
Generalized Linear Models for Describing Tree Gaps in Forest Management Areas in the Brazilian Amazon

 , , and
, , and
Abstract
:1. Introduction
2. Material and Methods
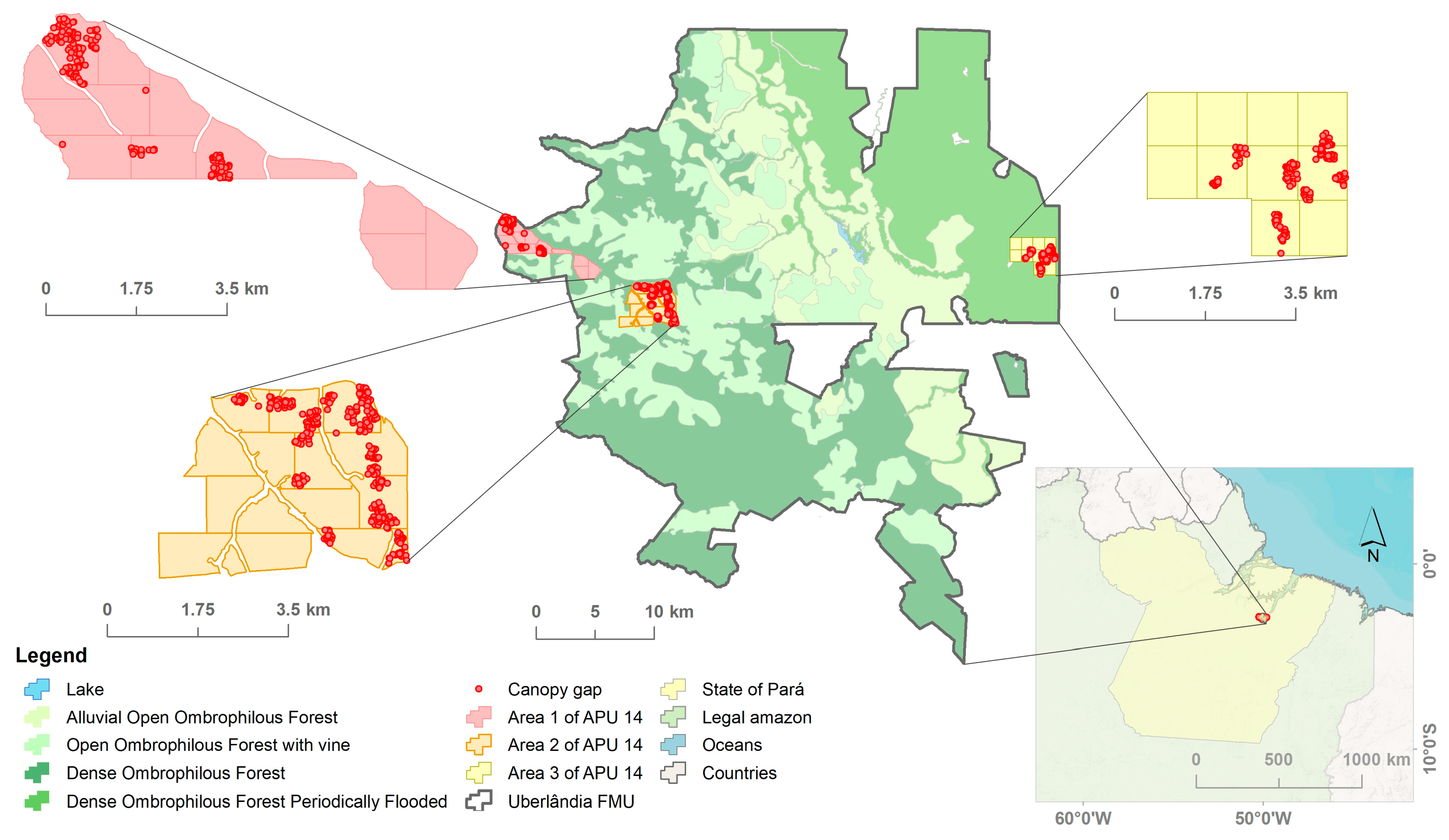
2.1. Characterization of the Study Area
2.2. Dataset
2.2.1. Determining Clearing Areas in the EFEA
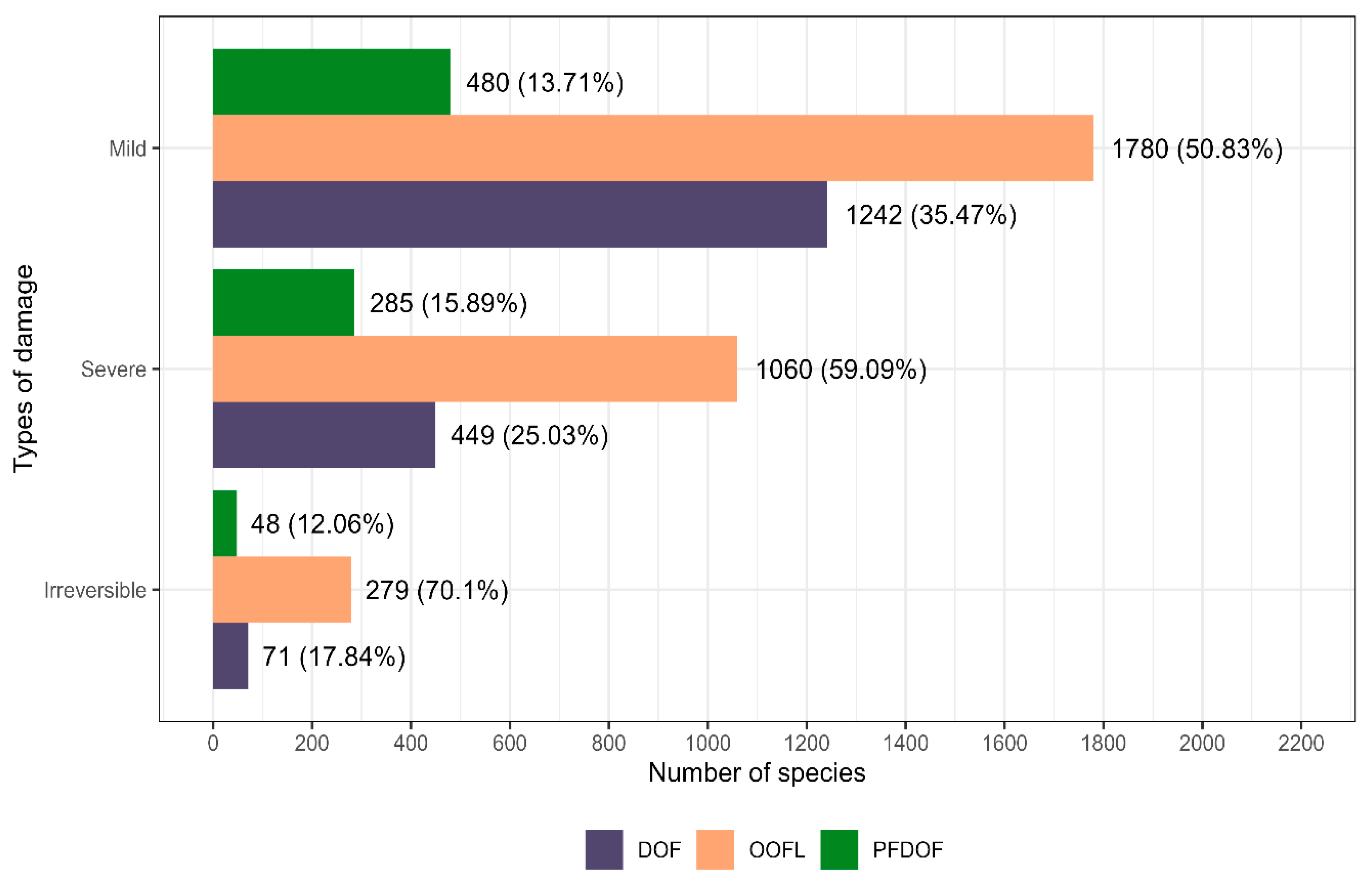
2.2.2. Classification of Damage to Remaining Trees
2.3. Generalized Linear Models
- (a)
- Random component: this consists of the random variable Y from a set of n independent observations (y1, y2, …, yn) with a distribution belonging to the exponential family.
