


Contribution of Tree Size and Species on Aboveground Biomass across Land Cover Types in the Taita Hills, Southern Kenya

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Field Inventory Data and Biomass Calculations
2.3. Data Analysis
3. Results
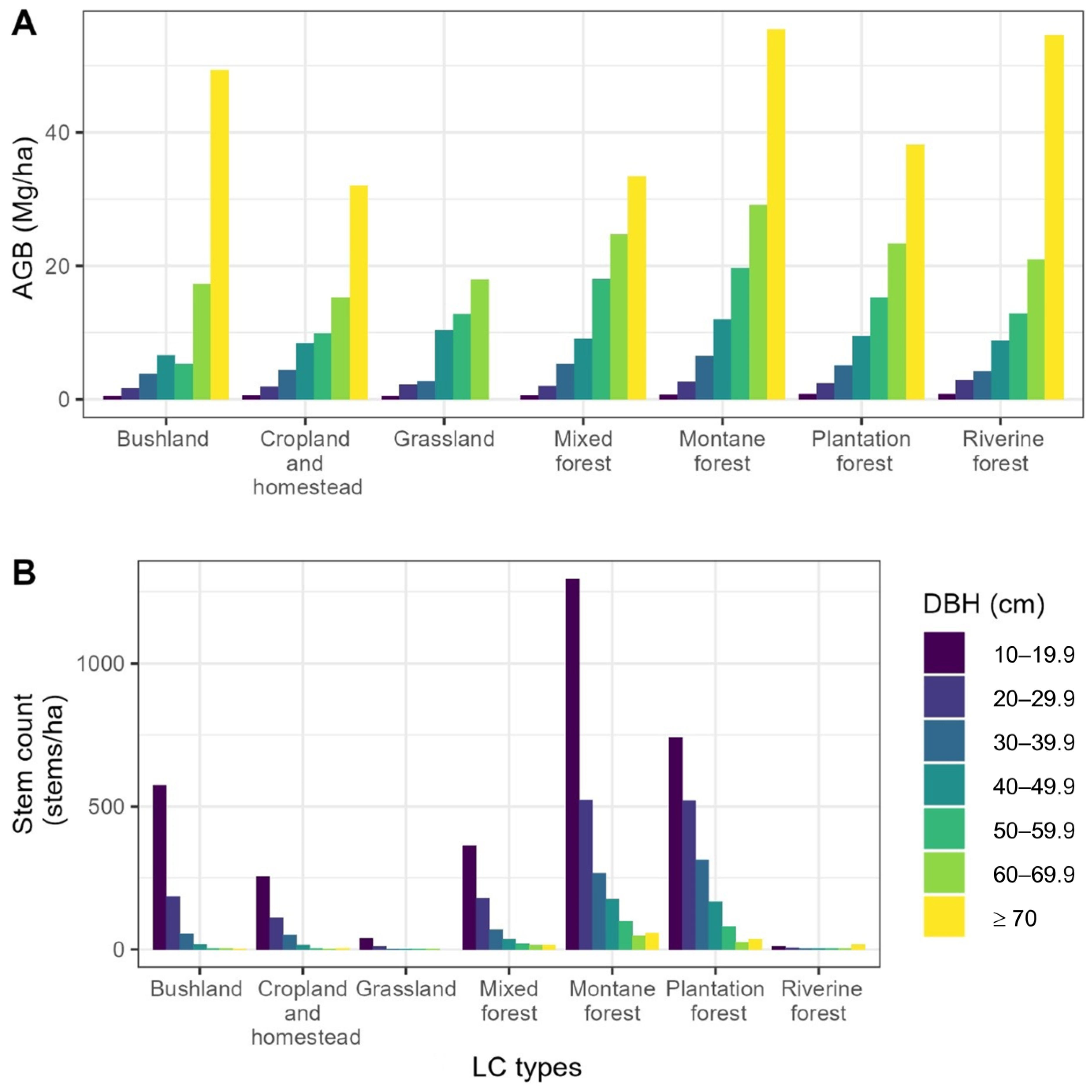
3.1. Contribution of Diameter Classes to Aboveground Biomass
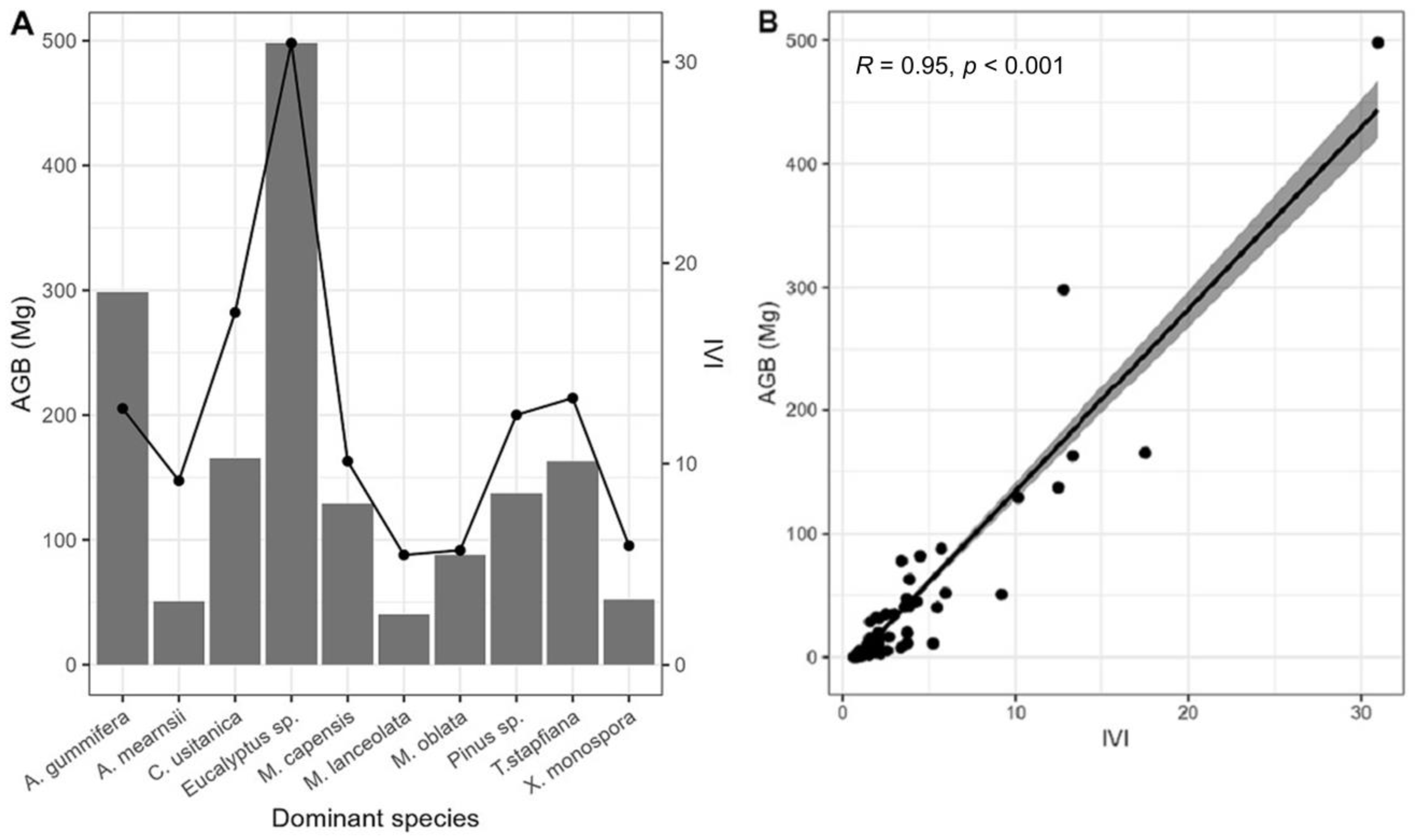
3.2. Contribution of Tree Species on Aboveground Biomass
3.3. Relationship between AGB and IVI of the Dominant Tree Species
4. Discussion
4.1. Importance of Large Trees on Aboveground Biomass
4.2. Most Important Tree Species Contributing to Aboveground Biomass
4.3. Ecological Significance of the Dominant Tree Species
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Duncanson, L.; Armston, J.; Disney, M.; Avitabile, V.; Barbier, N.; Calders, K.; Carter, S.; Chave, J.; Herold, M.; MacBean, N.; et al. Aboveground Woody Biomass Product Validation Good Practices Protocol. Version 1.0. In Good Practices for Satellite Derived Land Product Validation; Duncanson, L., Disney, M., Armston, J., Nickeson, J., Minor, D., Camacho, F., Eds.; Land Product Validation Subgroup (WGCV/CEOS), 2021. Available online: https://lpvs.gsfc.nasa.gov/documents.html (accessed on 15 January 2023).
