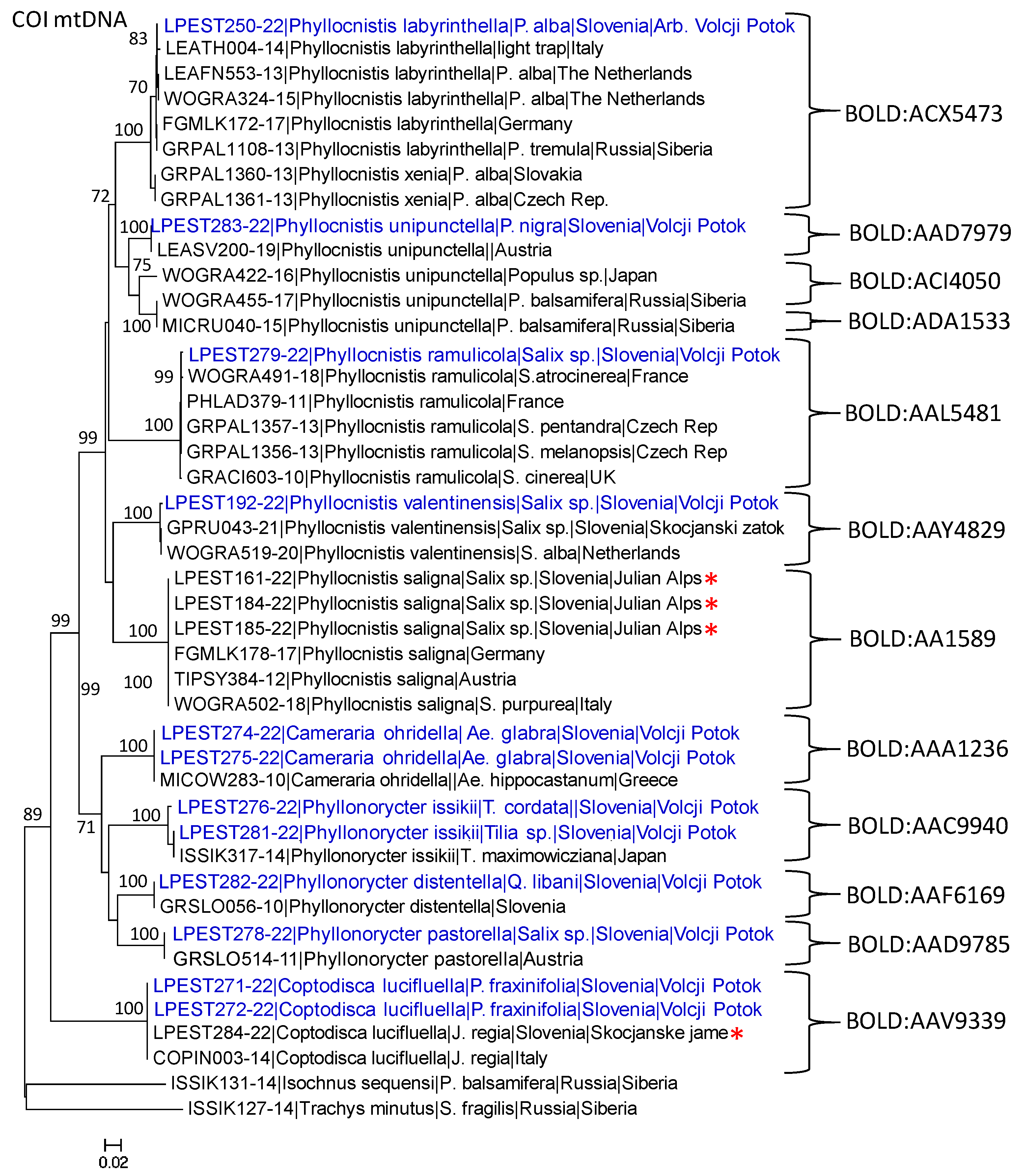
3.2. DNA Barcoding Data
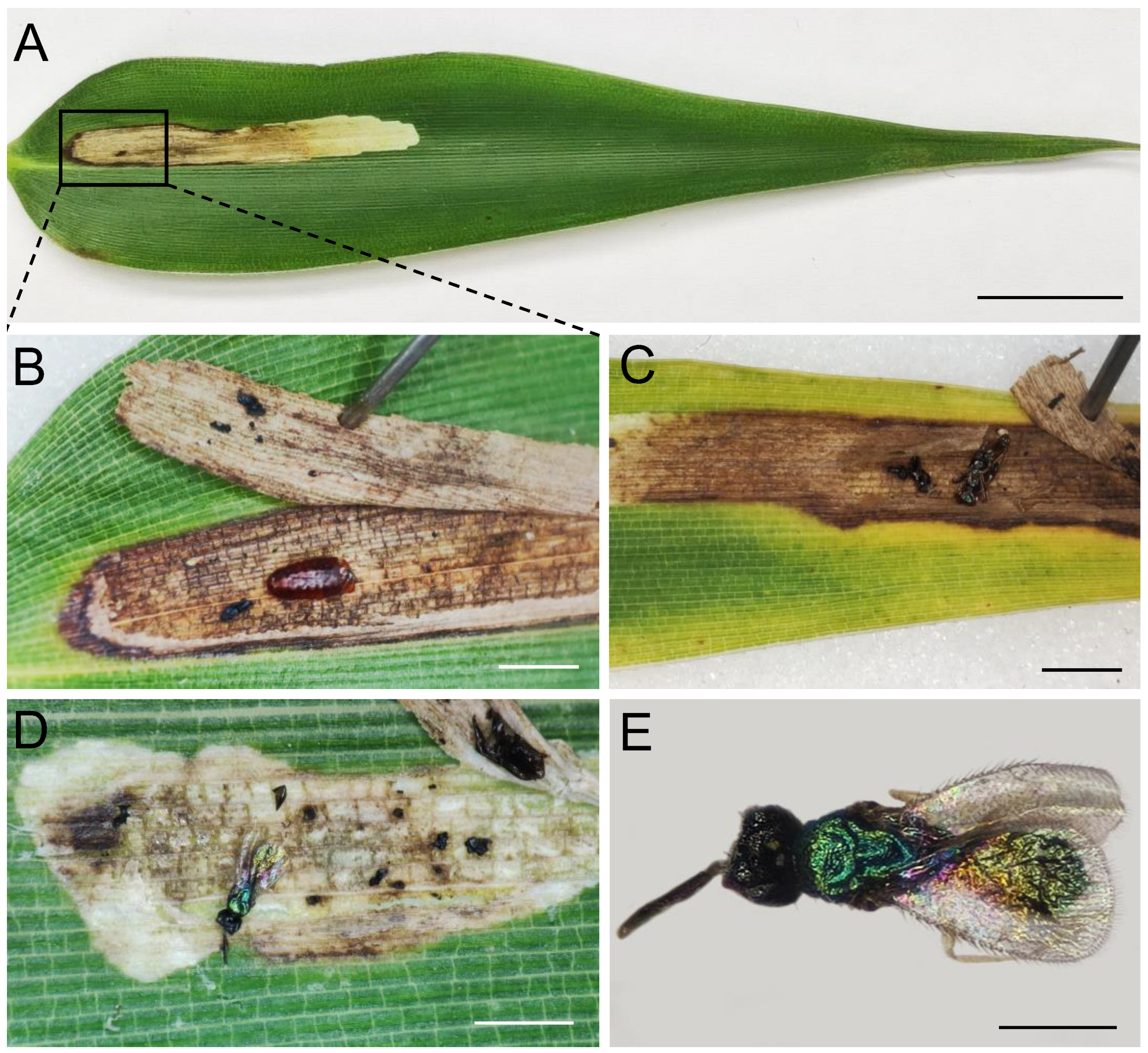
3.3. Leaf Miner Damage
3.4. Trophic Associations
3.5. Leaf-Mining Species New to Slovenia
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Diagne, C.; Leroy, B.; Vaissière, A.C.; Gozlan, R.E.; Roiz, D.; Jarić, I.; Salles, J.-M.; Corey, J.; Bradshaw, A.; Courchamp, F. High and rising economic costs of biological invasions worldwide. Nature 2021, 592, 571–576. [Google Scholar] [CrossRef] [PubMed]
- Meyer, S.E.; Callaham, M.A.; Stewart, J.E.; Warren, S.D. Invasive species response to natural and anthropogenic disturbance. In Invasive Species in Forests and Rangelands of the United States; Poland, T.M., Patel-Weynand, T., Finch, D.M., Miniat, C.F., Hayes, D.C., Lopez, V.M., Eds.; Springer: Cham, Switzerland, 2021; pp. 85–110. [Google Scholar] [CrossRef]
- MacLachlan, M.J.; Liebhold, A.M.; Yamanaka, T.; Springborn, M.R. Hidden patterns of insect establishment risk revealed from two centuries of alien species discoveries. Sci. Adv. 2021, 7, eabj1012. [Google Scholar] [CrossRef] [PubMed]
- de Groot, M.; O’Hanlon, R.; Bullas-Appleton, E.; Csóka, G.; Csiszár, Á.; Faccoli, M.; Gervasini, E.; Kirichenko, N.; Korda, M.; Marinšek, A.; et al. Challenges and solutions in early detection, rapid response and communication about potential invasive alien species in forests. Manag. Biol. Invasions 2020, 11, 637–660. [Google Scholar] [CrossRef]
- Bennett, B. Learning in paradise: The role of botanic gardens in university education. In Innovative Strategies for Teaching in the Plant Sciences; Quave, C., Ed.; Springer: New York, NY, USA, 2014; pp. 213–229. [Google Scholar] [CrossRef]
- Heywood, V.H. The future of plant conservation and the role of botanic gardens. Plant Divers. 2017, 39, 309–313. [Google Scholar] [CrossRef]
- Chen, G.; Sun, W. The role of botanical gardens in scientific research, conservation, and citizen science. Plant Divers. 2018, 40, 181–188. [Google Scholar] [CrossRef]
- Hulme, P.E. Addressing the threat to biodiversity from botanic gardens. Trends Ecol. Evol. 2011, 26, 168–174. [Google Scholar] [CrossRef]
- Ferus, P.; Hoťka, P.; Košútová, D.; Konôpková, J. Invasions of alien woody plant taxa across a cluster of villages neighbouring the Mlyňany Arboretum (SW Slovakia). Folia Oecol. 2020, 47, 121–130. [Google Scholar] [CrossRef]
