
UAV Inventory of the Last Remaining Dragon Tree Forest on Earth

 ,
,  , , , , , , and
, , , , , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
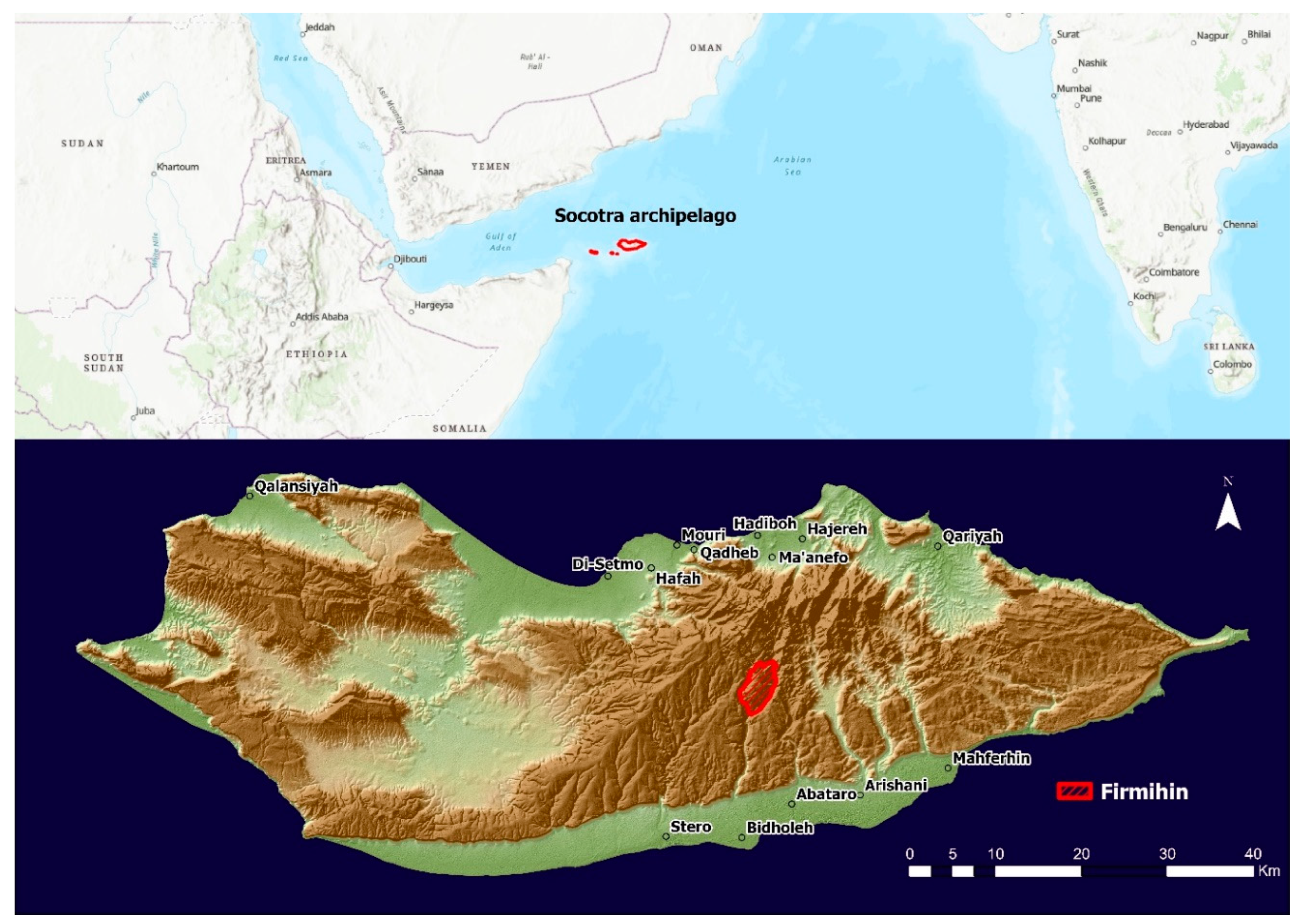
2.1. Study Area
