
Age Estimation of Dracaena cinnabari Balf. f. on Socotra Island: A Direct Method to Determine Its Lifespan



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Data Collection
2.3. Age Calculation
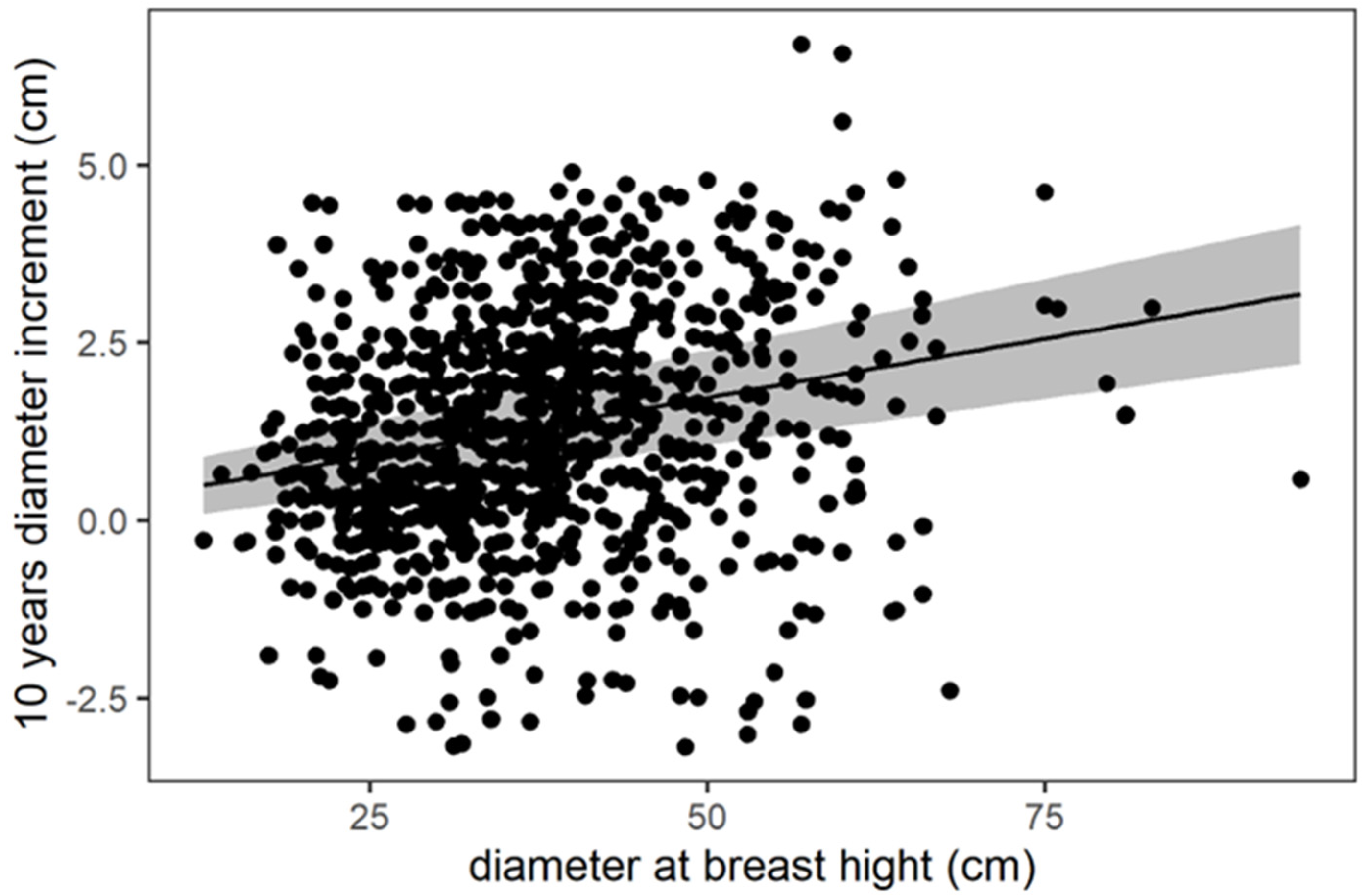
3. Results
3.1. Radial Stem Increment (RSI)
3.2. Dracaena cinnabari Age
4. Discussion
4.1. Radial Stem Increment (RSI)
4.2. Comparisons of the Dragon Tree Age
4.3. Uncertainties in Age Estimation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lu, P.L.; Morden, C.W. Phylogenetic relationships among dracaenoid genera (Asparagaceae: Nolinoideae) inferred from chloroplast DNA loci. Syst. Bot. 2014, 39, 90–104. [Google Scholar] [CrossRef]
- Lengálová, K.; Kalivodová, H.; Habrová, H.; Maděra, P.; Tesfamariam, B.; Šenfeldr, M. First Age-Estimation Model for Dracaena ombet and Dracaena draco subsp. caboverdeana. Forests 2020, 11, 264. [Google Scholar] [CrossRef]
- Govaerts, R.; Zonneveld, B.J.M.; Zona, S.A.; World Checklist of Asparagaceae. Facilitated by the Royal Botanic Gardens, Kew. Available online: http://apps.kew.org/wcsp (accessed on 19 October 2022).