- Wondafrash, M.; Wingfield, M.J.; Wilson, J.R.U.; Hurley, B.P.; Slippers, B.; Paap, T. Botanical gardens as key resources and hazards for biosecurity. Biodiv. Conserv. 2021, 30, 1929–1946. [Google Scholar] [CrossRef]
- Eschen, R.; O’Hanlon, R.; Santini, A.; Vannini, A.; Roques, A.; Kirichenko, N.; Kenis, M. Safeguarding global plant health: The rise of sentinels. J. Pest. Sci. 2019, 92, 29–36. [Google Scholar] [CrossRef] [Green Version]
- Morales-Rodríguez, C.; Bastianelli, G.; Aleandri, M.; Doğmuş-Lehtijärvi, T.; Oskay, F.; Vannini, A. Revealing novel interactions between oak and Tubakia species: Evidence of the efficacy of the sentinel arboreta strategy. Biol. Invasions 2021, 23, 3749–3765. [Google Scholar] [CrossRef]
- Roques, A.; Fan, J.; Courtial, B.; Zhang, Y.; Yart, A.; Auger-Rozenberg, M.-A.; Denux, O.; Kenis, M.; Baker, R.; Sun, J.-H. Planting sentinel European trees in Eastern Asia as a novel method to identify potential insect pest invaders. PLoS ONE 2017, 10, e0120864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Culley, T.M.; Dreisilker, K.; Clair Ryan, M.; Schuler, J.A.; Cavallin, N.; Gettig, R.; Havens, K.; Landel, H.; Shultz, B. The potential role of public gardens as sentinels of plant invasion. Biodivers. Conserv. 2022, 31, 1829–1844. [Google Scholar] [CrossRef]
- Tomoshevich, M.; Kirichenko, N.; Holmes, K.; Kenis, M. Foliar fungal pathogens of European woody plants in Siberia: An early warning of potential threats? For. Pathol. 2013, 43, 345–359. [Google Scholar] [CrossRef]
- Kirichenko, N.; Kenis, M. Using a botanical garden to assess factors influencing the colonization of exotic woody plants by phyllophagous insects. Oecologia 2016, 182, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Franić, I.; Prospero, S.; Adamson, K.C.; Allan, E.; Attorre, F.; Auger-Rozenberg, M.-A.; Augustin, S.; Avtzis, D.; Baert, W.; Barta, M.; et al. Worldwide diversity of endophytic fungi and insects associated with dormant tree twigs. Sci. Data 2022, 9, 1–9. [Google Scholar] [CrossRef]
- Barham, E. The unique role of sentinel trees, botanic gardens and arboreta in safeguarding global plant health. Plant Biosyst. Int. J. Deal. Asp. Plant Biol. 2016, 150, 377–380. [Google Scholar] [CrossRef]
