
Recent Trends and Future Challenges for Lichen Biomonitoring in Forests



Abstract
:1. Introduction
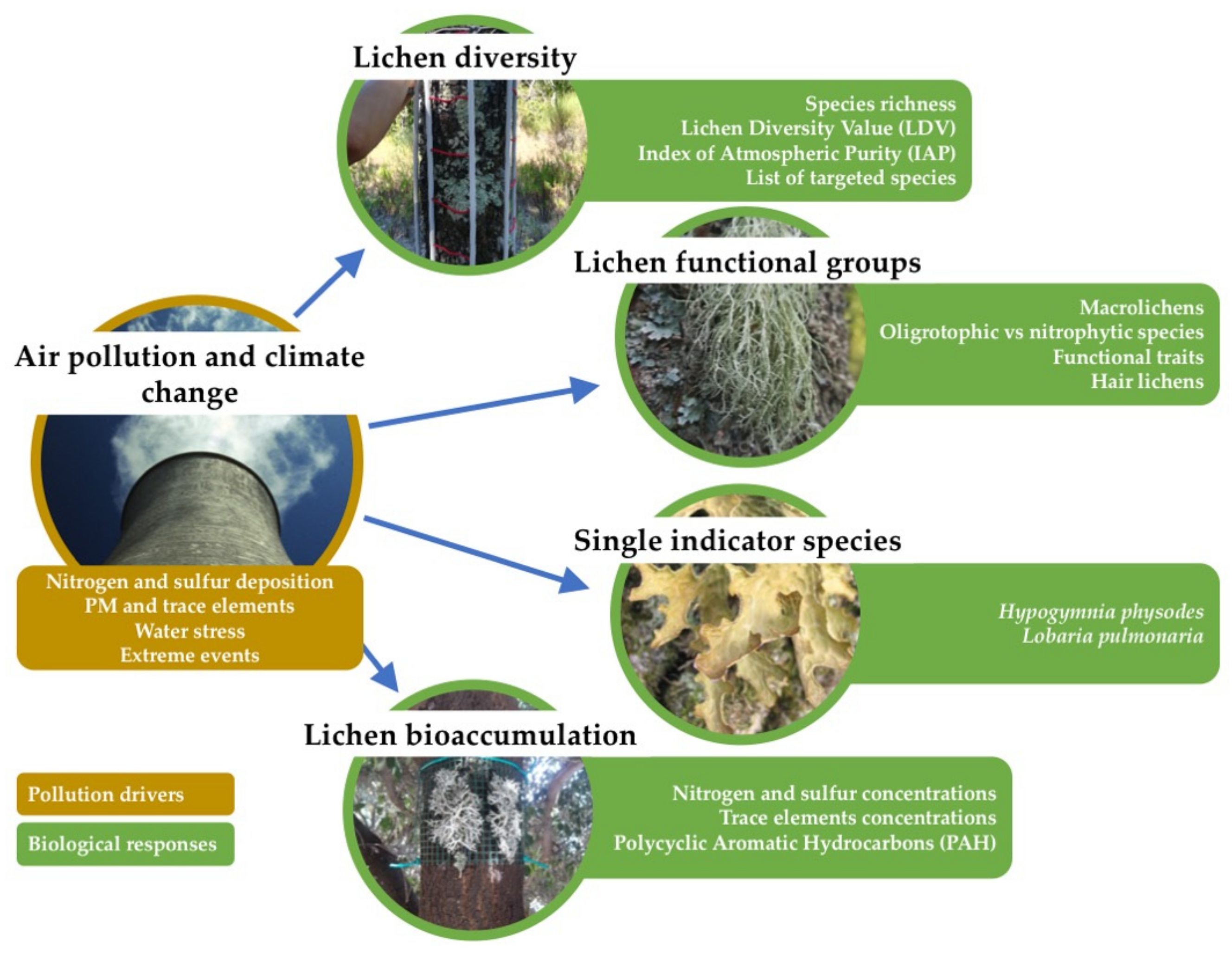
- Several aspects of lichen diversity: the assessment of (i) lichen diversity indices based on lichen presence and abundances, such as the lichen diversity index (LDV) and the index of atmospheric purity (IAP); and (ii) lichen diversity with other methods;
- The responses of lichen functional groups of target species: macrolichens, oligotrophic vs. nitrophytic species, functional traits, hair lichens;
- The viability of single indicator species, such as Hypogymnia physodes (L.) Nyl. and Lobaria pulmonaria (L.) Hoffm.;
- The bioaccumulation of several airborne pollutants, such as nitrogen and sulfur concentrations and other potentially toxic elements (including trace elements and PAHs).