- (b)
- Systematic component: a linear predictor covering a set of covariates through a linear combination of parameters.
- (c)
- Link function: a monotonous and differentiable function that associates the random component (mean of a distribution) with the systematic component (linear predictor). Different link functions can be used for each distribution assumed for the dependent variable.
3. Results
3.1. Species Richness
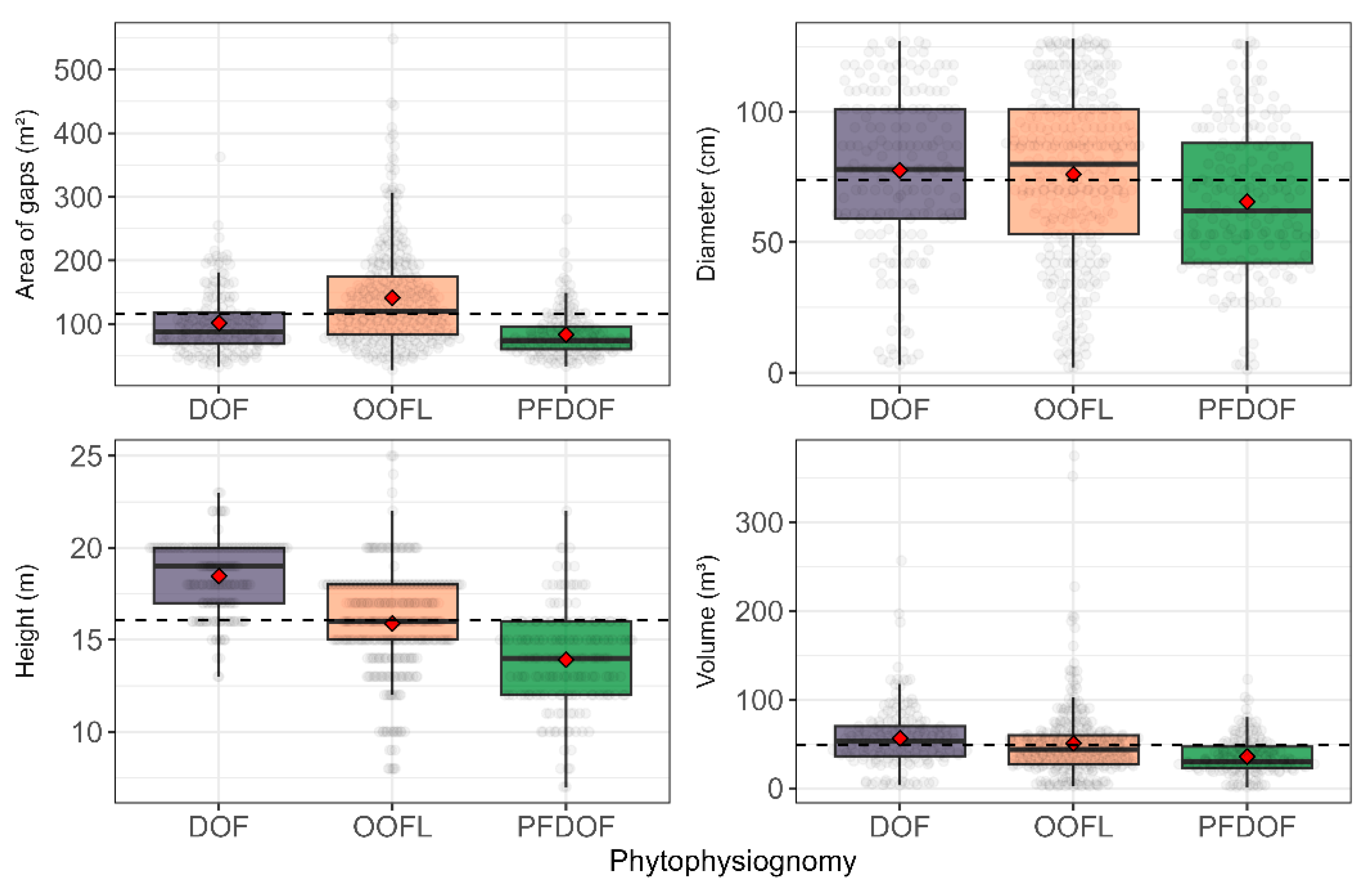
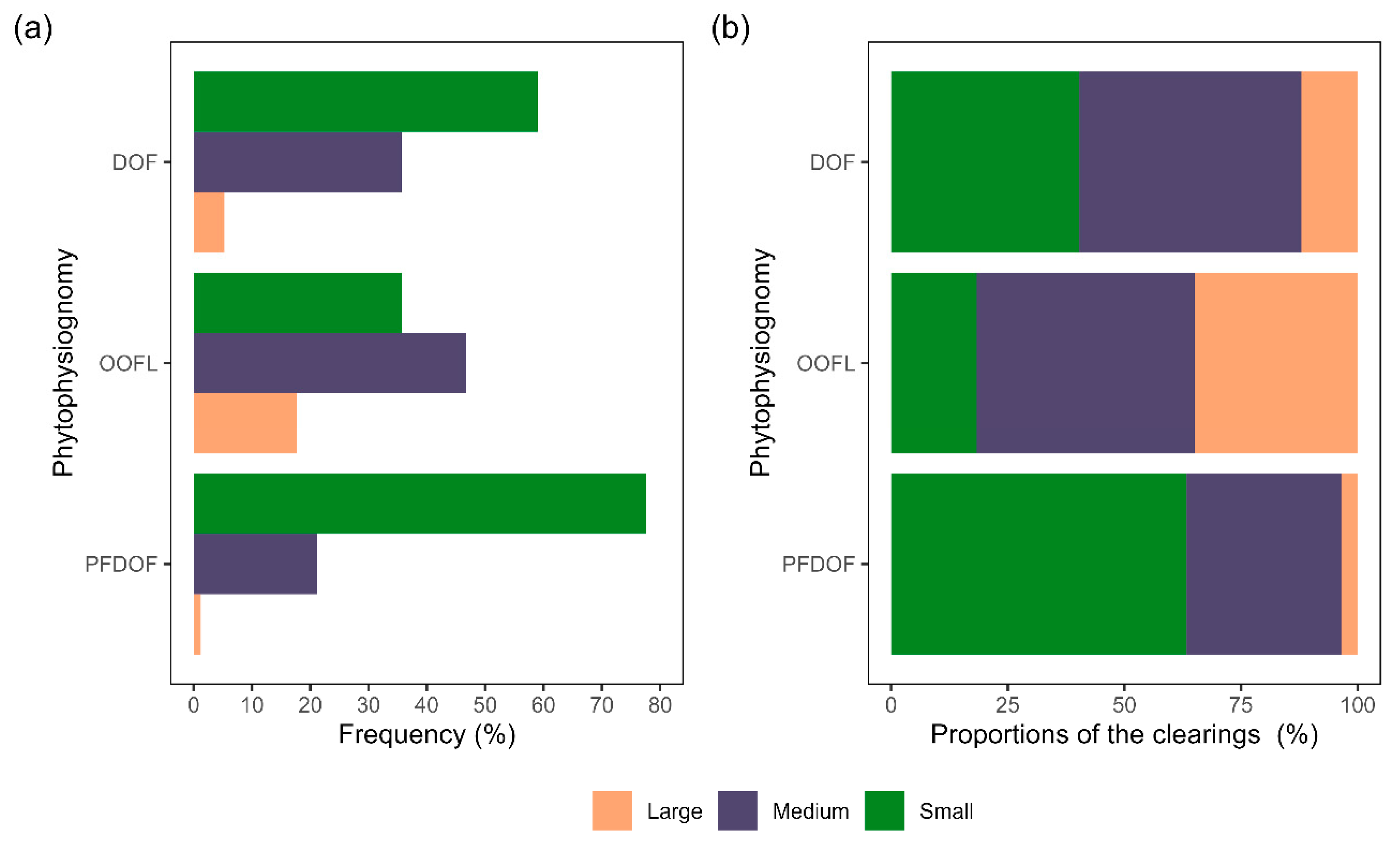
3.2. Dimensioning of the Clearings by Phytophysiognomies
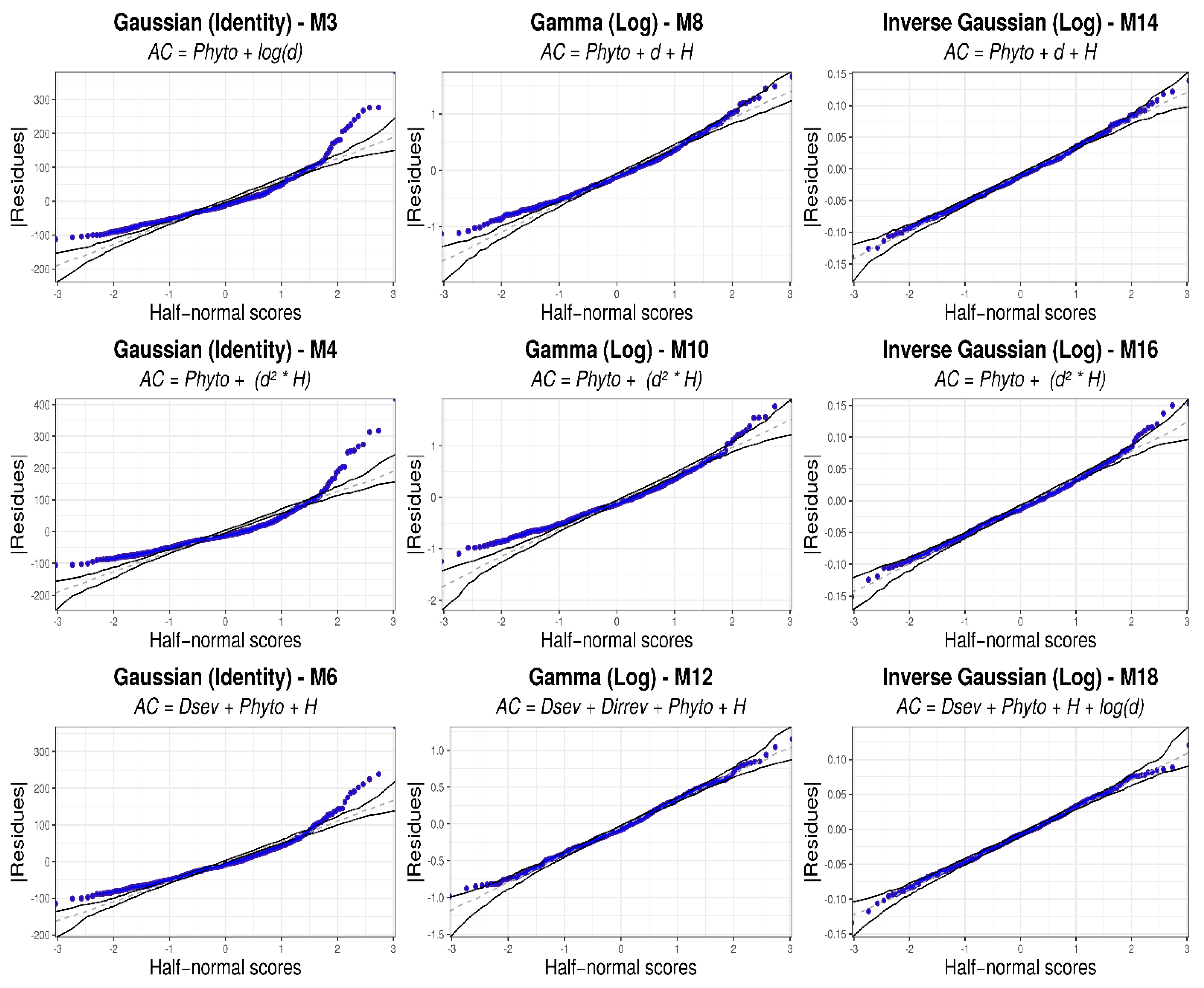
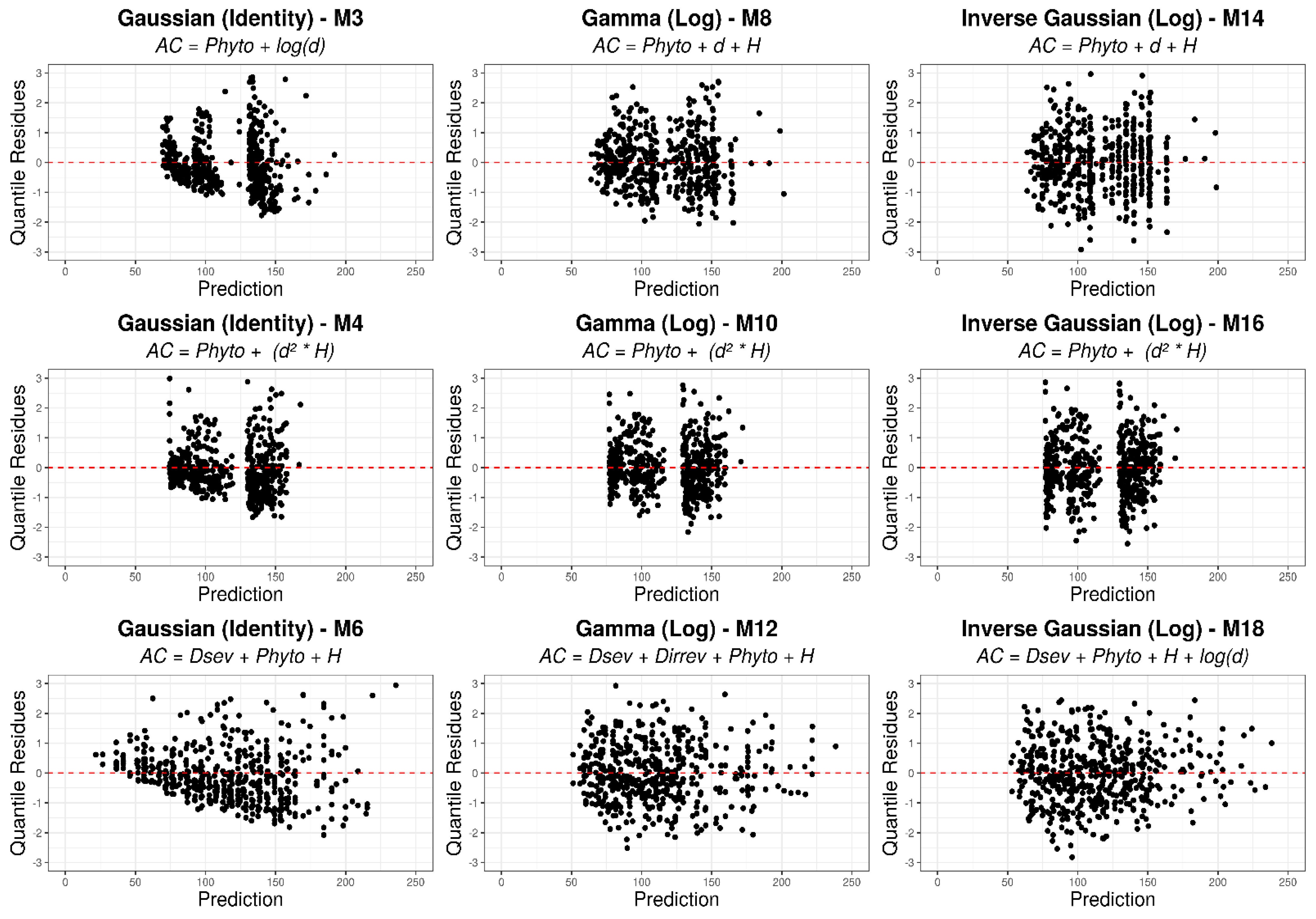
3.3. Generalized Linear Models
4. Discussion
4.1. Species Richness
4.2. Measuring the Clearings by Phytophysiognomy
4.3. Generalized Linear Models
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Locks, C.J.; Matricardi, E.A.T. Estimativa de impactos da extração seletiva de madeiras na Amazônia utilizando dados LIDAR. Ciência Florest. 2019, 29, 481–495. [Google Scholar] [CrossRef]
- Verissimo, A.; Barreto, P.; Mattos, M.; Tarifa, R. Impactos da atividade madeireira e perspectivas para o manejo sustentável da floresta numa velha fronteira da Amazônia: O caso de Paragominas. IMAZON 2002, 1, 26. [Google Scholar]
- de Paula, A.; Silva, A.F.; Marco, P.; Santos, F.A.M.; Souza, A.L. Sucessão ecológica da vegetação arbórea em uma Floresta Estacional Semidecidual, Viçosa, MG, Brasil. Acta Botânica Bras. 2004, 18, 407–423. [Google Scholar] [CrossRef]
- Arruda, E.C.; Cunha, C.N. Características de clareiras e seus efeitos sobre riqueza de espécies em floresta monodominante de Vochysia divergens. Ecol. Aust. 2012, 16, 832–845. [Google Scholar] [CrossRef]
- Frances, M.B.; Carvalho, J.O.P.; Batista, F.J.; Jardim, F.C.S.; Ramos, E.M.L.S. Influência da exploração florestal de impacto reduzido sobre as fases de desenvolvimento de uma floresta de terra firme, Pará, Brasil. Ciências Florestais 2013, 23, 743–753. [Google Scholar] [CrossRef]