- Herold, M.; Carter, S.; Avitabile, V.; Espejo, A.B.; Jonckheere, I.; Lucas, R.; McRoberts, R.E.; Næsset, E.; Nightingale, J.; Petersen, R.; et al. The Role and Need for Space-Based Forest Biomass-Related Measurements in Environmental Management and Policy. Surv. Geophys. 2019, 40, 757–778. [Google Scholar] [CrossRef] [Green Version]
- Baccini, A.; Walker, W.; Carvalho, L.; Farina, M.; Sulla-Menashe, D.; Houghton, R.A. Tropical Forests Are a Net Carbon Source Based on Aboveground Measurements of Gain and Loss. Science 2017, 358, 230–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amara, E.; Adhikari, H.; Heiskanen, J.; Siljander, M.; Munyao, M.; Omondi, P.; Pellikka, P. Aboveground Biomass Distribution in a Multi-Use Savannah Landscape in Southeastern Kenya: Impact of Land Use and Fences. Land 2020, 9, 381. [Google Scholar] [CrossRef]
- Dickens, O.; Faith, K.; Geoffrey, M.; Petri, P.; Rob, M. Comparison and Prediction of the above Ground Carbon Storage in Croplands on the Inhabited Slopes on Mount Kilimanjaro (Tanzania) and the Taita Hills (Kenya). J. Geogr. Inf. Syst. 2018, 10, 415–438. [Google Scholar] [CrossRef] [Green Version]
- Slik, J.W.F.; Aiba, S.I.; Brearley, F.Q.; Cannon, C.H.; Forshed, O.; Kitayama, K.; Nagamasu, H.; Nilus, R.; Payne, J.; Paoli, G.; et al. Environmental Correlates of Tree Biomass, Basal Area, Wood Specific Gravity and Stem Density Gradients in Borneo’s Tropical Forests. Glob. Ecol. Biogeogr. 2010, 19, 50–60. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Laurance, W.F.; Franklin, J.F. Global Decline in Large Old Trees. Science 2012, 338, 1305–1306. [Google Scholar] [CrossRef]
- Martin, T.A.; Brown, K.J.; Kučera, J.; Meinzer, F.C.; Sprugel, D.G.; Hinckley, T.M. Control of Transpiration in a 220-Year-Old Abies Amabilis Forest. For. Ecol. Manag. 2001, 152, 211–224. [Google Scholar] [CrossRef]
- Rambo, T.R.; North, M.P. Canopy Microclimate Response to Pattern and Density of Thinning in a Sierra Nevada Forest. For. Ecol. Manag. 2009, 257, 435–442. [Google Scholar] [CrossRef]
- Ali, A.; Wang, L.Q. Big-Sized Trees and Forest Functioning: Current Knowledge and Future Perspectives. Ecol. Indic. 2021, 127, 107760. [Google Scholar] [CrossRef]
- Enquist, B.J.; Abraham, A.J.; Harfoot, M.B.J.; Malhi, Y.; Doughty, C.E. The Megabiota Are Disproportionately Important for Biosphere Functioning. Nat. Commun. 2020, 11, 699. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.; Murphy, H.T. The Importance of Large-Diameter Trees in the Wet Tropical Rainforests of Australia. PLoS ONE 2019, 14, e0208377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mensah, S.; Noulèkoun, F.; Ago, E.E. Aboveground Tree Carbon Stocks in West African Semi-Arid Ecosystems: Dominance Patterns, Size Class Allocation and Structural Drivers. Glob. Ecol. Conserv. 2020, 24, e01331. [Google Scholar] [CrossRef]
- Bastin, J.F.; Barbier, N.; Réjou-Méchain, M.; Fayolle, A.; Gourlet-Fleury, S.; Maniatis, D.; De Haulleville, T.; Baya, F.; Beeckman, H.; Beina, D.; et al. Seeing Central African Forests through Their Largest Trees. Sci. Rep. 2015, 5, 13156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lutz, J.A.; Furniss, T.J.; Johnson, D.J.; Davies, S.J.; Allen, D.; Alonso, A.; Anderson-Teixeira, K.J.; Andrade, A.; Baltzer, J.; Becker, K.M.L.; et al. Global Importance of Large-Diameter Trees. Glob. Ecol. Biogeogr. 2018, 27, 849–864. [Google Scholar] [CrossRef] [Green Version]
- van Wagtendonk, J.W.; Moore, P.E. Fuel Deposition Rates of Montane and Subalpine Conifers in the Central Sierra Nevada, California, USA. For. Ecol. Manag. 2010, 259, 2122–2132. [Google Scholar] [CrossRef]
- Dimobe, K.; Kuyah, S.; Dabré, Z.; Ouédraogo, A.; Thiombiano, A. Diversity-Carbon Stock Relationship across Vegetation Types in W National Park in Burkina Faso. For. Ecol. Manag. 2019, 438, 243–254. [Google Scholar] [CrossRef]
- Atsri, H.K.; Kokou, K.; Abotsi, K.E.; Kokutse, A.D.; Cuni-Sanchez, A. Above-Ground Biomass and Vegetation Attributes in the Forest-Savannah Mosaic of Togo, West Africa. Afr. J. Ecol. 2020, 58, 733–745. [Google Scholar] [CrossRef]
- Mensah, S.; Veldtman, R.; du Toit, B.; Glèlè Kakaï, R.; Seifert, T. Aboveground Biomass and Carbon in a South African Mistbelt Forest and the Relationships with Tree Species Diversity and Forest Structures. Forests 2016, 7, 79. [Google Scholar] [CrossRef] [Green Version]
- Medjibe, V.P.; Putz, F.E.; Starkey, M.P.; Ndouna, A.A.; Memiaghe, H.R. Impacts of Selective Logging on Above-Ground Forest Biomass in the Monts de Cristal in Gabon. For. Ecol. Manag. 2011, 262, 1799–1806. [Google Scholar] [CrossRef]
- Memiaghe, H.R.; Lutz, J.A.; Korte, L.; Alonso, A.; Kenfack, D. Ecological Importance of Small-Diameter Trees to the Structure, Diversity and Biomass of a Tropical Evergreen Forest at Rabi, Gabon. PLoS ONE 2016, 11, e0154988. [Google Scholar] [CrossRef] [Green Version]