2. Lichen Communities—Lichen Diversity Indices
3. Lichen Functional Groups
4. The Focus on Single Indicator Species
5. Bioaccumulation Approach
6. Conclusions and Perspectives
- With regard to biodiversity methods, well-suited interpretative tools that can better describe air quality and climate changes induced by human activities in forests are still lacking. They could be elaborated based on the previous experience already acquired in the context of studies carried out in urban and industrial areas;
- With regard to functional diversity and traits, research should be encouraged and strengthened, especially delving into the ecophysiological responses of individual lichens or groups of species;
- Using single indicator species could be insufficient as a tool if not supported by biodiversity studies. However, simplified methods based on single forest species, sensitive or resistant to pollution, can be helpful in forest-monitoring programs conducted over large territories and involving many non-expert personnel for the identification of whole lichen communities. Furthermore, since these forest-dwelling species respond not only to air pollution but also to other variables, such as the structure of the forest, it is necessary to improve the interpretation of the results to distinguish the effects of the various environmental drivers;
- Regarding lichen bioaccumulation in forest ecosystems, further efforts to improve the detection of transboundary air pollutants and to promote the adoption of standard methods are needed. Moreover, epiphytic lichens could be effective indicators supporting the use of epigeic mosses, which have been already adopted by the European ICP Forests program to assess heavy metals, nitrogen, microplastics, and persistent organic pollutants (ICP Vegetation Manual [107]). These latter pollutants (POPs) are currently neglected in bioaccumulation studies with lichens in the forest environment. However, this kind of investigation could provide interesting information on the deposition of these pollutants in remote areas.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization. WHO Global Air Quality Guidelines. Particulate Matter (PM2.5 and PM10), Ozone, Nitrogen Dioxide, Sulfur Dioxide and Carbon Monoxide; World Health Organization: Geneva, Switzerland, 2021; Licence: CCBY-NC-SA 3.0IGO. [Google Scholar]
- Waldner, P.; Marchetto, A.; Thimonier, A.; Schmitt, M.; Rogora, M.; Granke, O.; Mues, V.; Hansen, K.; Pihl Karlsson, G.; Žlindra, D.; et al. Detection of temporal trends in atmospheric deposition of inorganic nitrogen and sulphate to forests in Europe. Atmos. Environ. 2014, 95, 363–374. [Google Scholar] [CrossRef] [Green Version]
- Ochoa-Hueso, R.; Munzi, S.; Alonso, R.; Arróniz-Crespo, M.; Avila, A.; Bermejo, V.; Bobbink, R.; Branquinho, C.; Concostrina-Zubiri, L.; Cruz, C.; et al. Ecological impacts of atmospheric pollution and interactions with climate change in terrestrial ecosystems of the Mediterranean Basin: Current research and future directions. Environ. Pollut. 2017, 227, 194–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nash, T.H., III. Lichen Biology, 2nd ed.; Cambridge University Press: Cambridge, UK, 2008. [Google Scholar] [CrossRef]
- Garty, J. Biomonitoring atmospheric heavy metals with lichens: Theory and application. Crit. Rev. Plant Sci. 2001, 20, 309–371. [Google Scholar] [CrossRef]
- Nimis, P.L.; Scheidegger, C.; Wolseley, P.A. Monitoring with lichens-monitoring lichens: An introduction. In Monitoring with Lichens-Monitoring Lichens; Nato science program-IV, Nimis, P.L., Scheidegger, C., Wolseley, P.A., Eds.; Kluwer Academic Publisher: Amsterdam, The Netherlands, 2002; Volume VII, pp. 1–4. [Google Scholar] [CrossRef]
- Giordani, P.; Calatayud, V.; Stofer, S.; Seidling, W.; Granke, O.; Fischer, R. Detecting the nitrogen critical loads on European forests by means of epiphytic lichens. A signal-to-noise evaluation. For. Ecol. Manag. 2014, 311, 29–40. [Google Scholar] [CrossRef]
- Giordani, P.; Brunialti, G. Sampling and interpreting lichen diversity data for biomonitoring purposes. In Recent Advances in Lichenology; Upreti, D.K., Divakar, P.K., Shukla, V., Bajpai, R., Eds.; Springer: Delhi, India, 2015; pp. 19–46. [Google Scholar] [CrossRef]
- Abas, A. A systematic review on biomonitoring using lichen as the biological indicator: A decade of practices, progress and challenges. Ecol. Indic. 2021, 121, 107197. [Google Scholar] [CrossRef]
- Brunialti, G.; Frati, L.; Calderisi, M.; Giorgolo, F.; Bagella, S.; Bertini, G.; Chianucci, F.; Fratini, R.; Gottardini, E.; Cutini, A. Epiphytic lichen diversity and sustainable forest management criteria and indicators: A multivariate and modelling approach in coppice forests of Italy. Ecol. Indic. 2020, 115, 106358. [Google Scholar] [CrossRef]
- Cutini, A.; Ferretti, M.; Bertini, G.; Brunialti, G.; Bagella, S.; Chianucci, F.; Fabbio, G.; Fratini, R.; Riccioli, F.; Caddeo, C.; et al. Testing an expanded set of sustainable forest management indicators in Mediterranean coppice area. Ecol. Indic. 2021, 130, 108040. [Google Scholar] [CrossRef]
- Frati, L.; Brunialti, G.; Landi, S.; Filigheddu, R.; Bagella, S. Exploring the biodiversity of key groups in coppice forests (Central Italy): The relationship among vascular plants, epiphytic lichens, and wood-decaying fungi. Plant Biosyst. 2021, 156, 835–846. [Google Scholar] [CrossRef]
- Oettel, J.; Lapin, K. Linking forest management and biodiversity indicators to strengthen sustainable forest management in Europe. Ecol. Indic. 2021, 122, 107275. [Google Scholar] [CrossRef]
- Ellis, C.J. Lichen epiphyte diversity: A species, community and trait-based review. Perspect. Plant Ecol. Evol. Syst. 2012, 14, 131–152. [Google Scholar] [CrossRef]
- Nascimbene, J.; Benesperi, R.; Giordani, P.; Grube, M.; Marini, L.; Vallese, C.; Mayrhofer, H. Could hair-lichens of high-elevation forests help detect the impact of global change in the Alps? Diversity 2019, 11, 45. [Google Scholar] [CrossRef] [Green Version]
- Ellis, C.J.; Asplund, J.; Benesperi, R.; Branquinho, C.; Di Nuzzo, L.; Hurtado, P.; Martínez, I.; Matos, P.; Nascimbene, J.; Pinho, P.; et al. Functional Traits in Lichen Ecology: A Review of Challenge and Opportunity. Microorganisms 2021, 9, 766. [Google Scholar] [CrossRef] [PubMed]
- Stofer, S.; Calatayud, V.; Giordani, P.; Neville, P. Assessment of epiphytic lichen diversity. In Manual on Methods and Criteria for Harmonized Sampling, Assessment, Monitoring and Analysis of the Effects of Air Pollution on Forests; UNECE ICP Forests Programme Co-ordinating Centre, Ed.; Thünen Institute of Forest Ecosystems: Eberswalde, Germany, 2016; Part VII.2; p. 13 + Annex. Available online: http://www.http://icp-forests.net/page/icp-forests-manual (accessed on 31 January 2023).