- Kenis, M.; Li, H.; Fan, J.; Courtial, B.; Auger-Rozenberg, M.-A.; Yart, A.; Eschen, R.; Roques, A. Sentinel nurseries to assess the phytosanitary risks from insect pests on importations of live plants. Sci. Rep. 2018, 8, 11217. [Google Scholar] [CrossRef] [Green Version]
- Roques, A.; Cleary, M.; Matsiakh, I.; Eschen, R. (Eds.) Field Guide for the Identification of Damage on Woody Sentinel Plants; CAB International: Wallingford, UK, 2017; p. 263. [Google Scholar] [CrossRef] [Green Version]
- Morales-Rodríguez, C.; Anslan, S.; Auger-Rozenberg, M.-A.; Augustin, S.; Baranchikov, Y.; Bellahirech, A.; Burokienė, D.; Čepukoit, D.; Çota, E.; Davydenko, K.; et al. Forewarned is forearmed: Harmonized approaches for early detection of potentially invasive pests and pathogens in sentinel plantings. NeoBiota 2019, 47, 95–123. [Google Scholar] [CrossRef] [Green Version]
- Ellis, W. Plant Parasites of Europe: Leafminers, Galls and Fungi. Available online: http://bladmineerders.nl/ (accessed on 24 January 2023).
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; deWaard, J.R. Biological identifications through DNA barcodes. Proc. Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Vaamonde, C.; Kirichenko, N.; Cama, A.; Doorenweerd, C.; Godfray, H.C.J.; Guiguet, A.; Gomboc, S.; Huemer, P.; Landry, J.-F.; Laštůvka, A.; et al. Evaluating DNA barcoding for species identification and discovery in European gracillariid moths. Front. Ecol. Evol. 2021, 9, 626752. [Google Scholar] [CrossRef]
- Hering, E.M. Biology of the Leaf Miners; Junk, W., Ed.; Gravenhage: The Hague, The Netherlands, 1951; p. 490. [Google Scholar]
- Connor, E.F.; Taverner, M.P. The evolution and adaptive significance of the leaf-mining habit. Oikos 1997, 79, 6–25. [Google Scholar] [CrossRef] [Green Version]
- Kirichenko, N.; Augustin, S.; Kenis, M. Invasive leafminers on woody plants: A global review of pathways, impact and management. J. Pest Sci. 2019, 92, 93–106. [Google Scholar] [CrossRef] [Green Version]
- Mastnak, M. A Short Guide to the Volčji Potok Arboretum; Arboretum: Volčji Potok, Slovenia, 2005; p. 50. [Google Scholar]
- ESRI ArcGIS Pro Software. Available online: https://www.esri.com/en-us/arcgis/products/arcgis-pro/overview (accessed on 30 January 2022).