- Kreft, H.; Jetz, W. Global Patterns and Determinants of Vascular Plant Diversity. Proc. Natl. Acad. Sci. USA 2007, 104, 5925–5930. [Google Scholar] [CrossRef] [Green Version]
- Bennett, A.C.; Mcdowell, N.G.; Allen, C.D.; Anderson-Teixeira, K.J. Larger Trees Suffer Most during Drought in Forests Worldwide. Nat. Plants 2015, 1, 15139. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Laurance, W.F. The Ecology, Distribution, Conservation and Management of Large Old Trees. Biol. Rev. 2017, 92, 1434–1458. [Google Scholar] [CrossRef]
- Vanak, A.T.; Thaker, M.; Slotow, R. Do Fences Create an Edge-Effect on the Movement Patterns of a Highly Mobile Mega-Herbivore? Biol. Conserv. 2010, 143, 2631–2637. [Google Scholar] [CrossRef]
- Pellegrini, A.F.A.; Pringle, R.M.; Govender, N.; Hedin, L.O. Woody Plant Biomass and Carbon Exchange Depend on Elephant-Fire Interactions across a Productivity Gradient in African Savanna. J. Ecol. 2017, 105, 111–121. [Google Scholar] [CrossRef]
- Slik, J.W.F.; Paoli, G.; Mcguire, K.; Amaral, I.; Barroso, J.; Bastian, M.; Blanc, L.; Bongers, F.; Boundja, P.; Clark, C.; et al. Large Trees Drive Forest Aboveground Biomass Variation in Moist Lowland Forests across the Tropics. Glob. Ecol. Biogeogr. 2013, 22, 1261–1271. [Google Scholar] [CrossRef]
- Mensah, S.; Seifert, T.; Glèlè Kakaï, R. Patterns of Biomass Allocation between Foliage and Woody Structure: The Effects of Tree Size and Specific Functional Traits. Ann. For. Res. 2016, 59, 49–60. [Google Scholar] [CrossRef] [Green Version]
- Lutz, J.A.; Larson, A.J.; Freund, J.A.; Swanson, M.E.; Bible, K.J. The Importance of Large-Diameter Trees to Forest Structural Heterogeneity. PLoS ONE 2013, 8, e0082784. [Google Scholar] [CrossRef] [Green Version]
- Musavi, T.; Migliavacca, M.; Reichstein, M.; Kattge, J.; Wirth, C.; Black, T.A.; Janssens, I.; Knohl, A.; Loustau, D.; Roupsard, O.; et al. Stand Age and Species Richness Dampen Interannual Variation of Ecosystem-Level Photosynthetic Capacity. Nat. Ecol. Evol. 2017, 1, 0048. [Google Scholar] [CrossRef] [Green Version]
- Bordin, K.M.; Esquivel-Muelbert, A.; Bergamin, R.S.; Klipel, J.; Picolotto, R.C.; Frangipani, M.A.; Zanini, K.J.; Cianciaruso, M.V.; Jarenkow, J.A.; Jurinitz, C.F.; et al. Climate and Large-Sized Trees, but Not Diversity, Drive above-Ground Biomass in Subtropical Forests. For. Ecol. Manag. 2021, 490, 119126. [Google Scholar] [CrossRef]
- Keeton, W.S.; Franklin, J.F. Do Remnant Old-Growth Trees Accelerate Rates of Succession in Mature Douglas-Fir Forests? Ecol. Monogr. 2005, 75, 103–118. [Google Scholar] [CrossRef]
- Binkley, D.; Stape, J.L.; Bauerle, W.L.; Ryan, M.G. Explaining Growth of Individual Trees: Light Interception and Efficiency of Light Use by Eucalyptus at Four Sites in Brazil. For. Ecol. Manag. 2010, 259, 1704–1713. [Google Scholar] [CrossRef]
- Das, A.J.; Stephenson, N.L.; Davis, K.P. Why Do Trees Diefl Characterizing the Drivers of Background Tree Mortality. Ecology 2016, 97, 2616–2627. [Google Scholar] [CrossRef]
- Spies, T.A.; Franklin, J.F. The Structure of Natural Young, Mature, and Old-Growth Douglas-Fir Forests in Oregon and Washington. In Wildlife and Vegetation of Unmanaged Douglas-fir Forests; Ruggiero, L.F., Aubry, K.B., Carey, A.B., Huff, M.H., Eds.; U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 1991; pp. 91–109. [Google Scholar]
- Das, A.J.; Larson, A.J.; Lutz, J.A. Individual Species–Area Relationships in Temperate Coniferous Forests. J. Veg. Sci. 2018, 29, 317–324. [Google Scholar] [CrossRef]
- Lutz, J.A.; Larson, A.J.; Furniss, T.J.; Donato, D.C.; Freund, J.A.; Swanson, M.E.; Bible, K.J.; Chen, J.; Franklin, J.F. Equilibrium Pattern in an Old-Growth Pseudotsuga—Tsuga Forest R Eports. Ecology 2014, 95, 2047–2054. [Google Scholar] [CrossRef] [Green Version]
- Reshad, M.; Muhammed, M.; Beyene, A. Distribution and Importance Value Index of Woody Species under Different Successional Stages at Jello-Muktar Dry Afromontane Forest, South-Eastern Ethiopia. Am. J. Agric. For. 2020, 8, 1–8. [Google Scholar] [CrossRef]
- Almazán-Núñez, R.C.; Del Coro Arizmendi, M.; Eguiarte, L.E.; Corcuera, P. Changes in Composition, Diversity and Structure of Woody Plants in Successional Stages of Tropical Dry Forest in Southwest Mexico. Rev. Mex. Biodivers 2012, 83, 1096–1109. [Google Scholar] [CrossRef] [Green Version]
- Platts, P.J.; Burgess, N.D.; Gereau, R.E.; Lovett, J.C.; Marshall, A.R.; McClean, C.J.; Pellikka, P.K.E.; Swetnam, R.D.; Marchant, R. Delimiting Tropical Mountain Ecoregions for Conservation. Environ. Conserv. 2011, 38, 312–324. [Google Scholar] [CrossRef] [Green Version]
- Sombroek, W.G.; Braun, H.M.H.; van der Pouw, B.J. Exploratory Soil Map and Agro-Climatic Zone Map of Kenya; Republic of Kenya, Ministry of Agriculture: Nairobi, Kenya, 1982; p. 60. [Google Scholar]