- IM Programme Centre. Manual for Integrated Monitoring, August 1998; ICP IM Programme Centre, Finnish Environment Institute: Helsinki, Finland, 1998; p. 248. Available online: www.syke.fi/nature/icpim (accessed on 31 January 2023).
- USFS. Phase 3 Field Guide–Lichen Communities; Version 5.1 October, 2011; United States Forest Service: Washington, DC, USA, 2011. Available online: https://www.fia.fs.usda.gov/library/field-guides-methods-proc/index.php (accessed on 31 January 2023).
- Mayer, A.L.; Vihermaa, L.; Nieminen, N.; Luomi, A.; Posch, M. Epiphytic macrolichen community correlates with modeled air pollutants and forest conditions. Ecol. Indic. 2009, 9, 992–1000. [Google Scholar] [CrossRef]
- LeBlanc, S.C.F.; De Sloover, J. Relation between industrialization and the distribution and growth of epiphytic lichens and mosses in Montreal. Can. J. Bot. 1970, 48, 1485–1496. [Google Scholar] [CrossRef]
- Ammann, K.; Herzig, R.; Liebendoerfer, L.; Urech, M. Multivariate correlation of deposition data of 8 different air pollutants to lichen data in a small town in Switzerland. In Advances in Aerobiology; Boehm, G., Leuschner, R.M., Eds.; Birkhäuser: Basel, Switzerland, 1987; Volume 51, pp. 401–406. [Google Scholar] [CrossRef]
- Kricke, R.; Loppi, S. Bioindication: The I.A.P. approach. In Monitoring with Lichens—Monitoring Lichens; Nimis, P.L., Scheidegger, C., Wolseley, P.A., Eds.; Kluwer: Dordrecht, The Netherlands, 2002; pp. 21–37. [Google Scholar] [CrossRef]
- VDI. Richtlinie 3799, Blatt 1: Ermittlung und Beurteilung Phytotoxischer Wirkungen von Immissionswirkungen mit Flechten: Flechtenkartierung zur Ermittlung des Luftgütewertes (LGW); VDI/DIN-Kommission Reinhaltung der Luft (KRdL)-Normenausschuss: Düsseldorf, Germany, 1995. [Google Scholar]
- Asta, J.; Erhardt, W.; Ferretti, M.; Fornasier, F.; Kirschbaum, U.; Nimis, P.L.; Purvis, O.W.; Pirintsos, S.; Scheidegger, C.; Van Haluwyn, C.; et al. Mapping lichen diversity as an indicator of environmental quality. In Monitoring with Lichens—Monitoring Lichens; Nimis, P.L., Scheidegger, C., Wolseley, P.A., Eds.; Kluwer: Dordrecht, The Netherlands, 2002; pp. 273–279. [Google Scholar] [CrossRef] [Green Version]
- EN 16413. Ambient air-Biomonitoring with Lichens-Assessing Epiphytic Lichen Diversity; CEN-CENELEC Management Center: Brussels, Belgium, 2014. [Google Scholar]
- Giordani, P. Variables influencing the distribution of epiphytic lichens in heterogeneous areas: A case study for Liguria, NW Italy. J. Veg. Sci. 2006, 17, 195–206. [Google Scholar] [CrossRef]
- Giordani, P. Is the diversity of epiphytic lichens a reliable indicator of air pollution? A case study from Italy. Environ. Pollut. 2007, 146, 317–323. [Google Scholar] [CrossRef]
- Svoboda, D. Evaluation of the European method for mapping lichen diversity (LDV) as an indicator of environmental stress in the Czech Republic. Biologia 2007, 62, 424–431. [Google Scholar] [CrossRef]
- Poličnik, H.; Simončič, P.; Batič, F. Monitoring air quality with lichens: A comparison between mapping in forest sites and in open areas. Environ. Pollut. 2008, 151, 395–400. [Google Scholar] [CrossRef]
- Cristofolini, F.; Giordani, P.; Gottardini, E.; Modenesi, P. The response of epiphytic lichens to air pollution and subsets of ecological predictors: A case study from the Italian Prealps. Environ. Pollut. 2008, 151, 308–317. [Google Scholar] [CrossRef]
- Svoboda, D.; Peksa, O.; Veselá, J. Epiphytic lichen diversity in central European oak forests: Assessment of the effects of natural environmental factors and human influences. Environ. Pollut. 2010, 158, 812–819. [Google Scholar] [CrossRef]
- Pinho, P.; Bergamini, A.; Carvalho, P.; Branquinho, C.; Stofer, S.; Scheidegger, C.; Maguas, C. Lichen functional groups as ecological indicators of the effects of land-use in Mediterranean ecosystems. Ecol. Indic. 2012, 15, 36–42. [Google Scholar] [CrossRef]
- Giordani, P.; Brunialti, G.; Bacaro, G.; Nascimbene, J. Functional traits of epiphytic lichens as potential indicators of environmental conditions in forest ecosystems. Ecol. Indic. 2012, 18, 413–420. [Google Scholar] [CrossRef]
- Agnan, Y.; Probst, A.; Séjalon-Delmas, N. Evaluation of lichen species resistance to atmospheric metal pollution by coupling diversity and bioaccumulation approaches: A new bioindication scale for French forested areas. Ecol. Indic. 2017, 72, 99–110. [Google Scholar] [CrossRef] [Green Version]
- Gibson, M.D.; Heal, M.R.; Li, Z.; Kuchta, J.; King, G.H.; Hayes, A.; Lambert, S. The spatial and seasonal variation of nitrogen dioxide and sulfur dioxide in Cape Breton Highlands National Park, Canada, and the association with lichen abundance. Atmos. Environ. 2013, 64, 303–311. [Google Scholar] [CrossRef] [Green Version]
- McMullin, R.T.; Ure, D.; Smith, M.; Clapp, H.; Wiersma, Y.F. Ten years of monitoring air quality and ecological integrity using field-identifiable lichens at Kejimkujik National Park and National Historic Site in Nova Scotia, Canada. Ecol. Indic. 2017, 81, 214–221. [Google Scholar] [CrossRef]