- Vahalík, P.; Patočka, Z.; Drápela, K.; Habrová, H.; Ehrenbergerová, L.; Lengálová, K.; Kalivodová, H.; Pompeiano Vaníčková, L.; Al-Shamahi, E.; Lupton, D.; et al. The conservation status and population mapping of Endangered Dracaena serrulata in the Dhofar Mountains, Oman. Forests 2020, 11, 322. [Google Scholar] [CrossRef]
- Maděra, P.; Forrest, A.; Hanáček, P.; Vahalík, P.; Gebauer, R.; Plichta, R.; Jupa, R.; Van Rensburg, J.J.; Morris, M.; Nadezhdina, N.; et al. What We Know and What We Do Not Know about Dragon Trees? Forests 2020, 11, 236. [Google Scholar] [CrossRef]
- Maděra, P.; Attorre, F.; Habrová, H.; Van Damme, K. Dragon Trees, Tertiary Relicts in Current Reality. Forests 2021, 12, 756. [Google Scholar] [CrossRef]
- Marrero, A.; Almeida, S.R.; Martín-González, M. A new species of the wild Dragon Tree, Dracaena (Dracaenaceae) from Gran Canaria and its taxonomic and biogeographic Implications. Bot. J. Linn. Soc. 1998, 128, 291–314. [Google Scholar]
- Almeida Pérez, R.S. Censo, distribución, habitat y estado de conservation de Dracaena tamaranae A.Marrero, R.S.González-Martín. Gran Canaria, Islas Canarias. Bot. Macarónesica 2003, 24, 39–56. [Google Scholar]
- Zheng, D.J.; Xie, L.S.; Zhu, J.H.; Zhang, Z.L. Low genetic diversity and local adaptive divergence of Dracaena cambodiana (Liliaceae) populations associated with historical population bottlenecks and natural selection: An endangered long-lived tree endemic to Hainan Island, China. Plant Biol. 2012, 14, 828–838. [Google Scholar] [CrossRef]
- Tulik, M.; Wojtan, R.; Jura-Morawiec, J. Theoretical considerations regarding the functional anatomical traits of primary and secondary xylem in dragon tree trunk using the example of Dracaena draco. Planta 2022, 256, 52. [Google Scholar] [CrossRef]
- Habrová, H.; Čermák, Z.; Pavliš, J. Dragon’s blood tree-Threatened by overmaturity, not by extinction: Dynamics of a Dracaena cinnabari woodland in the mountains of Soqotra. Biol. Conserv. 2009, 142, 772–778. [Google Scholar] [CrossRef]
- González, A.G.; León, F.; Hernández, J.C.; Padrón, J.I.; Sánchez-Pinto, L.; Barrera, J.B. Flavans of dragon’s blood from Dracaena draco and Dracaena tamaranae. Biochem. Syst. Ecol. 2004, 32, 179–184. [Google Scholar] [CrossRef]
- Maděra, P.; Habrová, H.; Šenfeldr, M.; Kholová, I.; Lvončík, S.; Ehrenbergerová, L.; Roth, M.; Nadezhdina, N.; Němec, P.; Rosenthal, J.; et al. Growth dynamics of endemic Dracaena cinnabari Balf. f. of Socotra Island suggest essential elements for a conservation strategy. Biológia 2018, 74, 339–349. [Google Scholar] [CrossRef]
- Brown, G.; Mies, B.A. Vegetation Ecology of Socotra; Springer: Dordrecht, The Netherlands, 2012; Volume 7, p. 379. [Google Scholar]
- Hubálková, I.; Maděra, P.; Volařík, D. Growth dynamics of Dracaena cinnabari under controlled conditions as the most effective way to protect endangered species. Saudi. J. Biol. Sci. 2015, 24, 1445–1452. [Google Scholar] [CrossRef]