- Autio, A.E. Transnational Road Project and Regional Development: Expectations of Regional Administration and Realized Impacts in Taita Taveta County, Kenya. Master’s Thesis, Department of Geosciences and Geography, University of Helsinki, Helsinki, Finland, 2018. [Google Scholar]
- Cuni-Sanchez, A.; Pfeifer, M.; Marchant, R.; Calders, K.; Sørensen, C.L.; Pompeu, P.V.; Lewis, S.L.; Burgess, N.D. New Insights on above Ground Biomass and Forest Attributes in Tropical Montane Forests. For. Ecol. Manag. 2017, 399, 235–246. [Google Scholar] [CrossRef] [Green Version]
- Pellikka, P.K.E.; Clark, B.J.F.; Gosa, A.G.; Himberg, N.; Hurskainen, P.; Maeda, E.; Mwang’ombe, J.; Omoro, L.M.A.; Siljander, M. Agricultural Expansion and Its Consequences in the Taita Hills, Kenya. Dev. Earth Surf. Process. 2013, 16, 165–179. [Google Scholar] [CrossRef] [Green Version]
- Adhikari, H.; Heiskanen, J.; Siljander, M.; Maeda, E.; Heikinheimo, V.; Pellikka, P.K.E. Determinants of Aboveground Biomass across an Afromontane Landscape Mosaic in Kenya. Remote Sens. 2017, 9, 827. [Google Scholar] [CrossRef] [Green Version]
- Pellikka, P.K.E.E.; Heikinheimo, V.; Hietanen, J.; Schäfer, E.; Siljander, M.; Heiskanen, J. Impact of Land Cover Change on Aboveground Carbon Stocks in Afromontane Landscape in Kenya. Appl. Geogr. 2018, 94, 178–189. [Google Scholar] [CrossRef]
- Heiskanen, J.; Pellikka, P.K.E.; Aynekulu, E.; Packalen, P. Field Measurement Guidelines for Aboveground Biomass and Fuel Wood Stocks. In Building Biocarbon and Rural Development in West Africa (BIODEV); World Agroforestry Centre: Nairobi, Kenya, 2013. [Google Scholar]
- Heiskanen, J.; Adhikari, H.; Piiroinen, R.; Packalen, P.; Pellikka, P.K.E. Do Airborne Laser Scanning Biomass Prediction Models Benefit from Landsat Time Series, Hyperspectral Data or Forest Classification in Tropical Mosaic Landscapes? Int. J. Appl. Earth Obs. Geoinf. 2019, 81, 176–185. [Google Scholar] [CrossRef]
- Curtis, R.O. Height-Diameter and Height-Diameter-Age Equations for Second-Growth Douglas Fir. Forest Science 1967, 13, 365–375. [Google Scholar]
- Valbuena, R.; Heiskanen, J.; Aynekulu, E.; Pitkänen, S.; Packalen, P. Sensitivity of Above-Ground Biomass Estimates to Height-Diameter Modelling in Mixed-Species West African Woodlands. PLoS ONE 2016, 11, e0158198. [Google Scholar] [CrossRef] [Green Version]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D. Nlme: Linear and Nonlinear Mixed Effects Models. R Package Version 3.1-152; 2021; Available online: https://CRAN.R-project.org/package=nlme (accessed on 15 January 2023).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Chave, J.; Réjou-Méchain, M.; Búrquez, A.; Chidumayo, E.; Colgan, M.S.; Delitti, W.B.C.; Duque, A.; Eid, T.; Fearnside, P.M.; Goodman, R.C.; et al. Improved Allometric Models to Estimate the Aboveground Biomass of Tropical Trees. Glob. Chang. Biol. 2014, 20, 3177–3190. [Google Scholar] [CrossRef]
- Paul, K.I.; Roxburgh, S.H.; England, J.R.; Ritson, P.; Hobbs, T.; Brooksbank, K.; John Raison, R.; Larmour, J.S.; Murphy, S.; Norris, J.; et al. Development and Testing of Allometric Equations for Estimating Above-Ground Biomass of Mixed-Species Environmental Plantings. For. Ecol. Manag. 2013, 310, 483–494. [Google Scholar] [CrossRef]
- Henry, M.; Picard, N.; Trotta, C.; Manlay, R.J.; Bernoux, M.; Saint-andré, L.; Henry, M.; Picard, N.; Trotta, C.; Manlay, R.J.; et al. Estimating Tree Biomass of Sub-Saharan African Forests: A Review of Available Allometric Equations to Cite This Version: HAL Id: Hal-02651041 Estimating Tree Biomass of Sub-Saharan African Forests: A Review of Available Allometric Equations. Silva Fenn. 2011, 45, 477–596. [Google Scholar] [CrossRef] [Green Version]
- Penman, J.; Gytarsky, M.; Hiraishi, T.; Krug, T.; Kruger, D.; Pipatti, R. (Eds.) Good Practice Guidance for Land Use, Land-Use Change and Forestry; IPCC National Greenhouse Gas Inventories Programme and Institute for Global Environmental Strategies: Kanagawa, Japan, 2003. [Google Scholar]
- Réjou-Méchain, M.; Tanguy, A.; Piponiot, C.; Chave, J.; Hérault, B. Biomass: An R Package for Estimating Above-Ground Biomass and Its Uncertainty in Tropical Forests. Methods Ecol. Evol. 2017, 8, 1163–1167. [Google Scholar] [CrossRef] [Green Version]
- Cottam, G.; Curtis, J.T. The Use of Distance Measures in Phytosociological Sampling. Ecology 1956, 37, 451–460. [Google Scholar] [CrossRef]
- Nguyen, H.; Lamb, D.; Herbohn, J.; Firn, J. Designing Mixed Species Tree Plantations for the Tropics: Balancing Ecological Attributes of Species with Landholder Preferences in the Philippines. PLoS ONE 2014, 9, e0095267. [Google Scholar] [CrossRef]
- Nebel, G.; Kvist, L.P.; Vanclay, J.K.; Christensen, H.; Freitas, L.; Ruíz, J. Structure and Floristic Composition of Flood Plain Forests in the Peruvian Amazon I. Overstorey. For. Ecol. Manag. 2001, 150, 27–57. [Google Scholar] [CrossRef] [Green Version]