- Valtraz, S.; Carvalho, J.O.; Silva, J.N.; Castro, C.C. Efeito da exploração de impacto reduzido na dinâmica do crescimento de uma floresta natural. Sci. For. 2016, 44, 261–271. [Google Scholar]
- Guitet, S.; Pithon, S.; Brumaux, O.; Jubrlin, G.; Gond, V. Impacts of logging on the canopy and the consequences for forest management in French Guiana. For. Ecol. Manag. 2012, 277, 124–131. [Google Scholar] [CrossRef]
- Braz, E.M. Subsídios Para O Planejamento Do Manejo de Florestas Tropicais Da Amazônia; TESE (Doutorado em Engenharia Florestal); Universidade Federal de Santa Maria, Centro de Ciências Rurais: Santa Maria, Brazil, 2010. [Google Scholar]
- LIMA, R.A.F. Estrutura e regeneração de clareiras em Florestas Pluviais Tropicais. Rev. Bras. De Botânica 2005, 28, 651–670. [Google Scholar] [CrossRef]
- D’oliveira, M.V.N.; Reutebuch, S.E.; Mcgaughey, R.J.; Andersen, H.E. Estimating Forest biomass and identifying low-intensity logging areas using airborne scanning LIDAR in Antimary State Forest, Acre State, Western Brazilian Amazon. Remote Sens. Environ. 2012, 124, 479–491. [Google Scholar] [CrossRef]
- Altman, J.; Fibich, P.; Dolezal, J.; Akala, T. TRADER: A package for tree ring analysis of disturbance events in r. Dendrochronolgia 2014, 32, 107–112. [Google Scholar] [CrossRef]
- Mendes, F.d.S.; Jardim, F.C.d.S.; Carvalho, J.O.P.d.; Lima, T.T.S.; Souza, D.V. Dinâmica da composição florística do sub-bosque em floresta tropical manejada, no município de Moju, estado do Pará, Brasil. Rev. De Ciências Agrárias 2012, 55, 117–123. [Google Scholar] [CrossRef]
- Jardim, F.C.S.; Serrão, D.R.; Nemer, T.C. Efeito de diferentes tamanhos de clareiras, sobre o crescimento e a mortalidade de espécies arbóreas, em Moju-PA. Acta Amaz. 2007, 37, 37–48. [Google Scholar] [CrossRef]