- Rejžek, M.; Svátek, M.; Šebesta, J.; Adolt, R.; Maděra, P.; Matula, R. Loss of a single tree species will lead to an overall decline in plant diversity: Effect of Dracaena cinnabari Balf. f. on the vegetation of Socotra Island. Biol. Conserv. 2016, 196, 165–172. [Google Scholar] [CrossRef]
- Maděra, P.; Van Damme, K. Socotra Archipelago (Yemen). In Imperiled: The Encyclopedia of Conservation, 1st ed.; DellaSala, D.A., Goldstein, M.I., Eds.; Elsevier: Juneau, AK, USA, 2022; Volume 2, pp. 267–281. [Google Scholar] [CrossRef]
- Culek, M. Geological and morphological evolution of the Socotra archipelago (Yemen) from the biogeographical view. J. Landsc. Ecol. 2013, 6, 84–108. [Google Scholar] [CrossRef]
- Habrová, H.; Vahalík, P.; Drápela, K.; Ali, A.S.S. Distribution and population characteristics of the Cucumber Tree (Dendrosicyos socotranus Balf.f.). Rend. Fis. Acc. Lincei 2020, 31, 725–736. [Google Scholar] [CrossRef]
- Habrová, H.; Pavliš, J. Dynamic response of woody vegetation on fencing protection in semi-arid areas; Case study: Pilot exclosure on the Firmihin Plateau, Socotra Island. Saudi J. Biol. Sci. 2017, 24, 338–346. [Google Scholar] [CrossRef] [PubMed]
- Adolt, R.; Habrová, H.; Maděra, P. Crown age estimation of a monocotyledonous tree species Dracaena cinnabari using logistic regression. Trees Struct. Funct. 2012, 26, 1287–1298. [Google Scholar] [CrossRef]
- Maděra, P.; Volařík, D.; Patočka, Z.; Kalivodová, H.; Divín, J.; Rejžek, M.; Vybíral, J.; Lvončík, S.; Jeník, D.; Hanáček, P.; et al. Sustainable land use management needed to conserve the dragon’s blood tree of Socotra Island, a vulnerable endemic umbrella species. Sustainability 2019, 11, 3557. [Google Scholar] [CrossRef]
- Attorre, F.; Francesconi, F.; Taleb, N.; Scholte, P.; Saed, A.; Alfo, M.; Bruno, F. Will dragonblood survive the next period of climate change? Current and future potential distribution of Dracaena cinnabari (Socotra, Yemen). Biol. Conserv. 2007, 138, 430–439. [Google Scholar] [CrossRef]
- Bauerová, L.; Munie, S.A.; Houšková, K.; Habrová, H. Germination of Dracaena cinnabari Balf.f. Seeds under Controlled Temperature Conditions. Forests 2020, 11, 521. [Google Scholar] [CrossRef]
- García, C.; Vasconcelos, R. The beauty and the beast: Endemic mutualistic interactions promote community-based conservation on Socotra Island (Yemen). J. Nat. Conserv. 2017, 35, 20–23. [Google Scholar] [CrossRef]
- Vasconcelos, R.; Pujol-Buxó, E.; Llorente, G.A.; Saeed, A.; Carranza, S. Micro-Hotspots for Conservation: An Umbrella Tree Species for the Unique Socotran Reptile Fauna. Forests 2020, 11, 353. [Google Scholar] [CrossRef]
- Adolt, R.; Pavliš, J. Age structure and growth of Dracaena cinnabari populations on Socotra. Trees Struct. Funct. 2004, 18, 43–53. [Google Scholar] [CrossRef]
- Jura-Morawiec, J.; Tulik, M.; Iqbal, M. Lateral Meristems Responsible for Secondary Growth of the Monocotyledons: A Survey of the State of the Art. Bot. Rev. 2015, 81, 150–161. [Google Scholar] [CrossRef]