- Guariguata, M.R.; Chazdon, R.L.; Denslow, J.S.; Dupuy, J.M.; Anderson, L. Structure and Floristics of Secondary and Old-Growth Forest Stands in Lowland Costa Rica. Plant Ecol. 1997, 132, 107–120. [Google Scholar] [CrossRef]
- Lahoti, S.; Lahoti, A.; Joshi, R.K.; Saito, O. Vegetation Structure, Species Composition, and Carbon Sink Potential of Urban Green Spaces in Nagpur City, India. Land 2020, 9, 107. [Google Scholar] [CrossRef] [Green Version]
- Cuni-Sanchez, A.; Sullivan, M.J.P.; Platts, P.J.; Lewis, S.L.; Marchant, R.; Imani, G.; Hubau, W.; Abiem, I.; Adhikari, H.; Albrecht, T.; et al. High Aboveground Carbon Stock of African Tropical Montane Forests. Nature 2021, 596, 536–542. [Google Scholar] [CrossRef] [PubMed]
- Mensah, S.; Salako, V.K.; Seifert, T. Structural Complexity and Large-Sized Trees Explain Shifting Species Richness and Carbon Relationship across Vegetation Types. Funct. Ecol. 2020, 34, 1731–1745. [Google Scholar] [CrossRef]
- Woollen, E.; Ryan, C.M.; Williams, M. Carbon Stocks in an African Woodland Landscape: Spatial Distributions and Scales of Variation. Ecosystems 2012, 15, 804–818. [Google Scholar] [CrossRef]
- Clark, D.B.; Clark, D.A. Abundance, Growth and Mortality of Very Large Trees in Neotropical Lowland Rain Forest. For. Ecol. Manag. 1996, 80, 235–244. [Google Scholar] [CrossRef]
- Wekesa, C.; Leley, N.; Maranga, E.; Kirui, B.; Muturi, G.; Mbuvi, M.; Chikamai, B. Effects of Forest Disturbance on Vegetation Structure and Above-Ground Carbon in Three Isolated Forest Patches of Taita Hills. Open J. For. 2016, 06, 142–161. [Google Scholar] [CrossRef] [Green Version]
- Rogo, L.; Oguge, N. The Taita Hill Forest Remnants: A Disappearing World Heritage. Ambio 2000, 29, 522–525. [Google Scholar] [CrossRef]
- Zschauer, K. Households Energy Supply and the Use of Fuelwood in the Taita Hills, Kenya. Master’s Thesis, Department of Geosciences and Geography, University of Helsinki, Helsinki, Finland, 2012. [Google Scholar]
- Pokhrel, S.; Sherpa, C. Analyzing the Relationship, Distribution of Tree Species Diversity, and Above-Ground Biomass on the Chitwan-Annapurna Landscape in Nepal. Int. J. For. Res. 2020, 2020, 2789753. [Google Scholar] [CrossRef]
- Buma, B.; Krapek, J.; Edwards, R.T. Watershed-Scale Forest Biomass Distribution in a Perhumid Temperate Rainforest as Driven by Topographic, Soil, and Disturbance Variables. Can. J. For. Res. 2016, 46, 844–854. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Guan, D.S.; Song, M.W. Biomass and Carbon Storage of Eucalyptus and Acacia Plantations in the Pearl River Delta, South China. For. Ecol. Manag. 2012, 277, 90–97. [Google Scholar] [CrossRef]
- Moore, S.; Adu-Bredu, S.; Duah-Gyamfi, A.; Addo-Danso, S.D.; Ibrahim, F.; Mbou, A.T.; de Grandcourt, A.; Valentini, R.; Nicolini, G.; Djagbletey, G.; et al. Forest Biomass, Productivity and Carbon Cycling along a Rainfall Gradient in West Africa. Glob. Chang. Biol. 2018, 24, e496–e510. [Google Scholar] [CrossRef] [PubMed]
- Olorunfemi, I.E.; Komolafe, A.A.; Fasinmirin, J.T.; Olufayo, A.A. Biomass Carbon Stocks of Different Land Use Management in the Forest Vegetative Zone of Nigeria. Acta Oecologica 2019, 95, 45–56. [Google Scholar] [CrossRef]
- Guldemond, R.; Van Aarde, R. A Meta-Analysis of the Impact of African Elephants on Savanna Vegetation. J. Wildl. Manag. 2008, 72, 892–899. [Google Scholar] [CrossRef]
- Thomas, N.; Baltezar, P.; Lagomasino, D.; Stovall, A.; Iqbal, Z.; Fatoyinbo, L. Trees Outside Forests Are an Underestimated Resource in a Country with Low Forest Cover. Sci. Rep. 2021, 11, 7919. [Google Scholar] [CrossRef]
- Kehlenbeck, K.; Arifin, H.S.; Maass, B.L. Plant Diversity in Homegardens in a Socio-Economic and Agro-Ecological Context. In Stability of Tropical Rainforest Margins; Springer: Berlin/Heidelberg, Germany, 2007; pp. 295–317. [Google Scholar] [CrossRef]
- Yirga, A.; Addisu Legesse, S.; Mekuriaw, A. Carbon Stock and Mitigation Potentials of Zeghie Natural Forest for Climate Change Disaster Reduction, Blue Nile Basin, Ethiopia. Earth Syst. Environ. 2020, 4, 27–41. [Google Scholar] [CrossRef]
- Kendie, G.; Addisu, S.; Abiyu, A. Biomass and Soil Carbon Stocks in Different Forest Types, Northwestern Ethiopia. Int. J. River Basin Manag. 2021, 19, 123–129. [Google Scholar] [CrossRef]
- Henry, M.; Tittonell, P.; Manlay, R.J.; Bernoux, M.; Albrecht, A.; Vanlauwe, B. Biodiversity, Carbon Stocks and Sequestration Potential in Aboveground Biomass in Smallholder Farming Systems of Western Kenya. Agric. Ecosyst. Environ. 2009, 129, 238–252. [Google Scholar] [CrossRef]
- Stromgaard, P. Biomass, Growth, and Burning of Woodland in a Shifting Cultivation Area of South Central Africa. For. Ecol. Manag. 1985, 12, 163–178. [Google Scholar] [CrossRef]
- Orwa, A.E. Acacia Mearnsii De Wild. Fabaceae—Mimosoideae (Black Wattle). Agroforestry Database 4.0. 2009, 0, 1–5. Available online: https://apps.worldagroforestry.org/treedb/AFTPDFS/Acacia_mearnsii.PDF (accessed on 15 January 2023).