- Correa-Ochoa, M.A.; Vélez-Monsalve, L.C.; Saldarriaga-Molina, J.C.; Jaramillo-Ciro, M.M. Evaluation of the Index of Atmospheric Purity in an American tropical valley through the sampling of corticulous lichens in different phorophyte species. Ecol. Indic. 2020, 115, 106355. [Google Scholar] [CrossRef]
- Conti, M.E.; Cecchetti, G. Biological monitoring: Lichens as bioindicators of air pollution assessment—A review. Environ. Pollut. 2001, 114, 471–492. [Google Scholar] [CrossRef] [PubMed]
- Jovan, S.E.; McCune, B. Air-quality bioindication in the greater Central Valley of California, with epiphytic macrolichen communities. Ecol. Appl. 2005, 15, 1712–1726. [Google Scholar]
- Geiser, L.H.; Neitlich, P.N. Air pollution and climate gradients in western Oregon and Washington indicated by epiphytic macrolichens. Environ. Pollut. 2007, 145, 203–218. [Google Scholar] [CrossRef]
- Blasco, M.; Domeño, C.; López, P.; Nerín, C. Lichens biomonitoring as feasible methodology to assess air pollution in natural ecosystems: Combined study of quantitative PAHs analyses and lichen biodiversity in the Pyrenees Mountains. Anal Bioanal Chem 2008, 391, 759–771. [Google Scholar] [CrossRef]
- Brunialti, G.; Frati, L.; Incerti, G.; Rizzi, G.; Vinci, M.; Giordani, P. Lichen Biomonitoring of air pollution: Issues for applications in complex environments. In Air Quality in the 21st Century; Romano, G.C., Conti, A.G., Eds.; Nova Science Publishers, Inc.: New York, NY, USA, 2009; pp. 211–259. ISBN 9781604567939. [Google Scholar]
- Marmor, L.; Tõrra, T.; Randlane, T. The vertical gradient of bark pH and epiphytic macrolichen biota in relation to alkaline air pollution. Ecol. Indic. 2010, 10, 1137–1143. [Google Scholar] [CrossRef]
- Gadsdon, S.R.; Dagley, J.R.; Wolseley, P.A.; Power, S.A. Relationships between lichen community composition and concentrations of NO2 and NH3. Environ. Pollut. 2010, 158, 2553–2560. [Google Scholar] [CrossRef]
- Pinho, P.; Theobald, M.R.; Dias, T.; Tang, Y.S.; Cruz, C.; Martins-Loução, M.A.; Máguas, C.; Sutton, M.; Branquinho, C. Critical loads of nitrogen deposition and critical levels of atmospheric ammonia for semi-natural Mediterranean evergreen woodlands. Biogeosciences 2012, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Mayer, W.; Pfefferkorn-Dellali, V.; Türk, R.; Dullinger, S.; Mirtl, M.; Dirnböck, T. Significant decrease in epiphytic lichen diversity in a remote area in the European Alps, Austria. Basic Appl. Ecol. 2013, 14, 396–403. [Google Scholar] [CrossRef]
- Geiser, L.H.; Jovan, S.E.; Glavich, D.A.; Fenn, M.E. Predicting lichen-based critical loads for nitrogen deposition in temperate forests. In Nitrogen Deposition, Critical Loads and Biodiversity; Sutton, M., Mason, K., Sheppard, L., Sverdrup, H., Haeuber, R., Hicks, W., Eds.; Springer: Dordrecht, The Netherland, 2014; pp. 311–318. [Google Scholar] [CrossRef]
- McDonough, A.M.; Watmough, S.A. Impacts of nitrogen deposition on herbaceous ground flora and epiphytic foliose lichen species in southern Ontario hardwood forests. Environ. Pollut. 2015, 196, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Degtjarenko, P.; Matos, P.; Marmor, L.; Branquinho, C.; Randlane, T. Functional traits of epiphytic lichens respond to alkaline dust pollution. Fungal Ecol. 2018, 36, 81–88. [Google Scholar] [CrossRef]
- Papitto, G.; Quatrini, V.; Cindolo, C.; Cocciufa, C.; Brunialti, G.; Frati, L. Rilevamento della diversità dei licheni epifiti nell’ambito del monitoraggio in continuo dell’inquinamento atmosferico nei siti della rete CON.ECO.FOR. di Livello II ICP- Forests. Natura 2019, 111, 19. [Google Scholar]
- Geiser, L.H.; Nelson, P.R.; Jovan, S.E.; Root, H.T.; Clark, C.M. Assessing ecological risks from atmospheric deposition of nitrogen and sulfur to US forests using epiphytic macrolichens. Diversity 2019, 11, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanona, M.; Czarnota, P. Index of Atmospheric Purity reflects the ecological conditions better than the environmental pollution in the Carpathian forests. J. Mt. Sci. 2020, 17, 2691–2706. [Google Scholar] [CrossRef]
- Geiser, L.H.; Root, H.T.; Smith, R.J.; Jovan, S.E.; St Clair, L.; Dillman, K.L. Lichen-based critical loads for deposition of nitrogen and sulfur in US forests. Environ. Pollut. 2021, 291, 118187. [Google Scholar] [CrossRef]
- Morillas, L.; Roales, J.; Cruz, C.; Munzi, S. Resilience of epiphytic lichens to combined effects of increasing nitrogen and solar radiation. J. Fungi 2021, 7, 333. [Google Scholar] [CrossRef] [PubMed]
- Stofer, S.; Catalayud, V.; Ferretti, M.; Fischer, R.; Giordani, P.; Keller, C.; Stapper, N.; Scheidegger, C. Epiphytic Lichen Monitoring within the EU/ICP Forests Biodiversity Test-Phase on Level II Plots; Forest Biota project, Technical Report. 2003. Available online: http://icp-forests.net/page/forestbiota (accessed on 3 February 2023).