- Hunter, M.O.; Keller, M.; Morton, D.; Cook, B.; Lefsky, M.; Ducey, M.; Saleska, S.; de Oliveira, R.C.J.; Schietti, J. Structural dynamics of tropical moist forest gaps. PLoS ONE 2015, 10, e0132144. [Google Scholar] [CrossRef]
- Braz, E.M.; Mattos, P.P.; Verde, M.F.; Basso, R.O.; Canetti, A. Otimização do ciclo de corte na Floresta Amazônica sob o ponto de Vista Econômico; Comunicado Técnico N. 402; Embrapa: Belém, Brasil, 2017; 7p. [Google Scholar]
- Lingner, D.V.; Schorn, L.A.; Sevegnani, L.; de Gasper, A.L.; Meyer, L.; Vibrans, A.C. Floresta Ombrófila Densa de Santa Catarina-Brasil: Agrupamento e ordenação baseados em amostragem sistemática. Ciência Florest. 2015, 25, 933–946. [Google Scholar] [CrossRef]
- Martins, L.; Cavararo, R. Manual Técnico da Vegetação Brasileira: Sistema Fitogeográfico: Inventário das Formações Florestais e Campestres: Técnicas e Manejo de Coleções Botânicas: Procedimentos para Mapeamentos; IBGE: Rio de Janeiro, Brazil, 2012; p. 272.
- Perigolo, N.A. Caracterização dos Tipos Vegetacionais do Médio Rio Madeira, Rondônia; Dissertação (Mestrado em Botânica)—Universidade de Brasília: Brasília, Brazil, 2014. [Google Scholar]
- Souza, P.B.; Neto, J.A.A.M.; Souza, A.L. Diversidade florística e estrutura fitossociológica de um gradiente topográfico em floresta estacional semidecídua sub montana, MG. Cerne 2013, 19, 489–499. [Google Scholar] [CrossRef]
- Estado do Meio Ambiente. Instrução Normativa nº. 5, de 11 de Dezembro de 2006. In Dispõe Sobre Procedimentos Técnicos Para Elaboração, Apresentação, Execução e Avaliação Técnica de Planos de Manejo Florestal Sustentável; IBAMA: Brasília, Brazil, 2006.
- Silva, P.H.; Gomide, L.R.; Figueiredo, E.O.; Carvalho, L.M.T.; Ferraz-filho, A.C. Optimal selective logging regime and log landing location models: A case study in the Amazon forest. Acta Amaz. 2018, 48, 18–27. [Google Scholar] [CrossRef]