- ESRI ArcGIS Online Countries and River Layers. Available online: https://www.arcgis.com/home/index.html (accessed on 30 January 2022).
- Ministry of Public Administration of the Republic of Slovenia: GIS Portal. Map Services. Available online: https://gis.gov.si/arcgis/services (accessed on 30 January 2022).
- Ogrin, D.; Vysoudil, M.; Ogrin, M.; Koželj, T. Topoklimatske razmere. In Kamniška Bistrica—Geografska Podoba Gorske Doline; Ogrin, D., Ed.; GeograFF 22; Znanstvena Založba Filozofske Fakultete Univerze v Ljubljani: Ljubljana, Slovenia, 2018; pp. 45–70. [Google Scholar] [CrossRef]
- Mastnak, M.; Arboretum Volčji Potok, Gorenjska region, Slovenia. On insecticide treatment in the Arboretum Volčji Potok. Personal communication, 2023. [Google Scholar]
- Martinčič, A. (Ed.) Mala Flora Slovenije, 4th ed.; Tehniška Založba Slovenije: Ljubljana, Slovenia, 2007. (In Slovenian) [Google Scholar]
- GBIF.org, GBIF Home Page. 2023. Available online: https://www.gbif.org (accessed on 24 January 2023).
- Laštůvka, A.; Laštůvka, Z.; Liska, J.; Šumpich, J. Motyli a Housenky Stredni Evropy, V. Drobni Motyli I [Moths and Caterpillars of Central Europe V—Micromoths; Atlas, I., Ed.; Academia: Praha, Czech Republic, 2018; p. 532. (In Czech) [Google Scholar]
- Maček, J. Hiponomološka Favna Slovenije/Hyponomologische Fauna Sloweniens [Leaf Mining Fauna of Slovenia]. Slovenska akademija znanosti in umetnosti, Razred za naravoslovne vede, Classis IV. Hist. Nat. 1999, 37, 385. (In Slovenian) [Google Scholar]
- Gomboc, S.; Kirichenko, N.I. An Overview of Gracillariidae leaf mining moths in Slovenia with new records for the country. Diversity 2022, 14, 811. [Google Scholar] [CrossRef]
- de Waard, J.; Ivanova, N.; Hajibabaei, M.; Hebert, P.D.N. Assembling DNA barcodes: Analytical methods. In Methods in Molecular Biology: Environmental Genetic; Cristofre, M., Ed.; Humana Press: Totowa, NJ, USA, 2008; pp. 275–293. [Google Scholar]
- Ratnasingham, S.; Hebert, P.D.N. A DNA-Based Registry for All Animal Species: The Barcode Index Number (BIN) System. PLoS ONE 2013, 8, e66213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeanmougin, F.; Thompson, J.D.; Gouy, M.; Higgins, D.G.; Gibson, T.J. Multiple sequence alignment with Clustal, X. Trends Biochem. Sci. 1998, 23, 403–405. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Molec. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- De Prins, J.; De Prins, W. Global Taxonomic Database of Gracillariidae. Available online: http://www.gracillariidae.net/ (accessed on 27 January 2023).
- Dormann, C.F.; Fruend, J.; Gruber, B.; Beckett, S.; Devoto, M.; Felix, M.F.; Gabriel, M.F.; Iriondo, J.M.; Opsahl, T.; Pinheiro, R.B.P.; et al. Visualising Bipartite Networks and Calculating Some (Ecological) Indices. Version 2.18. Available online: chrome-extension://efaidnbmnnnibpcajpcglclefindmkaj/https://cran.r-project.org/web/packages/bipartite/bipartite.pdf (accessed on 3 February 2023).
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. Available online: https://www.R-project.org/ (accessed on 24 January 2023).
- Dormann, C.F.; Gruber, B.; Fruend, J. Introducing the bipartite Package: Analysing Ecological Networks. R News 2008, 8, 8–11. [Google Scholar]
- Zlobin, V.V. Review of mining flies of the genus Cerodontha. IV. Subgenus Poemyza (Diptera: Agromyzidae). Zoosyst. Rossica 1993, 1, 117–141. [Google Scholar]
- Süss, L. Cerodontha (Poemyza) unisetiorbita Zlobin (Diptera Agromyzidae) nuova per l´Europa. Boll. Zool. Agrar. Bachic. Ser. II 2001, 33, 73–77. [Google Scholar]
- Hannover, R. Available online: http://www.diptera.info/forum/viewthread.php?forum_id=4&thread_id=56490 (accessed on 27 January 2023).