- Brunialti, G.; Frati, L.; Giordani, P.; Nascimbene, J.; Canullo, R.; Cindolo, C.; Papitto, G. Rete NEC Italia: I risultati della prima campagna di monitoraggio della diversità dei licheni epifiti. Not. Soc. Lich. Ital. 2020, 33, 75–83. [Google Scholar]
- Landgrebe, R.; Best, A.; Hayes, F.; Stein, U.; Harker, G.; Schritt, H.; Duin, L.; Deacon, S. Guidance Note on Site Selection. Support to Member States Regarding the Monitoring of Effects of Air Pollution on Ecosystems according to Article 9(1) of the NEC Directive (Directive (EU) 2016/2284); Technical Report; Ecologic Institute: Berlin, Germany, 2022. [Google Scholar]
- Michel, A.; Kirchner, T.; Prescher, A.-K.; Schwärzel, K. editors Forest Condition in Europe: The 2022 Assessment. ICP Forests Technical Report under the UNECE Convention on Long-range Transboundary Air Pollution (Air Convention); Thünen Institute: Eberswalde, Germany, 2022. [Google Scholar] [CrossRef]
- Petchey, O.L.; Kevin, J.; Gaston, K.J. Functional diversity: Back to basics and looking forward. Ecol. Lett. 2006, 9, 741–758. [Google Scholar] [CrossRef] [PubMed]
- Wolseley, P.A.; Pryor, K.V. The potential of epiphytic twig communities on Quercus petraea in a welsh woodland site (Tycanol) for evaluating environ-mental changes. Lichenologist 1999, 31, 41–61. [Google Scholar] [CrossRef]
- Kinnunen, H.; Holopainen, T.; Kärenlampi, L. Sources of error in epiphytic lichen variables mapped as bioindicators: Needs to modify the Finnish standard. Ecol. Indic. 2003, 3, 1–11. [Google Scholar] [CrossRef]
- Pitcairn, C.E.R.; Leith, I.D.; Sheppard, L.J.; van Dijk, N.; Tang, Y.S.; Wolseley, P.; James, P.; Sutton, M. Field inter-comparison of different bio-indicator methods to assess the impacts of atmospheric nitrogen deposition. In Bioindicator and Biomonitoring Methods for Assessing the Effects of Atmospheric Nitrogen on Statutory Nature Conservation Sites; Sutton, M.A., Pitcairn, C.E.R., Whitfield, C.P., Eds.; JNCC Report 2004; Joint Nature Conservation Committee: Peterborough, UK, 2004; Number 356, Appendix 1; pp. 141–181. [Google Scholar]
- Wolseley, P.A.; Stofer, S.; Mitchell, A.-M.; Vanbergen, A.; Chimonides, J.; Scheidegger, C. Variation of lichen communities with land use in Aberdeenshire, UK. Lichenologist 2006, 38, 307–322. [Google Scholar] [CrossRef] [Green Version]
- Will-Wolf, S.; Geiser, L.H.; Neitlich, P.; Reis, A.H. Forest lichen communities and environment—How consistent are relationships across scales? J. Veg. Sci. 2006, 17, 171–184. [Google Scholar] [CrossRef]
- Wolseley, P.A.; Leith, I.D.; van Dijk, N.; Sutton, M.A. Macrolichens on twigs and trunks as indicators of ammonia concentrations across the UK—A practical method. In Atmospheric Ammonia; Sutton, M.A., Reis, S., Baker, S.M., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 101–108. [Google Scholar] [CrossRef]
- Marini, L.; Nascimbene, J.; Nimis, P.L. Large-scale patterns of epiphytic lichen species richness: Photobiont-dependent response to climate and forest structure. Sci. Total Environ. 2011, 409, 4381–4386. [Google Scholar] [CrossRef]
- Jovan, S.; Riddell, J.; Padgett, P.E.; Nash, T.H. Eutrophic lichens respond to multiple forms of N: Implications for critical levels and critical loads research. Ecol. Appl. 2012, 22, 1910–1922. [Google Scholar] [CrossRef]
- Root, H.T.; Geiser, L.H.; Jovan, S.; Neitlich, P. Epiphytic macrolichen indication of air quality and climate in interior forested mountains of the Pacific Northwest, USA. Ecol. Indic. 2015, 53, 95–105. [Google Scholar] [CrossRef]
- Hurtado, P.; Prieto, M.; De Bello, F.; Aragón, G.; López-Angulo, J.; Giordani, P.; Díaz-Peña, E.M.; Vicente, R.; Merinero, S.; Košuthová, A.; et al. Contrasting environmental drivers determine biodiversity patterns in epiphytic lichen communities along a European gradient. Microorganisms 2020, 8, 1913. [Google Scholar] [CrossRef]
- Hurtado, P.; Prieto, M.; Aragón, G.; De Bello, F.; Martínez, I. Intraspecific variability drives functional changes in lichen epiphytic communities across Europe. Ecology 2020, 101, e03017. [Google Scholar] [CrossRef] [PubMed]
- Łubek, A.; Kukwa, M.; Jaroszewicz, B.; Czortek, P. Shifts in lichen species and functional diversity in a primeval forest ecosystem as a response to environmental changes. Forests 2021, 12, 686. [Google Scholar] [CrossRef]
- SFS 5670; Ilmansuojelu. Bioindikaatio. Jäkäläkartoitus. Finnish Standards Association: Helsinki, Finland, 1990.