- Miller, S.D.; Goulden, M.L.; Hutyra, L.R.; Keller, M.; Saleska, S.R.; Wofsy, S.C.; Figueira, M.S.; da Rocha, H.R.; de Camargo, P.B. Reduced impact logging minimally alters tropical rainforest carbon and energy exchange. Proc. Natl. Acad. Sci. USA 2011, 108, 19431–19435. [Google Scholar] [CrossRef]
- Oliveira, L.C.L.; Jardim, F.C.; Gomes, J.M. Predição do ciclo de corte de espécies arbóreas comerciais por grupos ecológicos em uma floresta na Amazônia brasileira. Rev. Biom 2020, 38, 18–34. [Google Scholar] [CrossRef]
- Instituto de Desenvolvimento Florestal e da Biodiversidade do Estado do Pará—Ideflor-bio. Diretriz de Orientação Para Elaboração do Relatório Anual de Gestão dos Recursos Naturais; Ideflor-Bio: Belém, Brazil, 2019; p. 38.
- Holmes, T.P.; Blate, G.M.; Zweede, J.C.; Pereira, R., Jr.; Barreto, P.; Boltz, F. Custos e Benefícios Financeiros da Exploração de Impacto Reduzido em Comparação à Exploração Florestal Convencional na Amazônia Oriental, 2nd ed.; Fundação Floresta Tropical/Instituto Floresta Tropical (IFT): Belém, Brazil, 2004; p. 68. [Google Scholar]
- Nelder, J.A.; Wedderburn, R.W. Generalized linear models. J. R. Stat. Soc. Ser. A 1972, 135, 370–384. [Google Scholar] [CrossRef]
- Mccullagh, P.; Nelder, J.A. Generalized Linear Models. Monographs on Statistics and Applied Probabibilty; Chapman and Hall: London, UK, 1989. [Google Scholar]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Schwarz, G. Estimating the dimensional of a model. Ann. Stat. 1978, 6, 461–464. [Google Scholar] [CrossRef]
- Sileshi, G.W. A critical review of forest biomass estimation models, common mistakes and corrective measures. For. Ecol. Manag. 2014, 329, 237–254. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. In R Foundation for Statistical Computing; R Core Team: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 31 January 2023).