- Omoro, L.M.A.; Starr, M.; Pellikka, P.K.E. Tree Biomass and Soil Carbon Stocks in Indigenous Forests in Comparison to Plantations of Exotic Species in the Taita Hills of Kenya. Silva Fenn. 2013, 47, 935. [Google Scholar] [CrossRef] [Green Version]
- Piiroinen, R.; Fassnacht, F.E.; Heiskanen, J.; Maeda, E.; Mack, B.; Pellikka, P. Invasive tree species detection in the Eastern Arc Mountains biodiversity hotspot using one class classification. Remote Sens. Environ. 2018, 218, 119–131. [Google Scholar] [CrossRef]
- Kluthe, B.G.; Chen, D. Eucalyptus Sp. at the Intersection of Environment and Culture in Kenya. Ethnobiol. Lett. 2017, 8, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Bennett, B.M. The El Dorado of Forestry: The Eucalyptus in India, South Africa, and Thailand, 1850–2000. Int. Rev. Soc. Hist. 2010, 55, 27–50. [Google Scholar] [CrossRef] [Green Version]
- Servie, K.F. A Guide to On-Farm Eucalyptus Growing in Kenya; Kenya Forest Service: Nairobi, Kenya, 2009; pp. 1–36. Available online: https://www.bebuffered.com/downloads/Kenya_Forest_Service_Eucalyptus_Guidelines.pdf (accessed on 15 January 2023).
- Oballa, P.O.; Konuche, P.K.A.; Muchiri, M.N.; Kigomo, B.N. Facts on Growing and Use of Eucalyptus in Kenya; Kenya Forestry Research Institute: Nairobi, Kenya, 2010. [Google Scholar]
- Kalibo, H.W. A Participatory Assessment of Forest Resource Use at Mt Kasigau, Kenya; Miami University: Oxford, OH, USA, 2004. [Google Scholar]
- Williams, M.; Ryan, C.M.; Rees, R.M.; Sambane, E.; Fernando, J.; Grace, J. Carbon Sequestration and Biodiversity of Re-Growing Miombo Woodlands in Mozambique. For. Ecol. Manag. 2008, 254, 145–155. [Google Scholar] [CrossRef]
- Schoeneberger, M.M. Agroforestry: Working Trees for Sequestering Carbon on Agricultural Lands. Agrofor. Syst. 2009, 75, 27–37. [Google Scholar] [CrossRef] [Green Version]
- Kuyah, S.; Sileshi, G.W.; Njoloma, J.; Mng’omba, S.; Neufeldt, H. Estimating Aboveground Tree Biomass in Three Different Miombo Woodlands and Associated Land Use Systems in Malawi. Biomass Bioenergy 2014, 66, 214–222. [Google Scholar] [CrossRef]
- Montagnini, F.; Nair, P.K.R. Carbon Sequestration: An Underexploited Environmental Benefit of Agroforestry Systems. In New Vistas in Agroforestry; Springer: Dordrecht, The Netherlands, 2004; pp. 281–295. [Google Scholar] [CrossRef]
- Abbot, J.I.O.; Mace, R. Managing Protected Woodlands: Fuelwood Collection and Law Enforcement in Lake Malawi National Park. Conserv. Biol. 1999, 13, 418–421. [Google Scholar] [CrossRef]
- Muchiri, M.N. Grevellia Robusta in Agroforestry Systems in Kenya. J. Trop. For. Sci. 2004, 16, 396–401. [Google Scholar]
- Michael Lock, J. Ecosystems of Africa. In Encyclopedia of Biodiversity, 2nd ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2013; Volume 1, pp. 45–57. [Google Scholar] [CrossRef]
- Mengich, E.K.; Macharia, J.M.; Mitloehner, R.; Too, D.K.; Muturi, G.M. Diameter Distribution of Indigenous Trees as Indicator of Adapted Species in Semi-Arid Rangelands of Kenya. J. Trop. For. Sci. 2020, 32, 144–153. [Google Scholar] [CrossRef]
- Singh, G. Growth, Biomass Production, and Soil Water Dynamics in Acacia Tortilis Plantation in Relation to Microhabitat and Surface Vegetation in the Hot Arid Region of Indian Desert. Arid. Land Res. Manag. 2004, 18, 153–169. [Google Scholar] [CrossRef]
- Kituyi, E.; Marufu, L.; Wandiga, S.O.; Jumba, I.O.; Andreae, M.O.; Helas, G. Biofuel Availability and Domestic Use Patterns in Kenya. Biomass Bioenergy 2001, 20, 71–82. [Google Scholar] [CrossRef]
- Alemagi, D.; Duguma, L.; Minang, P.A.; Nkeumoe, F.; Feudjio, M.; Tchoundjeu, Z. Intensification of Cocoa Agroforestry Systems as a REDD+ Strategy in Cameroon: Hurdles, Motivations, and Challenges. Int. J. Agric. Sustain. 2014, 13, 187–203. [Google Scholar] [CrossRef]
- Dessie1, G.; Wolka, K. Teklu Erkossa Effects of Albizia gummifera, Millettia ferruginea and Cordia africana Leaf Litter on the Germination of Coffea arabica L. Seed. Int. J. Agric. Res. 2012, 7, 315–323. [Google Scholar]
- Bogale, T.; Datiko, D.; Belachew, S. Structure and Natural Regeneration Status of Woody Plants of Berbere Afromontane Moist Forest, Bale Zone, South East Ethiopia; Implication to Biodiversity Conservation. Open J. For. 2017, 7, 352–371. [Google Scholar] [CrossRef]
- Gelasso, M.; Li, J. Structure and Regeneration Status of Woody Species in the Munessa Forest, Southern Ethiopia. J. For. Res. 2021, 32, 493–501. [Google Scholar] [CrossRef]