- Worbes, M. One hundred years of tree-ring research in the tropics—A brief history and an outlook to future challenges. Dendrochronologia 2002, 20, 217–231. [Google Scholar] [CrossRef]
- Humboldt, A.v. Relation historique du Voyage aux Régions équinoctiales du Nouveau Continent; F. Schoell: Paris, France, 1814. [Google Scholar]
- Christ, H. Eine Frühlingsfahrt zu den Canarischen Inseln, 1st ed.; H. Georg’s Verlag: Basel, Switzerland, 1886; p. 249. [Google Scholar]
- Schenck, H. Beitrage zur Kenntnis der Vegetation der Canarischen Inseln; Wissensch. Ergebn. d. dtsch. Tiefsee-Expedition “Valdivia”, 2/1; Verlag, G., Ed.; Fischer: Jena, Germany, 1907; pp. 225–406. [Google Scholar]
- Mägdefrau, K. Das Alter der Drachenbäume auf Tenerife. Flora 1975, 164, 347–357. [Google Scholar] [CrossRef]
- Krawczyszyn, J.; Krawczyszyn, T. Photomorphogenesis in Dracaena draco. Trees-Struct. Funct. 2016, 30, 647–664. [Google Scholar] [CrossRef]
- Maděra, P.; Habrová, H.; Čermák, M.; Adolt, R.; Volařík, D. Age estimation of juvenile stages of Dracaena cinnabari Balf. f., the last stone in the mosaic of knowing its lifespan. Rend. Lincei. Sci. Fis. Nat. 2020, 31, 677–686. [Google Scholar] [CrossRef]
- Altman, J.; Doležal, J.; Čížek, L. Age estimation of large trees: New method based on particular increment core tested on an example of veteran oaks. For. Ecol. Manag. 2016, 380, 82–89. [Google Scholar] [CrossRef]
- Al-Okaishi, A. Local Management System of Dragon’s Blood Tree (Dracaena cinnabari Balf. f.) Resin in Firmihin Forest, Socotra Island, Yemen. Forests 2020, 11, 389. [Google Scholar] [CrossRef]
- Culek, M.; Král, K.; Habrová, H.; Adolt, R.; Pavliš, J.; Maděra, P. Socotra´s annual wetaher pattern. In Socotra—A Natural History of the Island and Their People, 1st ed.; Cheung, C., Devantier, L., Eds.; Odyssey Books and Guides; Airphoto International Ltd.: Hong Kong, China, 2006; pp. 62–65. [Google Scholar]
- Habrová, H.; Král, K.; Maděra, P. The weathern pattern in one of the oldest forest ecosystems on Earth—Dragon’s Blood Tree forest (Dracaena cinnabari) on Firmihin—Soqotra island. In Klima lesa, 1st ed.; Rožnovský, J., Litschmann, T., Vyskot, I., Eds.; Křtiny, Czech Bioclimatological Institute: Prague, Czech Republic, 2007; p. 43. [Google Scholar]
- Adolt, R.; Maděra, P.; Abraham, J.; Čupa, P.; Svátek, M.; Matula, R.; Šebesta, J.; Čermák, M.; Volařík, D.; Koutecký, T.; et al. Field survey of Dracaena cinnabari populations in Firmihin, Socotra Island: Methodology and preliminary results. J. Landsc. Ecol. 2013, 6, 7–34. [Google Scholar] [CrossRef]
- Team, R.C. R: A language and environment for statistical computing. R Foundation for Statistical Computing: Vienna, Austria, 2021; p. 2012. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis, 2nd ed.; Springer: New York, NY, USA, 2016; p. 259. Available online: https://ggplot2.tidyverse.org (accessed on 10 January 2023).