- Černý, M.; Roháček, J. Cerodontha (Poemyza) unisetiorbita Zlobin, 1993 (Diptera: Agromyzidae), a leaf-miner on bamboo: First records from Central Europe. Acta Musei Sil. Sci. Nat. 2015, 64, 91–96. [Google Scholar]
- British Leafminers. Cerodontha Unisetiorbita Zlobin. 1993. Available online: http://www.leafmines.co.uk/html/Diptera/C.unisetiorbita.htm (accessed on 27 January 2023).
- Heckford, R.J. Cerodontha (Poemyza) unisetiorbita Zlobin (Diptera, Agromyzidae), an adventive species new to the British Isles. Dipterists. Digest. 2017, 24, 49–51. [Google Scholar]
- Kirichenko, N.; Sukachev Institute of Forest, Siberian Branch of the Russian Academy of Sciences, Federal Research Center “Krasnoyarsk Science Center SB RAS”, Krasnoyarsk, Russia. Detection of leaf mines of Cerodontha unisetiorbita on bamboo in the Parc Floral de La Source, Orleans. Unpublished work. 2014. [Google Scholar]
- Bernardo, U.; Sasso, R.; Gebiola, M.; Viggiani, G. First record of a walnut shield bearer Coptodisca (Lepidoptera: Heliozelidae) in Europe. J. Appl. Entomol. 2011, 136, 638–640. [Google Scholar] [CrossRef]
- Takács, A.; Szabóky, C.; Tóth, B.; Bozsó, M.; Kutas, J.; Molnár, S.; Richter, I. Nearctic walnut leafminers invade Europe: First Coptodisca lucifluella (Clemens, 1860) and now Coptodisca juglandiella (Chambers, 1874) (Lepidoptera, Heliozelidae). Nota Lepidopterol. 2020, 43, 77–93. [Google Scholar] [CrossRef]
- North American Moth Photographers Group. Coptodisca Lucifluella (Clemens, 1860); Mississippi State University: Starkville, MI, USA, 2013; Available online: http://mothphotographersgroup.msstate.edu/large_map.php?hodges=247 (accessed on 27 January 2023).
- Bernardo, U.; van Nieukerken, E.J.; Sasso, R.; Gebiola, M.; Gualtieri, L.; Viggiani, G. Characterization, distribution, biology and impact on Italian walnut orchards of the invasive North-American leafminer Coptodisca lucifluella (Lepidoptera: Heliozelidae). Bull. Entomol. Res. 2015, 105, 210–224. [Google Scholar] [CrossRef]
- Takács, A.; Szabóky, C.; Kutas, J. A dióaknázó fényesmoly (Coptodisca lucifluella Clemens, 1860 Lepidoptera—Heliozelidae) magyarországi megjelenése. [The appearance of the walnut leafminer (Coptodisca lucifluella Clemens, 1860 Lepidoptera—Heliozelidae) in Hungary]. Növényvédelem 2017, 78, 539–541. (In Hungarian) [Google Scholar]
- Pályi, B.; Takács, A.; Szabóky, C.S. Új diókártevő Kárpátalján. [New pest of walnut in Subcarpathia]. Kárpátaljai Vállalkozók Lapja 2019, 4, 16. (In Hungarian) [Google Scholar]
- Tomov, R. First Records of the Walnut Shield Bearer Coptodisca lucifluella (Clemens, 1860) (Lepidoptera: Heliozelidae) in Bulgaria. Acta Zool. Bulg. 2020, 72, 697–700. [Google Scholar]
- Chireceanu, C.; Mustățea, R.-V.; Teodoru, A. The walnut shield bearer Coptodisca lucifluella (Clemens, 1860) (Lepidoptera: Heliozelidae)—The first record in Romania. Rom. J. Plant Prot. 2022, XV, 15–23. [Google Scholar] [CrossRef]
- Gregor, F.; Patočka, J. Die Puppen der mitteleuropischen Lithocolletinae. Mitteilungen des internationalen entomologischen Vereins. Supplement 2001, 8, 186. (In German) [Google Scholar]