- Takaoka, S. Long-Term Growth Performance of Cordia Africana and Grevillea Robusta Trees in the Mount Kenya Region. Agrofor. Syst. 2008, 72, 169–172. [Google Scholar] [CrossRef]
- Torquebiau, E.F. A Renewed Perspective on Agroforestry Concepts and Classification. Comptes Rendus De L’academie Des Sci.—Serie III 2000, 323, 1009–1017. [Google Scholar] [CrossRef]
- Tshering, S. Importance Value Index and Assessment of Carbon Stocks in Western Bhutan Himalaya (Thimphu). Curr. J. Appl. Sci. Technol. 2019, 32, 1–8. [Google Scholar] [CrossRef]
- Belsky, A.J. Influences of Trees on Savanna Productivity: Tests of Shade, Nutrients, and Tree-Grass Competition. Ecology 1994, 75, 922–932. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LC Type | Description | n | Number of Tree Species | Elevation (m) |
|---|---|---|---|---|
| Montane forest | Montane forest of native tree species. | 39 | 74 | 1506–2147 |
| Plantation forest | Plantation forest of exotic tree species (Eucalyptus spp., Cupressus lusitanica, Pinus spp.) | 39 | 30 | 1407–2133 |
| Mixed forest | Montane or plantation forest with mixed species composition (<80% of the species are either native or exotic). | 11 | 38 | 1641–2136 |
| Riverine forest | Forest and woodland along the river. | 7 | 4 | 859–887 |
| Bushland | Acacia-Commiphora bushland and thicket in the plains and lower height Acacia mearnsii plantations in hills. | 70 | 70 | 675–1987 |
| Grassland | Open savanna grassland with scattered trees. | 9 | 8 | 880–1049 |
| Cropland and homestead | Open crop fields, agroforestry systems, and homestead with woody vegetation. | 64 | 62 | 669–1765 |
| LC Type | Variable | Mean | SD | Range |
|---|---|---|---|---|
| Montane forest (n = 39) | Density (trees ha−1) | 664.0 | 220.2 | 150.0–1214.0 |
| Basal area (m2 ha−1) | 44.9 | 14.9 | 16.9–85.6 | |
| Mean height (m) | 13.8 | 2.2 | 8.2–18.2 | |
| Mean DBH (cm) | 24.7 | 15.8 | 10.0–123.5 | |
| AGB (Mg ha−1) | 323.6 | 134.2 | 117.3–643.2 | |
| Plantation forest (n = 39) | Density (trees ha−1) | 505.0 | 236.6 | 120.0–1192.8 |
| Basal area (m2 ha−1) | 36.6 | 19.8 | 5.1–94.3 | |
| Mean height (m) | 16.4 | 4.4 | 10.1–29.7 | |
| Mean DBH (cm) | 26.8 | 14.7 | 10.0–118.5 | |
| AGB (Mg ha−1) | 211.4 | 131.7 | 21.5–671.1 | |
| Mixed forest (n = 11) | Density (trees ha−1) | 646.0 | 200.9 | 411.3–1060.0 |
| Basal area (m2 ha−1) | 40.1 | 13.3 | 16.6–555.5 | |
| Mean height (m) | 11.9 | 2.8 | 7.7–16.6 | |
| Mean DBH (cm) | 23.7 | 14.9 | 10.0–110.1 | |
| AGB (Mg ha−1) | 223.3 | 95.5 | 85.4–388.4 | |
| Riverine forest (n = 7) | Density (trees ha−1) | 69.0 | 58.1 | 10.0–190.7 |
| Basal area (m2 ha−1) | 21.7 | 12.6 | 61.0–42.7 | |
| Mean height (m) | 16.2 | 5.0 | 9.6–24.1 | |
| Mean DBH (cm) | 53.7 | 34.0 | 10.6–170.0 | |
| AGB (Mg ha−1) | 155.7 | 121.2 | 25.9–379.1 | |
| Bushland (n = 70) | Density (trees ha−1) | 123.0 | 145.0 | 10.0–960.0 |
| Basal area (m2 ha−1) | 4.2 | 4.8 | 0.1–23.6 | |
| Mean height (m) | 7.4 | 2.3 | 3.0–13.5 | |
| Mean DBH (cm) | 18.6 | 9.1 | 10.0–109.0 | |
| AGB (Mg ha−1) | 15.9 | 23.7 | 0.1–127.1 | |
| Grassland (n = 9) | Density (trees ha−1) | 59.0 | 53.0 | 20.3–171.3 |
| Basal area (m2 ha−1) | 2.4 | 2.1 | 0.4–7.3 | |
| Mean height (m) | 8.4 | 1.3 | 6.8–11.2 | |
| Mean DBH (cm) | 19.4 | 12.2 | 10.1–63.5 | |
| AGB (Mg ha−1) | 11.6 | 12.5 | 1.9–42.3 | |
| Cropland and homestead (n = 64) | Density (trees ha−1) | 64.0 | 72.0 | 10.0–240 |
| Basal area (m2 ha−1) | 2.4 | 2.1 | 0.4–7.3 | |
| Mean height (m) | 8.4 | 1.3 | 3.0–20.3 | |
| Mean DBH (cm) | 21.5 | 11.8 | 10.0–100.7 | |
| AGB (Mg ha−1) | 15.0 | 21.8 | 0.2–108.9 |
| LC Type | Species | Contribution to AGB (%) | Contribution to AGB (%) of DBH Class (cm) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 10–19.9 | 20–29.9 | 30–39.9 | 40–49.9 | 50–59.9 | 60–69.9 | ≥70 | |||
| Bushland | Acacia mearnsii | 22.3 | 32.7 | 28.8 | 8.9 | 12.9 | 22.4 | ||
| Albizia amara | 17.6 | 2.5 | 12.1 | 30.1 | 40.2 | 37.4 | 41.0 | ||
| Newtonia hildebrandtii | 4.6 | 100.0 | |||||||
| Commiphora africana | 4.6 | 3.3 | 5.9 | 5.5 | 46.1 | ||||
| Agauria salicifolia | 4.3 | 2.8 | 9.9 | 22.1 | |||||
| Cropland and homestead | Grevillea robusta | 25.2 | 8.6 | 21.4 | 39.2 | 57.6 | 47 | ||
| Eucalyptus sp. | 12.1 | 6.1 | 4.9 | 16.8 | 16.8 | 29.2 | 76.7 | ||
| Mangifera indica | 11.6 | 5.7 | 3.4 | 3.6 | 5.3 | 49.4 | |||
| Ficus thonningii | 4.5 | 13.1 | 23.3 | 18.4 | |||||
| Persea americana | 4.4 | 4.3 | 7.1 | 6.9 | 10.7 | ||||
| Grassland | Vachellia tortillis | 63.9 | 38.1 | 75.6 | 100.0 | 100.0 | 53.1 | ||
| Acacia reficiens | 16.1 | 46.9 | |||||||
| Portulacaria afra | 8.1 | 35.6 | 5.0 | ||||||