- Pütter, A. Altersbestimmung an Drachenbäumen von Tenerife. Sitz. Heidelb. Akad. Wissenschäften. Math.Nat. Kl. 1925, 12, 12–18. [Google Scholar]
- Byström, K. Dracaena draco L. in the Cape Verde Islands. Acta Horti.Gotobg. 1960, 23, 179–214. [Google Scholar]
- Symon, D.E. The growth of Dracaena draco-dragon’s blood tree. J. Arnold Arbor. 1974, 55, 51–58. [Google Scholar] [CrossRef]
- Weiner, J.; Thomas, S.C. The nature of tree growth and the “age related decline in forest productivity. Oikos 2001, 94, 374–376. [Google Scholar] [CrossRef]
- Kalivodová, H.; Culek, M.; Čermák, M.; Maděra, P.; Habrová, H. Potential importance of Socotra dragon´s blood tree cloud forests and woodlands for capturing horizontal precipitation. Rend. Lincei. Sci. Fis. Nat. 2020, 31, 607–621. [Google Scholar] [CrossRef]
- Jura-Morawiec, J.; Tulik, M. Morpho-anatomical basis of dragon´s blood secret in Dracaena draco stem. Flora 2015, 213, 1–5. [Google Scholar] [CrossRef]
- Elzinga, C.; Shearer, R.C.; Elzinga, G. Observer variation in tree diameter measurements. WJAF 2005, 20, 134–137. [Google Scholar] [CrossRef]
- Nadezhdina, N.; Al-Okaishi, A.; Madera, P. Sap flow measurements in a Socotra dragon’s blood tree (Dracaena cinnabari) in its area of origin. Trop. Plant Biol. 2018, 11, 107–118. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Coefficients | Coefficient Estimate | Standard Error | t Value | p Value |
|---|---|---|---|---|
| Intercept | 0.075355 | 0.169486 | 0.445 | 0.657 |
| DBH | 0.033060 | 0.004328 | 7.638 | <0.0001 |
| DBH Class (cm) | Fit Model RSI10 (cm) | RSI10 Lower Confidence Interval | RSI10 Upper Confidence Interval | Age Fit Model | Age Lower Interval | Age Upper Interval |
|---|---|---|---|---|---|---|
| 10.1–15.0 | 0.41 | 0.06 | 0.76 | 111 | 808 | 60 |
| 15.1–20.0 | 0.57 | 0.19 | 0.96 | 198 | 1078 | 112 |
| 20.1–25.0 | 0.74 | 0.32 | 1.16 | 266 | 1237 | 155 |
| 25.1–30.0 | 0.90 | 0.44 | 1.36 | 322 | 1349 | 192 |
| 30.1–35.0 | 1.07 | 0.57 | 1.56 | 369 | 1436 | 224 |
| 35.1–40.0 | 1.23 | 0.70 | 1.76 | 409 | 1507 | 252 |
| 40.1–45.0 | 1.40 | 0.83 | 1.96 | 445 | 1567 | 278 |
| 45.1–50.0 | 1.56 | 0.96 | 2.16 | 477 | 1619 | 301 |
| 50.1–55.0 | 1.73 | 1.09 | 2.36 | 506 | 1665 | 322 |
| 55.1–60.0 | 1.89 | 1.22 | 2.56 | 532 | 1706 | 341 |
| 60.1–65.0 | 2.06 | 1.35 | 2.77 | 557 | 1743 | 359 |
| 65.1–70.0 | 2.22 | 1.48 | 2.97 | 579 | 1777 | 376 |
| 70.1–75.0 | 2.39 | 1.61 | 3.17 | 600 | 1808 | 392 |
| 75.1–80.0 | 2.55 | 1.74 | 3.37 | 619 | 1836 | 407 |
| 80.1–85.0 | 2.72 | 1.87 | 3.57 | 638 | 1863 | 421 |
| 85.1–90.0 | 2.89 | 2.00 | 3.77 | 655 | 1888 | 434 |
| 90.1–95.0 | 3.05 | 2.13 | 3.97 | 672 | 1911 | 447 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bauerová, L.; Maděra, P.; Šenfeldr, M.; Habrová, H. Age Estimation of Dracaena cinnabari Balf. f. on Socotra Island: A Direct Method to Determine Its Lifespan. Forests 2023, 14, 840. https://doi.org/10.3390/f14040840
Bauerová L, Maděra P, Šenfeldr M, Habrová H. Age Estimation of Dracaena cinnabari Balf. f. on Socotra Island: A Direct Method to Determine Its Lifespan. Forests. 2023; 14(4):840. https://doi.org/10.3390/f14040840
Chicago/Turabian StyleBauerová, Lucie, Petr Maděra, Martin Šenfeldr, and Hana Habrová. 2023. "Age Estimation of Dracaena cinnabari Balf. f. on Socotra Island: A Direct Method to Determine Its Lifespan" Forests 14, no. 4: 840. https://doi.org/10.3390/f14040840
APA StyleBauerová, L., Maděra, P., Šenfeldr, M., & Habrová, H. (2023). Age Estimation of Dracaena cinnabari Balf. f. on Socotra Island: A Direct Method to Determine Its Lifespan. Forests, 14(4), 840. https://doi.org/10.3390/f14040840