- Gosiewska, A.; Biecek, P. Auditor: An R Package for Model-Agnostic Visual Validation and Diagnostics. R J. 2019, 11, 85–98. [Google Scholar] [CrossRef]
- Moral, R.A.; Hinde, J.; Demétrio, C.G.B. Half-Normal Plots and Overdispersed Models in R: The hnp Package. J. Stat. Softw. 2017, 81, 1–23. [Google Scholar] [CrossRef]
- Castro, T.d.C.; Carvalho, J.O.P.d.; Schwartz, G.; Silva, J.N.M.; Ruschel, A.R.; de Freitas, L.J.M.; Gomes, J.M.; de Siqueira Pinto, R. The Continuous Timber Production over Cutting Cycles in the Brazilian Amazon Depends on Volumes of Species Not Harvested in Previous Cuts. For. Ecol. Manag. 2021, 490, 119124. [Google Scholar] [CrossRef]
- Scotti, M.S.V.; Biazatti, S.C.; Junior, J.F.B.; Fagundes, S.T.S. Regeneração natural de espécies madeireiras na amazônia ocidental. Rev. Agroecossistemas 2020, 11, 129–145. [Google Scholar] [CrossRef]
- Conceição, A.K.C.; Lira, A.G.S.; Sousa, L.M.R.; Maestri, M.P.; Aquino, M.G.C. Exploração e valoração em tora de 10 espécies florestais no baixo amazonas, estado do Pará, entre 2006–2016. Enciclopédia Biosf. 2020, 17, 80. [Google Scholar] [CrossRef]
- Souza, M.A.S.; Azevedo, C.P.; Souza, C.R.; França, M.; Neto, E.L.V. Dinâmica e produção de uma floresta sob regime de manejo sustentável na Amazônia central. Floresta 2017, 47, 55–63. [Google Scholar] [CrossRef]
- Oliveira, E.K.B.; Nagy, A.C.G.; Barros, Q.S.; Martins, B.C.; Murta, L.S. Composição florística e fitossociológica de fragmento Florestal no sudoeste da Amazônia. Enciclopédia Biosf. 2015, 11, 21–46. [Google Scholar]
- Salomão, R.P.; Santana, A.C.; Brienza, S. Selection of sprecies fron natural and indication of planting for reclamation of degraded areas in the amazon forest. Ciência Florest. 2013, 23, 139–151. [Google Scholar] [CrossRef]
- Instrução normativa no-9, de 8 de maio de 2015; Estabelece os Procedimentos Para Autorizar O Aproveitamento de Matéria-Prima Florestal. IBAMA: Brasília, Brazil, 2015.
- Gerwing, J.; Vidal, E. Manejo de cipós na Amazônia. Ciência Hoje 2005, 37, 220. [Google Scholar]
- Vidal, E.; Viana, V.M.; Batista, J.L.F. Crescimento de floresta tropical três anos após colheita de madeira com e sem manejo florestal na Amazônia oriental. Sci. For. 2002, 61, 133–143. [Google Scholar]
- Popescu, V.D.; Govindarajulu, P.; Palen, W.; Bishop, C.A. Defining conservation-relevant habitat selection by the highly imperiled Oregon Spotted Frog, Rana pretiosa. Herpetol. Conserv. Biol. 2013, 8, 688–706. [Google Scholar]
- Sanquetta, C.R.; Corte, A.P.; Kovalek, N. Estrutura e composição de copas e clareiras em um fragmento de floresta ombrófila mista no centro sul do estado Paraná. Rev. De Estud. Ambient. 2011, 13, 68–77. [Google Scholar]
- Cassol, H.L.G.; Fleig, F.D. Geoprocessing use for the simulation of gaps areas in a management gardening system. Ciência Florest. 2014, 24, 171–183. [Google Scholar] [CrossRef]
- Figueiredo, E.O.; D’oliveira, M.V.N.; Fearnside, P.M.; Braz, E.M.; Papa, D.A. Equations to estimate tree gaps in a precision forest management area the amazon based on crown morphometry. Rev. Árvore 2017, 41, 1–12. [Google Scholar] [CrossRef]
- Dalagnol, R.; Wagner, F.H.; Galvão, L.S.; Streher, A.S.; Phillips, O.L.; Gloor, E.; Pugh, T.A.M.; Ometto, J.P.H.B.; Aragão, L.E.O.C. Large-scale variations in the dynamics of Amazon Forest canopy gaps from airborne lidar data and opportunities for tree mortality estimates. Sci. Rep. 2021, 11, 1388. [Google Scholar] [CrossRef]
- Vidal, E.; Johns, J.; Gerwing, J.; Gerwing, J.; Uhl, C. Manejo de Cipós para a Redução do Impacto da Exploração Madeireira na Amazônia Oriental. IMAZON 1998, 22, 1–24. [Google Scholar]
- Braz, E.M.; Schneider, P.R.; Mattos, P.P.; Selle, G.L.; Thaines, F.; Ribas, L.A.; Vuanden, E. Taxa de corte sustentável para manejo das florestas tropicais. Ciências Florest. 2012, 22, 137–145. [Google Scholar] [CrossRef]
- de Abreu, M.L.; de Azevedo, C.P.; Anholetto, C.R., Jr.; Higashikawa, E.M.; da Silva, K.E. Predição do tamanho de clareira em área de várzea sob regime de manejo florestal: Estudo de caso na reserva de desenvolvimento sustentável mamirauá. Pesqui. Florest. Bras. 2019, 39, 160. [Google Scholar]
- Dearmond, D.; Ferraz, J.B.S.; Marra, D.M.; Amaral, M.R.M.; Lima, A.J.N.; Higuchi, N. Logging Intensity Affects Growth and Lifespan Trajectories for Pioneer Species in Central Amazonia. For. Ecol. Manag. 2022, 522, 120450. [Google Scholar] [CrossRef]