- Huemer, P. Die Schmetterlinge Österreichs (Lepidoptera). Systematische und Faunistische Checkliste; Studiohefte 12; Tiroler Landesmuseen: Innsbruck, Austria, 2013; p. 304. [Google Scholar]
- Baryshnikova, S.V. Gracillariidae. In Catalogue of the Lepidoptera of Russia, 2nd ed.; Sinev, S.Y., Ed.; Zoological Institute RAS: St. Petersburg, Russia, 2019; pp. 36–43. (In Russian) [Google Scholar]
- Kuznetzov, V.I. The family Gracillariidae (Lithocolletidae). In Insects and Mites—The Pests of Agricultural Plants. III (2). Lepidoptera; Kuznetsov, V.I., Ed.; Nauka: St. Petersburg, Russia, 1999; pp. 9–46. (In Russian) [Google Scholar]
- Leraut, P. Contribution à l’étude des Lépidoptéres de la Brie occidentale. Alexanor 2003, 22, 3–237. [Google Scholar]
- Huisman, K.J.; Koster, J.C.; van Nieukerken, E.J.; Ulenberg, S.A. Microlepidoptera in Nederland in 2003. Entomol. Ber. 2005, 65, 30–42. [Google Scholar]
- Aarvik, L.; Bengtsson, B.Å.; Elven, H.; Ivinskis, P.; Jürivete, U.; Karsholt, O.; Mutanen, M.; Savenkov, N. Nordic-Baltic Checklist of Lepidoptera. Nor. J. Entomol. 2017, 3, 1–236. [Google Scholar]
- Emmet, A.M. Phyllocnistidae. In The Moths and Butterflies of Great Britain and Ireland; Heath, J., Emmet, A.M., Eds.; Harley Books: Devon, UK, 1985; Volume 2, pp. 363–368. [Google Scholar]
- Huemer, P.; Tarmann, G. Die Schmetterlinge Österreichs. Systematisches Verzeichnis mit Verbreitungsangaben für die einzelnen Bundesländer. Veröff. tirol. Landesmus Ferdinandeum Innsbr. 1993, 5, 1–224. (In German) [Google Scholar]
- Langmaid, J.; Corley, M. Phyllocnistis ramulicola sp.nov (Lepidoptera: Gracillariidae) in England and Portugal. Entomol. Gazette 2007, 58, 227–237. [Google Scholar]
- Tokár, Z.; Laštůvka, A.; Pastorális, G.; Šumpich, J.; Štefanovič, R.; Elsner, G. Nové druhy drobných motýľov (Microlepidoptera) pre faunu Slovenska. Entomofauna Carpathica 2021, 33, 1–20. (In Slovenian) [Google Scholar]
- Mutanen, M.; Kivelä, S.M.; Vos, R.A.; Doorenweerd, C.; Ratnasingham, S.; Hausmann, A.; Huemer, P.; Dincă, V.; van Nieukerken, E.J.; Lopez-Vaamonde, C.; et al. Species-level para- and polyphyly in dna barcode gene trees: Strong operational bias in European Lepidoptera. Syst. Biol. 2016, 65, 1024–1040. [Google Scholar] [CrossRef] [PubMed]
- Thomas, E. Phyllocnistis ramulicola Langmaid & Corley. 2007. Available online: https://oreina.org/artemisiae/observatoire/index.php?module=fiche&action=fiche&d=micro&id=814417 (accessed on 27 December 2022).
- Beránek, J. First records of Leptoglossus occidentalis Heidemann, 1910 (Heteroptera: Pentatomorpha: Coreidae) in the Czech Republic. Plant Protect. Sci. 2007, 43, 165–168. [Google Scholar] [CrossRef] [Green Version]
- Matošević, D. Box Tree Moth (Cydalima perspectalis, Lepidoptera; Crambidae), new invasive insect pest in Croatia. South-East Eur. For. 2013, 4, 89–94. [Google Scholar] [CrossRef] [Green Version]
- Baranchikov, Y.N.; Seraya, L.G.; Grinash, M.N. All European ash species are susceptible to Emerald ash borer Agrilus planipennis Fairmaire (Coleoptera: Buprestidae)—A Far Eastern Invader. Sib. For. J. 2014, 6, 80–85. (In Russian) [Google Scholar]