- Ellis, C.J. Climate Change, Bioclimatic Models and the Risk to Lichen Diversity. Diversity 2019, 11, 54. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, S.G.; Arup, U.; Baranowski, R.; Ekman, S. Tree-dependent lichens and beetles as indicators in conservation forests. Conserv. Biol. 1995, 9, 1208–1215. [Google Scholar] [CrossRef]
- Campbell, J.; Fredeen, A.L. Lobaria pulmonaria abundance as an indicator of macrolichen diversity in Interior Cedar-Hemlock forests of east–central British Columbia. Canad. J. Bot. 2004, 82, 970–982. [Google Scholar] [CrossRef] [Green Version]
- Jeran, Z.; Mrak, T.; Jaćimović, R.; Batič, F.; Kastelec, D.; Mavsar, R.; Simončič, P. Epiphytic lichens as biomonitors of atmospheric pollution in Slovenian forests. Environ. Pollut. 2007, 146, 324–331. [Google Scholar] [CrossRef]
- Nascimbene, J.; Brunialti, G.; Ravera, S.; Frati, L.; Caniglia, G. Testing Lobaria pulmonaria (L.) Hoffm as an indicator of lichen conservation importance of Italian forests. Ecol. Indic. 2010, 10, 353–360. [Google Scholar] [CrossRef]
- Nascimbene, J.; Benesperi, R.; Brunialti, G.; Catalano, I.; Vedove, M.D.; Grillo, M.; Isocrono, D.; Matteucci, E.; Potenza, G.; Puntillo, D.; et al. Patterns and drivers of beta-diversity and similarity of Lobaria pulmonaria communities in Italian forests. J. Ecol. 2013, 101, 493–505. [Google Scholar] [CrossRef]
- Brunialti, G.; Frati, L.; Ravera, S. Structural variables drive the distribution of the sensitive lichen Lobaria pulmonaria in Mediterranean old-growth forests. Ecol. Indic. 2015, 53, 37–42. [Google Scholar] [CrossRef]
- Brunialti, G.; Frati, L.; Ravera, S. Ecology and conservation of the sensitive lichen Lobaria pulmonaria in Mediterranean old-growth forests. In Old-Growth Forests and Coniferous Forests. Ecology, Habitat and Conservation; Weber, P.R., Ed.; Nova Science Publisher: New York, NY, USA, 2015; pp. 1–20. ISBN 978-1-63482-369-2. [Google Scholar]
- Paoli, L.; Guttová, A.; Sorbo, S.; Lackovičová, A.; Ravera, S.; Landi, S.; Landi, M.; Basile, A.; Sanità di Toppi, L.; Vannini, A.; et al. Does air pollution influence the success of species translocation? Trace elements, ultrastructure and photosynthetic performances in transplants of a threatened forest macrolichen. Ecol. Indic. 2020, 117, 106666. [Google Scholar] [CrossRef]
- Di Nuzzo, L.; Giordani, P.; Benesperi, R.; Brunialti, G.; Fačkovcová, Z.; Frati, L.; Nascimbene, J.; Ravera, S.; Vallese, C.; Paoli, L.; et al. Microclimatic alteration after logging affects the growth of the endangered lichen Lobaria pulmonaria. Plants 2022, 11, 995. [Google Scholar] [CrossRef] [PubMed]
- Wolterbeek, B. Biomonitoring of trace element air pollution: Principles, possibilities and perspectives. Environ. Pollut. 2002, 120, 11–21. [Google Scholar] [CrossRef]
- Augusto, S.; Máguas, C.; Branquinho, C. Guidelines for biomonitoring persistent organic pollutants (POPs), using lichens and aquatic mosses—A review. Environ. Pollut. 2013, 180, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Van der Wat, L.; Forbes, P.B.C. Lichens as biomonitors for organic air pollutants. Trends Analyt. Chem. 2015, 64, 165–172. [Google Scholar] [CrossRef] [Green Version]
- Bargagli, R. Moss and lichen biomonitoring of atmospheric mercury: A review. Sci. Total Environ. 2016, 572, 216–231. [Google Scholar] [CrossRef]
- Loppi, S.; Cenni, E.; Bussotti, F.; Ferretti, M. Biomonitoring of geothermal air pollution by epiphytic lichens and forest trees. Chemosphere 1998, 36, 1079–1082. [Google Scholar] [CrossRef]
- Loppi, S.; Pirintsos, S.A. Epiphytic lichens as sentinels for heavy metal pollution at forest ecosystems (central Italy). Environ. Pollut. 2003, 121, 327–332. [Google Scholar] [CrossRef]
- Blasco, M.; Domeño, C.; Nerín, C. Use of lichens as pollution biomonitors in remote areas: comparison of PAHs extracted from lichens and atmospheric particles sampled in and around the Somport Tunnel (Pyrenees). Environ. Sci. Technol. 2006, 40, 6384–6391. [Google Scholar] [CrossRef]
- Otnyukova, T. Epiphytic lichen growth abnormalities and element concentrations as early indicators of forest decline. Environ. Pollut. 2007, 146, 359–365. [Google Scholar] [CrossRef]
- Conti, M.E.; Pino, A.; Botrè, F.; Bocca, B.; Alimonti, A. Lichen Usnea barbata as biomonitor of airborne elements deposition in the Province of Tierra del Fuego (southern Patagonia, Argentina). Ecotoxicol. Environ. Saf. 2009, 72, 1082–1089. [Google Scholar] [CrossRef]
- Blasco, M.; Domeño, C.; López, P.; Nerín, C. Behaviour of different lichen species as biomonitors of air pollution by PAHs in natural ecosystems. J. Environ. Monit 2011, 13, 2588. [Google Scholar] [CrossRef]
- Root, H.T.; Geiser, L.H.; Fenn, M.E.; Jovan, S.; Hutten, M.A.; Ahuja, S.; Dillman, K.; Schirokauer, D.; Berryman, S.; McMurray, J.A. A simple tool for estimating throughfall nitrogen deposition in forests of western North America using lichens. For. Ecol. Manag. 2013, 306, 1–8. [Google Scholar] [CrossRef]
- Nascimbene, J.; Tretiach, M.; Corana, F.; Lo Schiavo, F.; Kodnik, D.; Dainese, M.; Mannucci, B. Patterns of traffic polycyclic aromatic hydrocarbon pollution in mountain areas can be revealed by lichen biomonitoring: A case study in the Dolomites (Eastern Italian Alps). Sci. Total Environ. 2014, 475, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Root, H.T.; Jovan, S.E.; Fenn, M.; Amacher, M.; Hall, J.; Shaw, J.D. Lichen bioindicators of nitrogen and sulfur deposition in dry forests of Utah and New Mexico, USA. Ecol. Indic. 2021, 127, 107727. [Google Scholar] [CrossRef]