| Albizia anthelmintica | 7.6 | 4.4 | 19.4 | 100.0 | |||||
| Acacia senegal | 1.8 | 9.3 | |||||||
| Mixed forest | Albizia gummifera | 18.3 | 5.5 | 5.5 | 14 | 16.3 | 23.2 | 29.4 | 27.2 |
| Eucalyptus sp. | 11.5 | 7.9 | 5.6 | 5.8 | 8.6 | 9.3 | 14.5 | 22.8 | |
| Cupressus lusitanica | 9.7 | 4.4 | 8.0 | 6.6 | 28.1 | 9.8 | 9.4 | 4.5 | |
| Xymalos monospora | 6.8 | 5.7 | 8.0 | 5.8 | 4.1 | 4.9 | 14.6 | ||
| Nuxia floribunda | 6.6 | 1.4 | 4.7 | 8.4 | 18.6 | 5.6 | 6.1 | ||
| Montane forest | Albizia gummifera | 21.1 | 2.6 | 3.8 | 10.0 | 7.5 | 13.9 | 20.1 | 54.3 |
| Tabernaemontana stapfiana | 12.9 | 12.3 | 20.3 | 19.7 | 17.4 | 17.1 | 9.0 | 2.2 | |
| Macaranga capensis | 9.3 | 5.0 | 6.7 | 7.0 | 10.5 | 15.1 | 16.8 | 5.3 | |
| Millettia oblata | 6.7 | 6.2 | 7.1 | 11.6 | 13.4 | 4.7 | 3.8 | ||
| Syzygium guineense | 5.9 | 1.4 | 3.4 | 2.2 | 6.8 | 9.0 | 11.7 | 5.4 | |
| Plantation | Eucalyptus sp. | 57.2 | 43.6 | 45.9 | 41.0 | 47.3 | 55.3 | 79.1 | 93.9 |
| forest | Cupressus lusitanica | 17.5 | 14.2 | 22.8 | 24.0 | 27.7 | 16.9 | 8.0 | |
| Pinus sp. | 16.7 | 5.0 | 16.9 | 27.1 | 20.3 | 17.8 | 12.9 | 6.1 | |
| Acacia mearnsii | 2.5 | 14.6 | 6.1 | 2.0 | 1.0 | ||||
| Grevillea robusta | 1.0 | 0.1 | 0.6 | 2.8 | 2.1 | ||||
| Riverine | Newtonia hildebrandtii | 67.2 | 72.2 | 39.6 | 15.2 | 77.6 | 72.7 | ||
| forest | Vachellia xanthophloea | 31.7 | 100.0 | 75.5 | 27.8 | 29.4 | 84.8 | 22.4 | 27.3 |
| Cordia goetzei | 0.8 | 31.1 | |||||||
| Trichilia lepidota | 0.4 | 24.5 | |||||||
| LC Type | Species | RF | RDE | RDO | IVI | % IVI | % AGB |
|---|---|---|---|---|---|---|---|
| Bushland | Acacia mearnsii | 4.7 | 21.1 | 17.6 | 43.3 | 14.4 | 22.7 |
| Commiphora africana | 6.1 | 9.0 | 10.2 | 25.3 | 8.4 | 4.5 | |
| Commiphora schimperi | 4.7 | 9.3 | 4.9 | 18.9 | 6.3 | 2.9 | |
| Albizia amara | 3.3 | 5.0 | 10.4 | 18.7 | 6.3 | 1.8 | |
| Cropland and homestead | Grevillea robusta | 10.4 | 13.9 | 20.6 | 44.9 | 15.0 | 26 |
| Mangifera indica | 4.6 | 9.7 | 10.8 | 25.1 | 8.4 | 11 | |
| Persea americana | 8.7 | 5.9 | 6.4 | 21.0 | 7.0 | 1.5 | |
| Eucalyptus spp. | 2.9 | 5.0 | 8.8 | 16.6 | 5.5 | 13 | |
| Grassland | Acacia tortilis | 35.7 | 37.7 | 56.0 | 129.5 | 43.0 | 64 |
| Portulacaria afra | 7.1 | 30.2 | 12.9 | 50.3 | 16.8 | 8.1 | |
| Albizia anthelmintica | 21.4 | 13.2 | 12.4 | 47.1 | 15.7 | 7.6 | |
| Acacia reficiens | 7.1 | 1.9 | 13.0 | 22.1 | 7.4 | 16 | |
| Mixed forest | Albizia gummifera | 5.8 | 9.0 | 17.2 | 31.9 | 10.4 | 18.5 |
| Cupressus lusitanica | 6.7 | 9.3 | 13.0 | 28.9 | 9.6 | 10 | |
| Maesa lanceolata | 9.6 | 10.6 | 4.8 | 25.0 | 8.3 | 4.5 | |
| Eucalyptus spp. | 4.8 | 5.7 | 9.1 | 19.6 | 9.9 | 11.2 | |
| Montane forest | Tabernaemontana stapfiana | 5.8 | 14.6 | 14.4 | 34.7 | 11.7 | 13 |
| Albizia gummifera | 5.2 | 5.6 | 16.7 | 27.6 | 9.2 | 21 | |
| Macaranga capensis | 4.9 | 8.1 | 10.9 | 23.9 | 8.0 | 9 | |
| Millettia oblata | 3.9 | 5.6 | 5.7 | 15.3 | 5.1 | 7 | |
| Plantation forest | Eucalyptus spp. | 18.3 | 38.8 | 49.9 | 107.0 | 35.7 | 58 |
| Cupressus lusitanica | 11.1 | 23.2 | 21.6 | 55.9 | 18.6 | 18 | |
| Pinus spp. | 8.7 | 15.9 | 20.6 | 45.2 | 15.4 | 17 | |
| Acacia mearnsii | 12.7 | 7.9 | 3.1 | 23.7 | 7.9 | 3.0 | |
| Riverine forest | Newtonia hildebrandtii | 50.0 | 37.5 | 61.6 | 149.1 | 49.7 | 67 |
| Vachellia xanthophloea | 30.0 | 58.3 | 37.0 | 125.3 | 41.8 | 32 | |
| Cordia goetzei | 10.0 | 2.1 | 1.0 | 13.1 | 4.4 | 0.7 | |
| Trichilia lepidota | 10.0 | 2.1 | 0.4 | 12.5 | 4.1 | 0.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amara, E.; Adhikari, H.; Mwamodenyi, J.M.; Pellikka, P.K.E.; Heiskanen, J. Contribution of Tree Size and Species on Aboveground Biomass across Land Cover Types in the Taita Hills, Southern Kenya. Forests 2023, 14, 642. https://doi.org/10.3390/f14030642
Amara E, Adhikari H, Mwamodenyi JM, Pellikka PKE, Heiskanen J. Contribution of Tree Size and Species on Aboveground Biomass across Land Cover Types in the Taita Hills, Southern Kenya. Forests. 2023; 14(3):642. https://doi.org/10.3390/f14030642
Chicago/Turabian StyleAmara, Edward, Hari Adhikari, James M. Mwamodenyi, Petri K. E. Pellikka, and Janne Heiskanen. 2023. "Contribution of Tree Size and Species on Aboveground Biomass across Land Cover Types in the Taita Hills, Southern Kenya" Forests 14, no. 3: 642. https://doi.org/10.3390/f14030642
APA StyleAmara, E., Adhikari, H., Mwamodenyi, J. M., Pellikka, P. K. E., & Heiskanen, J. (2023). Contribution of Tree Size and Species on Aboveground Biomass across Land Cover Types in the Taita Hills, Southern Kenya. Forests, 14(3), 642. https://doi.org/10.3390/f14030642