- Kerchev, I.A.; Krivets, S.A. An attack of Ips amitinus (Coleoptera: Curculionidae: Scolytinae) on arboretum in West Siberia: New host of invasive bark beetle among exotic conifers. J. Asia-Pac. Entomol. 2021, 24, 148–152. [Google Scholar] [CrossRef]
- Rudoi, V.V.; Vinokurov, N.N.; Korshunov, A.V.; Efimov, D.A.; Kirichenko, N.I. New records of native and alien true bugs (Heteroptera) from Kemerovo Region, Western Siberia, Russia. Acta Biol. Sibirica 2022, 8, 483–506. [Google Scholar] [CrossRef]
- Gomboc, S. Novi vrsti listnih zavrtačev v Sloveniji [New records of leaf miners in Slovenia]. In Proceedings of the Zbornik Predavanj in Referatov 6, Slovenskega Posvetovanja o Varstvu Rastlin, Zreče, Slovenia, 4 June 2003; Društvo za Varstvo Rastlin Slovenije: Zreče, Slovenia, 2003; pp. 423–429. (In Slovenian). [Google Scholar]
- Seljak, G. The dynamics of introduction of alien phytophagous insects and mites into Slovenia. Acta Entomol. Slov. 2013, 21, 85–122. (In Slovenian) [Google Scholar]
- Seljak, G. Six new alien phytophagous insect species recorded in Slovenia in 2011. Acta Entomol. Slov. 2012, 20, 31–44. (In Slovenian) [Google Scholar]
- Mastnak, M.; Arboretum Volčji Potok, Gorenjska region, Slovenia. About absence of some woody plants from live collection of the Arboretum Volčji Potok. Personal communication, 2023. [Google Scholar]
- Kranzfelder, P.; Ekrem, T.; Stur, E. Trace DNA from insect skins: A comparison of five extraction protocols and direct PCR on chironomid pupal exuviae. Mol. Ecol. Resour. 2016, 16, 353–363. [Google Scholar] [CrossRef]
- Mlynarek, J.J.; Kim, J.-H.; Heard, S.B. Identification of leaf-mining insects via DNA recovered from empty mines. FACETS 2016, 1, 217–224. [Google Scholar] [CrossRef]
- Jordan, M.P.; Langmaid, J.R.; Doorenweerd, C. Morphological difference between upperside and underside leaf-mining larvae of Phyllocnistis unipunctella (Stephens, 1834) (Lep.: Gracillariidae) and its changing phenology. Entomol. Rec. J. Var. 2016, 128, 121–127. [Google Scholar]
- PM 3/91(1) Sentinel woody plants: Concepts and application. Bull. OEPP/EPPO Bull. 2020, 50, 429–436. [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
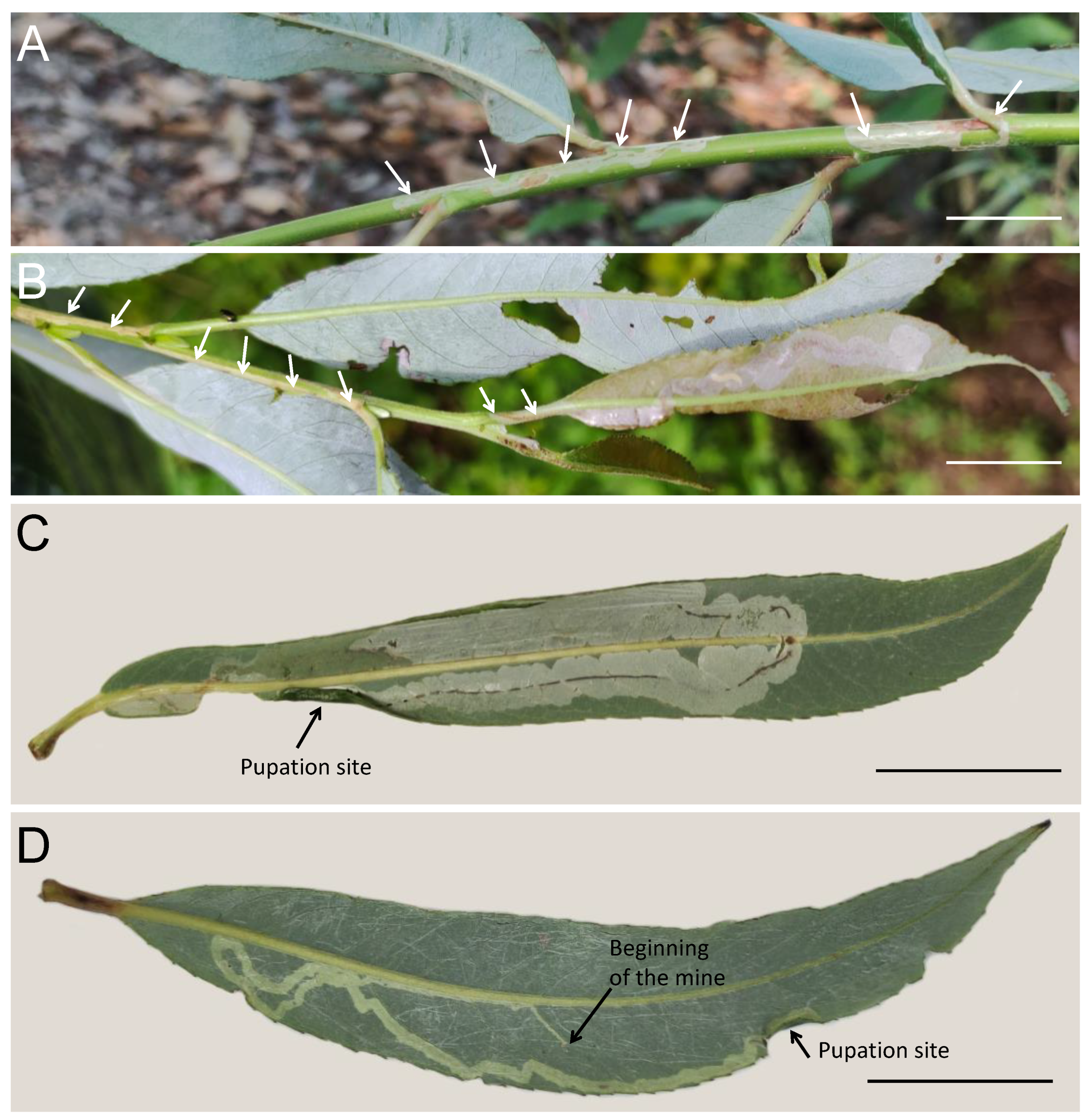{kind=link}
| No. | Species 1 | Species | |||||
|---|---|---|---|---|---|---|---|
| P. labyrinthella | P. xenia * | P. unipunctella | P. valentinensis | P. saligna | P. ramulicola | ||
| 1 | Phyllocnistis labyrinthella | [0.46] | |||||
| 2 | Phyllocnistis xenia * | 1.08 | [0] | ||||
| 3 | Phyllocnistis unipunctella | 8.21 | 7.85 | [5.81] | |||
| 4 | Phyllocnistis valentinensis | 11.19 | 11.19 | 11.47 | [0.31] | ||
| 5 | Phyllocnistis saligna | 12.09 | 12.27 | 12.09 | 10.84 | [0] | |
| 6 | Phyllocnistis ramulicola | 12.63 | 12.82 | 12.42 | 14.87 | 13.57 | [0.31] |
| No. | Plant Species | Plant Origin 1 | Insect Family | Insect Species 2 | Damage Level 3 | Sampling Date |
|---|---|---|---|---|---|---|
| 1 | Acer saccharinum | NA | Gracillariidae | Caloptilia sp. 3 |  | 9 August 2022 |
| 2 | Aesculus glabra var. arguta | NA | Nepticulidae | Stigmella sp. 1 | | 6 August 2022 |
| 3 | Betula utilis | EA | Nepticulidae | Stigmella lapponica | | 9 August 2022 |
| 4 | Betula utilis | EA | Gracillariidae | Parornix betulae | | 9 August 2022 |