- Agnan, Y.; Séjalon-Delmas, N.; Claustres, A.; Probst, A. Investigation of spatial and temporal metal atmospheric deposition in France through lichen and moss bioaccumulation over one century. Sci. Total Environ. 2015, 529, 285–296. [Google Scholar] [CrossRef] [Green Version]
- Manninen, S. Deriving nitrogen critical levels and loads based on the responses of acidophytic lichen communities on boreal urban Pinus sylvestris trunks. Sci. Total Environ. 2018, 613–614, 751–762. [Google Scholar] [CrossRef] [Green Version]
- Kłos, A.; Ziembik, Z.; Rajfur, M.; Dołhańczuk-Śródka, A.; Bochenek, Z.; Bjerke, J.W.; Tømmervik, H.; Zagajewski, B.; Ziółkowski, D.; Jerz, D.; et al. Using moss and lichens in biomonitoring of heavy-metal contamination of forest areas in southern and north-eastern Poland. Sci. Total Environ. 2018, 627, 438–449. [Google Scholar] [CrossRef] [PubMed]
- Cecconi, E.; Incerti, G.; Capozzi, F.; Adamo, P.; Bargagli, R.; Benesperi, R.; Candotto Carniel, F.; Favero-Longo, S.E.; Giordano, S.; Puntillo, D.; et al. Background element content of the lichen Pseudevernia furfuracea: A supra-national state of art implemented by novel field data from Italy. Sci. Total Environ 2018, 622–623, 282–292. [Google Scholar] [CrossRef] [PubMed]
- Benítez, Á.; Medina, J.; Vásquez, C.; Loaiza, T.; Luzuriaga, Y.; Calva, J. Lichens and bromeliads as bioindicators of heavy metal deposition in Ecuador. Diversity 2019, 11, 28. [Google Scholar] [CrossRef] [Green Version]
- Shukla, V.; Asati, A.; Patel, D.K.; Semwal, M.; Upreti, D.K. Lichens as sinks of airborne organic pollutants: A case study in the natural ecosystem of Himalayas. In Phyto and Rhizo Remediation. Microorganisms for Sustainability; Arora, N., Kumar, N., Eds.; Springer: Singapore, 2019; Volume 9, pp. 175–192. [Google Scholar] [CrossRef]
- Ancora, S.; Dei, R.; Rota, E.; Mariotti, G.; Bianchi, N.; Bargagli, R. Altitudinal variation of trace elements deposition in forest ecosystems along the NW side of Mt. Amiata (central Italy): Evidence from topsoil, mosses and epiphytic lichens. Atmos. Pollut. Res 2021, 12, 101200. [Google Scholar] [CrossRef]
- Carrillo, W.; Calva, J.; Benítez, Á. The Use of bryophytes, lichens and bromeliads for evaluating air and water pollution in an Andean city. Forests 2022, 13, 1607. [Google Scholar] [CrossRef]
- Bargagli, R. Trace Elements in Terrestrial Plants. An Ecophysiological Approach to Biomonitoring and Biorecovery; Springer: Berlin, Germany, 1998; ISBN 9783540645511. [Google Scholar]
- Cecconi, E.; Fortuna, L.; Benesperi, R.; Bianchi, E.; Brunialti, G.; Contardo, T.; Di Nuzzo, L.; Frati, L.; Monaci, F.; Munzi, S.; et al. New interpretative scales for lichen bioaccumulation data: The Italian proposal. Atmosphere 2019, 10, 136. [Google Scholar] [CrossRef] [Green Version]
- UNECE ICP Vegetation. Monitoring of Atmospheric Deposition of Heavy Metals, Nitrogen and POPs in Europe Using Bryophytes. Monitoring Manual 2020 Survey; United Nations Economic Commission for Europe Convention on Long-Range Transboundary Air Pollution: Bangor, UK, 2020; pp. 1–27. [Google Scholar]


{kind=link}
| Index of Atmospheric Purity (IAP) | Lichen Diversity Value (LDV) | Other Lichen Diversity Methods | |
|---|---|---|---|
| Description | The IAP detects lichen diversity within a sampling grid placed on tree trunks. It combines the number of species at the site with their sensitivity towards environmental stressors, primarily air pollution | The LDV is the most recent methodology, and it is strongly standardized to allow easier comparisons throughout Europe; it is not related to any specific pollutant but can be considered an indicator of general environmental quality | In these papers, lichen communities were studied at the tree or plot level without calculating the IAP or LDV |
| Standard methods | LeBlanc and De Sloover [21], Ammann et al. [22], VDI Richtlinie 3799 [24] | Stofer et al. [17], Asta et al. [25], EN 16413 [26] | IM Programme Centre [18], USFS [19] |
| Review papers | Nimis et al. [6], Abas [9], Kricke and Loppi [23], Conti and Cecchetti [39] | Nimis et al. [6], Giordani and Brunialti [8], Kricke and Loppi [23] | Nimis et al. [6], Ellis [14] |
| Year | |||
| 2005 | Jovan and McCune [40] | ||
| 2006 | Giordani [27] | ||
| 2007 | Giordani [28], Svoboda [29] | Geiser and Neitlich [41] | |
| 2008 | Poličnik et al. [30], Blasco et al. [42] | Cristofolini et al. [31] | |
| 2009 | Mayer et al. [20] | Brunialti et al. [43] | |
| 2010 | Svoboda [32] | Marmor et al. [44], Gadsdon et al. [45] | |
| 2012 | Pinho et al. [33], Giordani et al. [34] | ||
| 2013 | Gibson et al. [36] | Pinho et al. [46], Mayer et al. [47] | |
| 2014 | Giordani et al. [7] | Geiser et al. [48] | |
| 2015 | McDomough et al. [49] | ||
| 2017 | Agnan et al. [35], McMullin et al. [37] | Agnan et al. [35] | |
| 2018 | Degtjarenko et al. [50] | ||
| 2019 | Papitto et al. [51] | Geiser et al. [52] | |
| 2020 | Correa-Ochoa et al. [38], Tanona and Czarnota [53] | Geiser et al. [54] | |