- Cysneiros, V.C.; Junior, J.O.M.; Lanza, T.R.; Moraes, J.C.R.; Samor, O.J.M. Espécies madeireiras da Amazônia: Riqueza, nomes populares e suas peculiaridades. Pesqui. Florest. Bras. 2018, 38, 1–14. [Google Scholar] [CrossRef]
- Lopatin, J.; Dolos, K.; Hernández, H.J.; Galleguilos, M.; Fassnacht, F.E. Comparing Generalized Linear Models and random forest to model vascular plant species richness using LiDAR data in a natural forest in central Chile. Remote Sens. Environ. 2016, 173, 200–210. [Google Scholar] [CrossRef]
- Hess, A.F.; Cianorschi, L.D.; Silvestre, R.; Scariot, R.; Ricken, P. Aplicação dos modelos lineares generalizados para estimativa do crescimento em altura. Pesqui. Florest. Bras. 2015, 35, 427–433. [Google Scholar] [CrossRef]
- Cordeiro, M.G.; Demétrio, C.G.B. Modelos Lineares Generalizados e Extensões. ESALQ 2008, 1, 1–400. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| APU-14 | WT | TA | PPA | EFEA | Phytophysiognomy |
|---|---|---|---|---|---|
| A1 | 11 | 1075.9 | 55.14 | 1000.6 | PFDOF |
| A2 | 14 | 1328.6 | 128.91 | 1194.5 | DOF, OOFL |
| A3 | 14 | 1356.0 | 91.47 | 1264.2 | DOF, OOFL |
| Total | 39 | 3760 | 275.53 | 3460 | - |
| Damage Site | Classification According to Field Parameters | ||
|---|---|---|---|
| Mild | Severe | Irreversible | |
| Damage to treetops | 1/3 canopy of damaged trees | Between 1/3 and 2/3 canopy of damaged trees | Treetops destroyed |
| Damage to tree trunks | Superficial damage in the trunk of the trees | Deep damage of size < ½ d1.3 | Deep damage of size > ½ d1.3 |
| Symbol | Linear Predictor | Family | LF |
|---|---|---|---|
| M1 | β0 + β1d + β2H + β3V | Gaussian | Identity |
| M2 | β0 + β1Phyto + β2d + β3H | Gaussian | Identity |
| M3 | β0 + β1Phyto + β2ln(d) | Gaussian | Identity |
| M4 | β0 + β1Phyto + β2(d2H) | Gaussian | Identity |
| M5 | β0 + β1Phyto + β2ln(d) + β3ln(d2H) | Gaussian | Identity |
| M6 | β0 + β1Dsev + β2Phyto + β3H | Gaussian | Identity |
| M7 | β0 + β1d + β2H + β3V | Gamma | Log |
| M8 | β0 + β1Phyto + β2d + β3H | Gamma | Log |
| M9 | β0 + β1Phyto + β2ln(d) | Gamma | Log |
| M10 | β0 + β1Phyto + β2(d2H) | Gamma | Log |
| M11 | β0 + β1Phyto + β2ln(d) + β3(d2H) | Gamma | Log |
| M12 | β0 + β1Dsev + β2Dirrev + β3Phyto + β4H | Gamma | Log |
| M13 | β0 + β1d + β2H + β3V | Inverse Gaussian | Log |
| M14 | β0 + β1Phyto + β2d + β3H | Inverse Gaussian | Log |
| M15 | β0 + β1Phyto + β2ln(d) | Inverse Gaussian | Log |
| M16 | β0 + β1Phyto + β2(d2H) | Inverse Gaussian | Log |
| M17 | β0 + β1Phyto + β2ln(d) + β3ln(d2H) | Inverse Gaussian | Log |
| M18 | β0 + β1Dsev + β2Phyto + β3H + β4ln(d) + β5d | Inverse Gaussian | Log |
| Species | Family | DOF | PFDOF | OOFL | Total | |
|---|---|---|---|---|---|---|
| Astronium lecointei Ducke | Anacardiaceae | 2 | - | 10 | 12 | |
| Caryocar glabrum (Aubl.) Pers. | Caryocaraceae | - | 9 | 1 | 10 | |
| Caryocar villosum (Aubl.) Pers. | Caryocaraceae | 1 | - | 2 | 3 | |
| Chrysophyllum venezuelanense (Pierre) T.D.Penn. | Sapotaceae | 11 | 2 | 49 | 62 | |
| Cordia goeldiana Huber | Boraginaceae | 2 | - | - | 2 | |
| Couratari guianensis Aubl. | Lecythidaceae | 4 | 32 | 36 | ||
| Couratari stellata A.C.Sm. | Lecythidaceae | 1 | 1 | 3 | 5 | |
| Dinizia excelsa Ducke | Fabaceae | 15 | - | 20 | 35 | |
| Diplotropis martiusii Benth. | Fabaceae | 2 | 24 | - | 26 | |
| Diplotropis purpurea (Rich.) Amshoff | Fabaceae | 2 | 5 | 2 | 9 | |
| Dipteryx polyphylla Huber | Fabaceae | 8 | 13 | 8 | 29 | |
| Endopleura uchi (Huber) Cuatrec. | Humiriaceae | 1 | - | 2 | 3 | |
| Enterolobium schomburgkii (Benth.) Benth. | Fabaceae | 4 | - | 3 | 7 | |
| Erisma uncinatum Warm. | Vochysiaceae | - | 1 | - | 1 | |
| Goupia glabra Aubl. | Celastraceae | 10 | 48 | 22 | 80 | |
| Hymenaea courbaril L. | Fabaceae | 5 | 11 | 3 | 19 | |
| Hymenolobium petraeum Ducke. | Fabaceae | 5 | 11 | 4 | 20 | |
| Iryanthera paraensis Huber | Myristicaceae | - | 5 | 4 | 9 | |
| Lecythis pisonis Cambess. | Lecythidaceae | 1 | - | 1 | 2 | |
| Licaria cannella (Meisn.) Kosterm. | Lauraceae | 4 | - | 3 | 7 | |
| Manilkara huberi (Ducke) Chevalier | Sapotaceae | 21 | - | 38 | 59 | |
| Manilkara paraensis (Huber) Standl. | Sapotaceae | 36 | 4 | 22 | 62 | |
| Micropholis venulosa (Mart. & Eichler) Pierre | Sapotaceae | 7 | - | 12 | 19 | |