| 5 | Celtis julianae | EA | Gracillariidae | Phyllonorycter millierella | | 9 August 2022 |
| 6 | Celtis julianae | EA | Gracillariidae | Caloptilia fidella | | 9 August 2022 |
| 7 | Crataegus persimilis | NA | Lyonetiidae | Lyonetia clerkella | | 6 August 2022 |
| 8 | Malus × scheideckeri ‘Red Jade’ | EA | Gracillariidae | Callisto denticulella | | 6 August 2022 |
| 9 | Malus × scheideckeri ‘Red Jade’ | EA | Lyonetiidae | Leucoptera malifoliella | | 6 August 2022 |
| 10 | Malus × scheideckeri ‘Red Jade’ | EA | Lyonetiidae | Lyonetia clerkella | | 6 August 2022 |
| 11 | Malus × scheideckeri ‘Red Jade’ | EA | Nepticulidae | Stigmella incognitella | | 6 August 2022 |
| 12 | Malus floribunda | EA | Lyonetiidae | Lyonetia clerkella | | 6 August 2022 |
| 13 | Malus tschonoskii | EA | Lyonetiidae | Lyonetia clerkella | | 6 August 2022 |
| 14 | Phyllostachys viridiglaucescens | EA | Agromyzidae | Cerodontha unisetiorbita | | 9 August 2022 |
| 15 | Prunus incisa ‘February Pink’ | EA | Lyonetiidae | Lyonetia clerkella | | 6 August 2022 |
| 16 | Quercus libani | WA | Gracillariidae | Phyllonorycter distentella | | 9 August 2022 |
| 17 | Quercus libani | WA | Gracillariidae | Caloptilia alchimiella | | 9 August 2022 |
| 18 | Quercus libani | WA | Gracillariidae | Acrocercops brongniardella | | 9 August 2022 |
| 19 | Salix acutifolia | WA | Gracillariidae | Phyllocnistis valentinensis | | 9 August 2022 |
| 20 | Salix acutifolia | WA | Gracillariidae | Phyllocnistis ramulicola |  | 9 August 2022 |
| 21 | Salix acutifolia | WA | Gracillariidae | Phyllonorycter pastorella | | 9 August 2022 |
| 22 | Salix magnifica | EA | Gracillariidae | Phyllonorycter salicicolella | | 6 August 2022 |
| 23 | Viburnum rhytidophyllum | EA | Tortricidae | Acleris schalleriana | | 9 August 2022 |
—low (1%–24% of leaves with mines per 100 leaves), —medium (25%–49%).| Trophic Systems | Indices | ||||
|---|---|---|---|---|---|
| Nestedness | Web Asymmetry | Specialization | Links per Species | Number of Compartments | |
| Insects species—plant species | 6.74 | 0.18 | 0.86 | 1.37 | 24 |
| Insects species—plant genus | 9.40 | 0.45 | 0.98 | 1.03 | 18 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kirichenko, N.I.; Gomboc, S.; Piškur, B.; de Groot, M. Use of an Arboretum and DNA Barcoding for the Detection and Identification of Leaf-Mining Insects on Alien Woody Plants. Forests 2023, 14, 641. https://doi.org/10.3390/f14030641
Kirichenko NI, Gomboc S, Piškur B, de Groot M. Use of an Arboretum and DNA Barcoding for the Detection and Identification of Leaf-Mining Insects on Alien Woody Plants. Forests. 2023; 14(3):641. https://doi.org/10.3390/f14030641
Chicago/Turabian StyleKirichenko, Natalia I., Stanislav Gomboc, Barbara Piškur, and Maarten de Groot. 2023. "Use of an Arboretum and DNA Barcoding for the Detection and Identification of Leaf-Mining Insects on Alien Woody Plants" Forests 14, no. 3: 641. https://doi.org/10.3390/f14030641
APA StyleKirichenko, N. I., Gomboc, S., Piškur, B., & de Groot, M. (2023). Use of an Arboretum and DNA Barcoding for the Detection and Identification of Leaf-Mining Insects on Alien Woody Plants. Forests, 14(3), 641. https://doi.org/10.3390/f14030641