| 2021 | Morillas et al. [55] |
| Functional Groups | Oligotrophic vs. Nitrophytic Species | Macrolichens | Other Functional Traits |
|---|---|---|---|
| Description | The proportions of these two functional groups in the lower trunks of forest trees have been confirmed as a suitable indicator of the impact of oxidized and reduced nitrogen compounds. These studies have been used to establish the nitrogen critical loads for epiphytic lichen communities in North America and Europe | Several monitoring protocols consider the responses of macrolichens (foliose and fruticose species) as a suitable tool for assessing the effects of nitrogen and sulfur depositions. Most of these papers also focus on nitrogen-tolerant and -sensitive species | These papers deal with air pollution and climate change and take into account additional functional traits, such as photobiont type, structures for sexual (ascomata type and pigmentation) and asexual reproduction (vegetative propagules, such as pycnidia, sporodochia, isidia, or soredia), and secondary metabolites |
| Review papers | Ochoa-Hueso [3], Giordani and Brunialti [8], Ellis [14], Nascimbene et al. [15], Ellis et al. [16] | ||
| Year | |||
| 1999 | Wolseley et al. [61] | Wolseley et al. [61] | |
| 2003 | Kinnunen et al. [62] | ||
| 2004 | Pitcairn et al. [63] | Pitcairn et al. [63] | |
| 2005 | Jovan and McCune [40] | Jovan and McCune [40] | |
| 2006 | Wolseley et al. [64] | Will-Wolf et al. [65] | |
| 2007 | Geiser and Neitlich [41] | Geiser and Neitlich [41] | |
| 2009 | Wolseley et al. [66] | Wolseley et al. [66] | |
| 2010 | Marmor et al. [44], Gadsdon et al. [45] | Marmor et al. [44] | |
| 2011 | Marini et al. [67] | ||
| 2012 | Pinho et al. [33,46], Giordani et al. [34], Jovan et al. [68] | ||
| 2014 | Giordani et al. [7], Geiser et al. [48] | Geiser et al. [48] | |
| 2015 | McDonough et al. [49] | McDonough et al. [49], Root et al. [69] | |
| 2018 | Degtjarenko et al. [50] | ||
| 2019 | Geiser et al. [52] | Nascimbene et al. [15], Geiser et al. [52] | |
| 2020 | Hurtado et al. [70,71] | ||
| 2021 | Morillas et al. [55] | Geiser et al. [54] | Łubek et al. [72] |
| Indicator Species | Hypogymnia physodes | Lobaria pulmonaria |
|---|---|---|
| Description | This species is considered an indicator species that is rather tolerant to air pollution. Its abundance is determined by counting the number of dots in a sampling grid placed on tree bark. A five-class scale of damage is used to assess the most damaged individuals observed at heights of between 50 and 200 cm above ground | This large foliose species is very sensitive to air pollution and declining heavily throughout Europe. Several studies have demonstrated its suitability both as a flagship and as an umbrella species for nature conservation, and it is associated with many other rare and endangered forest-dwelling organisms |
| Year | ||
| 1995 | Nilsson et al. [75] | |
| 2003 | Kinnunen et al. [62] | |
| 2004 | Campbell and Fredeen [76] | |
| 2006 | Will-Wolf et al. [65] | |
| 2007 | Jeran et al. [77] | |
| 2009 | Mayer et al. [20] | |
| 2010 | Nascimbene et al. [78] | |
| 2013 | Mayer et al. [47] | Nascimbene et al. [79] |
| 2015 | Brunialti et al. [80,81] | |
| 2020 | Paoli et al. [82] | |
| 2022 | Di Nuzzo et al. [73] |
| Nitrogen and/or Sulfur Content | Trace Elements | PAHs | |
|---|---|---|---|
| Description | These papers consider nitrogen and sulfur content in lichen thalli | These papers deal with the bioaccumulation of trace elements, including potentially toxic elements | Epiphytic lichens are also useful biomonitors for persistent organic air pollutants (POPs), such as polycyclic aromatic hydrocarbons (PAHs). These papers represent a selection of these kinds of studies |
| Review papers | Wolterbeek [84], Augusto et al. [85], Van der Wat and Forbes [86], Bargagli [87] | ||
| Year | |||
| 1998 | Loppi et al. [88] | Loppi et al. [88] | |
| 2003 | Loppi and Pirintsos [89] | ||
| 2006 | Blasco et al. [90] | ||
| 2007 | Otnyukova [91] | Jeran et al. [77], Otnyukova [91] | |
| 2008 | Blasco et al. [42] | ||
| 2009 | Conti et al. [92] | Conti et al. [92] | |
| 2011 | Blasco et al. [93] | ||
| 2013 | Root et al. [94] | ||
| 2014 | Nascimbene et al. [95] | ||
| 2015 | Root et al. [96] | Agnan et al. [97] | |
| 2017 | Agnan et al. [35] | ||
| 2018 | Manninen [98] | Kłos et al. [99], Cecconi et al. [100] | |
| 2019 | Benítez et al. [101] | Shukla et al. [102] | |
| 2020 | Paoli et al. [82] | ||
| 2021 | Root et al. [96] | Ancora et al. [103] | |
| 2022 | Carrillo et al. [104] | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frati, L.; Brunialti, G. Recent Trends and Future Challenges for Lichen Biomonitoring in Forests. Forests 2023, 14, 647. https://doi.org/10.3390/f14030647
Frati L, Brunialti G. Recent Trends and Future Challenges for Lichen Biomonitoring in Forests. Forests. 2023; 14(3):647. https://doi.org/10.3390/f14030647
Chicago/Turabian StyleFrati, Luisa, and Giorgio Brunialti. 2023. "Recent Trends and Future Challenges for Lichen Biomonitoring in Forests" Forests 14, no. 3: 647. https://doi.org/10.3390/f14030647
APA StyleFrati, L., & Brunialti, G. (2023). Recent Trends and Future Challenges for Lichen Biomonitoring in Forests. Forests, 14(3), 647. https://doi.org/10.3390/f14030647