| Ocotea neesiana (Miq.) Kosterm. | Lauraceae | - | - | 2 | 2 | |
| Ocotea rubra Mez | Lauraceae | 4 | - | 7 | 11 | |
| Piptadenia suaveolens Miq. | Fabaceae | 3 | - | 15 | 18 | |
| Pouteria oblanceolata Pires | Sapotaceae | 8 | - | 4 | 12 | |
| Qualea paraensis Ducke | Vochysiaceae | - | 6 | - | 6 | |
| Sclerolobium paraense Huber | Fabaceae | 2 | - | - | 2 | |
| Simarouba amara Aubl. | Simaroubaceae | - | 6 | 5 | 11 | |
| Sterculia alata Roxb. | Malvaceae | 3 | 3 | 3 | 9 | |
| Terminalia amazonica (J.F.Gmel) Exell. | Arecaceae | 1 | - | 11 | 12 | |
| Tetragastris panamensis (Engl.) Kuntze | Burseraceae | - | 18 | 18 | ||
| Vantanea parviflora Lam. | Humiriaceae | 6 | - | 3 | 9 | |
| Vatairea paraensis Ducke | Sapotaceae | 2 | 2 | 8 | 12 | |
| Vochysia guianensis Aubl. | Vochysiaceae | 14 | - | 14 | ||
| Total | 36 | 15 | 171 | 165 | 317 | 653 |
| Mod | Coefficients | Adjustment Statistics | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Set Training | Set Test | |||||||||||
| β0 | β1 | β2 | β3 | β4 | β5 | AIC | BIC | rRMSE | AIC | BIC | rRMSE | |
| M1 | −30.0983 | 1.525 * | 1.7572 | 0.5655 | - | - | 5838.92 | 5860.24 | 56.26 | 5881.24 | 5902.55 | 55.24 |
| M2 | −11.7493 | −44.9291 * | 44.1305 * | 1.4245 * | 2.7063 * | - | 5771.37 | 5796.95 | 54.46 | 5617.29 | 5638.6 | 53.25 |
| M3 | −402.316 * | −38.476 * | −47.945 * | 125.77 * | - | - | 5776.53 | 5797.85 | 54.75 | 5565.34 | 5586.65 | 56.14 |
| M4 | 91.56 * | −37.38 * | −49.98 * | 0.0079 * | - | - | 5784.94 | 5806.25 | 55.09 | 5849.14 | 5874.72 | 53.32 |
| M5 | −476.198 * | −44.977 * | −43.689 * | 38.683 | 39.528 * | - | 5771.66 | 5797.24 | 53.52 | 5555.88 | 5581.47 | 54.47 |
| M6 | −5.859 | 20.411 * | −36.245 * | −16.484 * | 4.899 * | - | 5708.05 | 5733.63 | 47.62 | 5708.05 | 5733.63 | 47.56 |
| M7 | 3.1836 * | 0.0169 * | 0.025 * | −0.0263 | - | - | 5565.97 | 5587.29 | 57.25 | 5505.97 | 5531.55 | 57.59 |
| M8 | 3.7453 * | −0.3555 * | −0.4278 * | 0.011 * | 0.0204 * | - | 5467.08 | 5492.66 | 54.36 | 5852.67 | 5873.98 | 53.03 |
| M9 | 0.6113 | −0.3118 * | −0.4542 * | 0.9984 * | - | - | 5466.61 | 5487.93 | 54.62 | 5565.48 | 5586.8 | 55.49 |
| M10 | 4.565 * | −0.3495 * | −0.438 * | 0.0000035 * | - | - | 5478.48 | 5499.79 | 55.03 | 5516.7 | 5538.02 | 53.32 |
| M11 | 0.093 | −0.357 * | −0.4235 * | 0.3136 | 0.3051 * | - | 5461.15 | 5486.73 | 53.22 | 5862.59 | 5883.91 | 53.69 |
| M12 | 3.567 * | 0.1162 * | 0.2111 * | −0.2289 * | −0.095 * | 0.0461 * | 5298.14 | 5327.98 | 41.22 | 5298.14 | 5327.98 | 47.64 |
| M13 | 3.4579 * | 0.0134 * | 0.0154 | 0.0024 | - | - | 5521.60 | 5542.92 | 57.36 | 5569.44 | 5590.76 | 57.47 |
| M14 | 3.7237 * | −0.3407 * | −0.4372 * | 0.0122 * | 0.0166 * | - | 5424.48 | 5450.06 | 54.39 | 5517.69 | 5539.01 | 52.93 |
| M15 | 0.521 | −0.3096 * | −0.4582 * | 0.0198 * | - | - | 5423.73 | 5445.05 | 54.76 | 5830.74 | 5856.32 | 55.31 |
| M16 | 4.48 * | −0.3459 * | −0.4309 * | 0.0000044* | - | - | 5428.13 | 5449.44 | 55.03 | 5537.03 | 5562.61 | 53.38 |
| M17 | 0.1709 | −0.3456 * | −0.4354 * | 0.4295 | 0.2544 * | - | 5420.80 | 5446.38 | 53.41 | 5491.88 | 5517.46 | 53.48 |
| M18 | 3.9067 * | 0.1456 * | −0.2808 * | −0.1968 * | 0.0375 * | −0.0244 | 5371.06 | 5400.91 | 46.01 | 5371.06 | 5400.91 | 47.70 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeferino, S.L.; Gama, N.C.; Souza, D.V.; de Souza, A.S.; Ruz, E.J.H.; da Silva, S.D.B. Generalized Linear Models for Describing Tree Gaps in Forest Management Areas in the Brazilian Amazon. Forests 2023, 14, 841. https://doi.org/10.3390/f14040841
Zeferino SL, Gama NC, Souza DV, de Souza AS, Ruz EJH, da Silva SDB. Generalized Linear Models for Describing Tree Gaps in Forest Management Areas in the Brazilian Amazon. Forests. 2023; 14(4):841. https://doi.org/10.3390/f14040841
Chicago/Turabian StyleZeferino, Suzana Ligoski, Natally Celestino Gama, Deivison Venicio Souza, Alex Soares de Souza, Emil José Hernández Ruz, and Sandra Dezuite Balieiro da Silva. 2023. "Generalized Linear Models for Describing Tree Gaps in Forest Management Areas in the Brazilian Amazon" Forests 14, no. 4: 841. https://doi.org/10.3390/f14040841
APA StyleZeferino, S. L., Gama, N. C., Souza, D. V., de Souza, A. S., Ruz, E. J. H., & da Silva, S. D. B. (2023). Generalized Linear Models for Describing Tree Gaps in Forest Management Areas in the Brazilian Amazon. Forests, 14(4), 841. https://doi.org/10.3390/f14040841