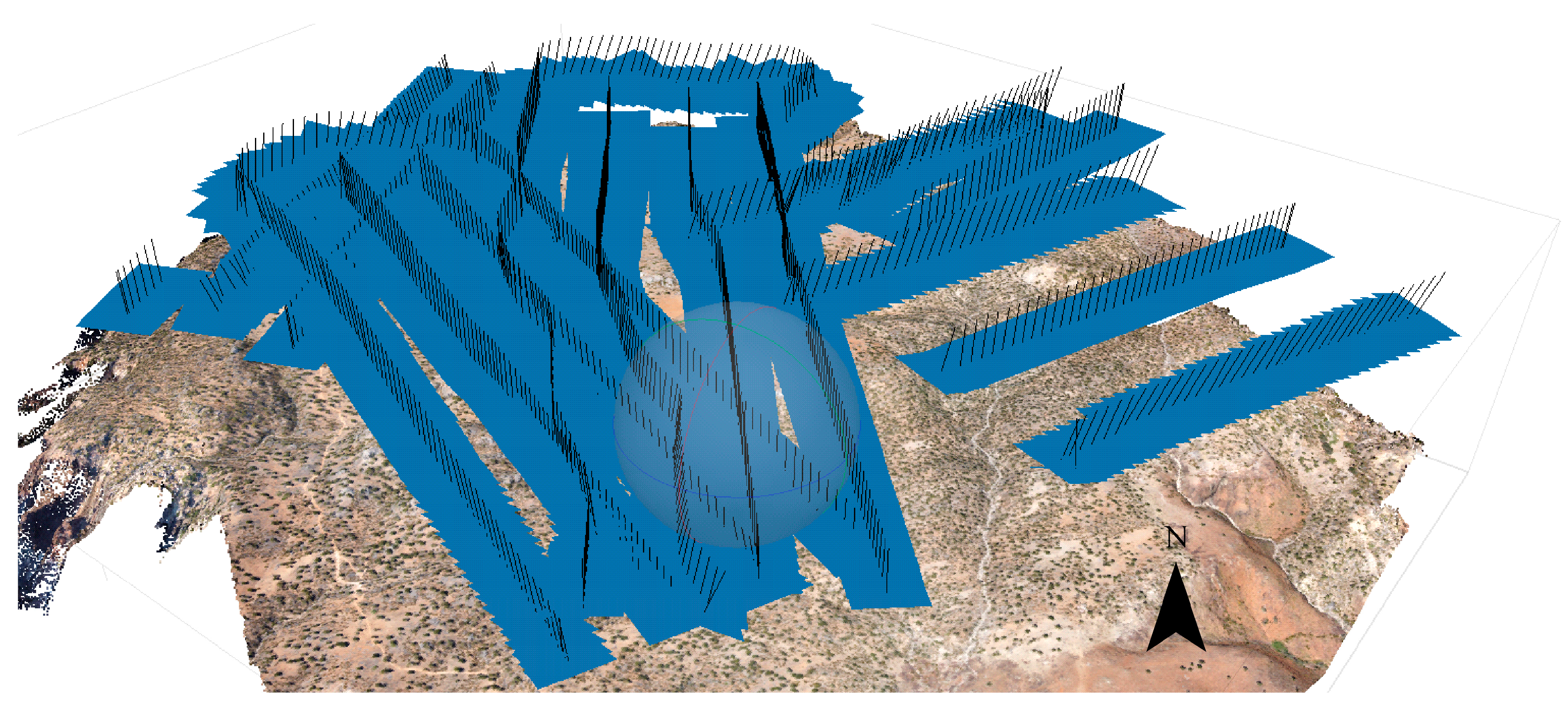
2.2. Field Survey and Aerial Flights
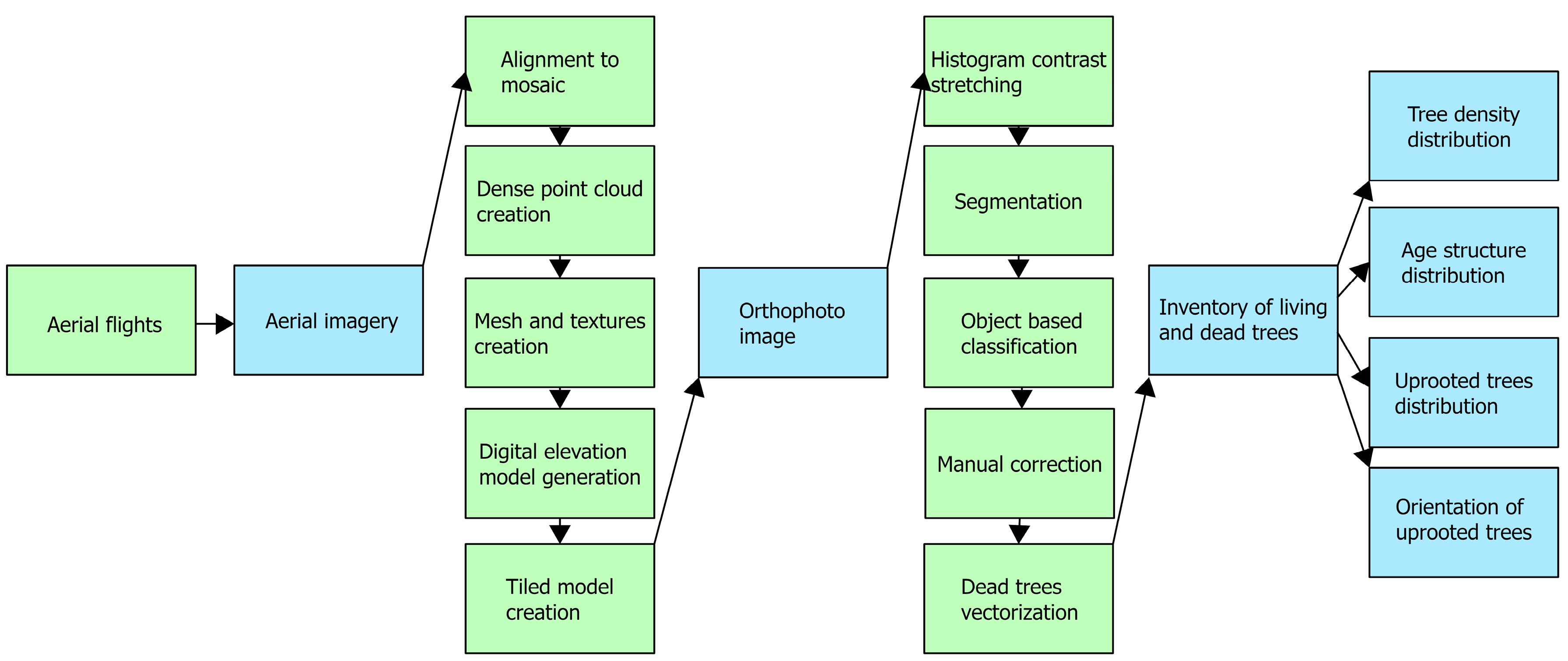
2.3. Data Processing
2.3.1. Processing of Aerial Images
2.3.2. Tree Inventory
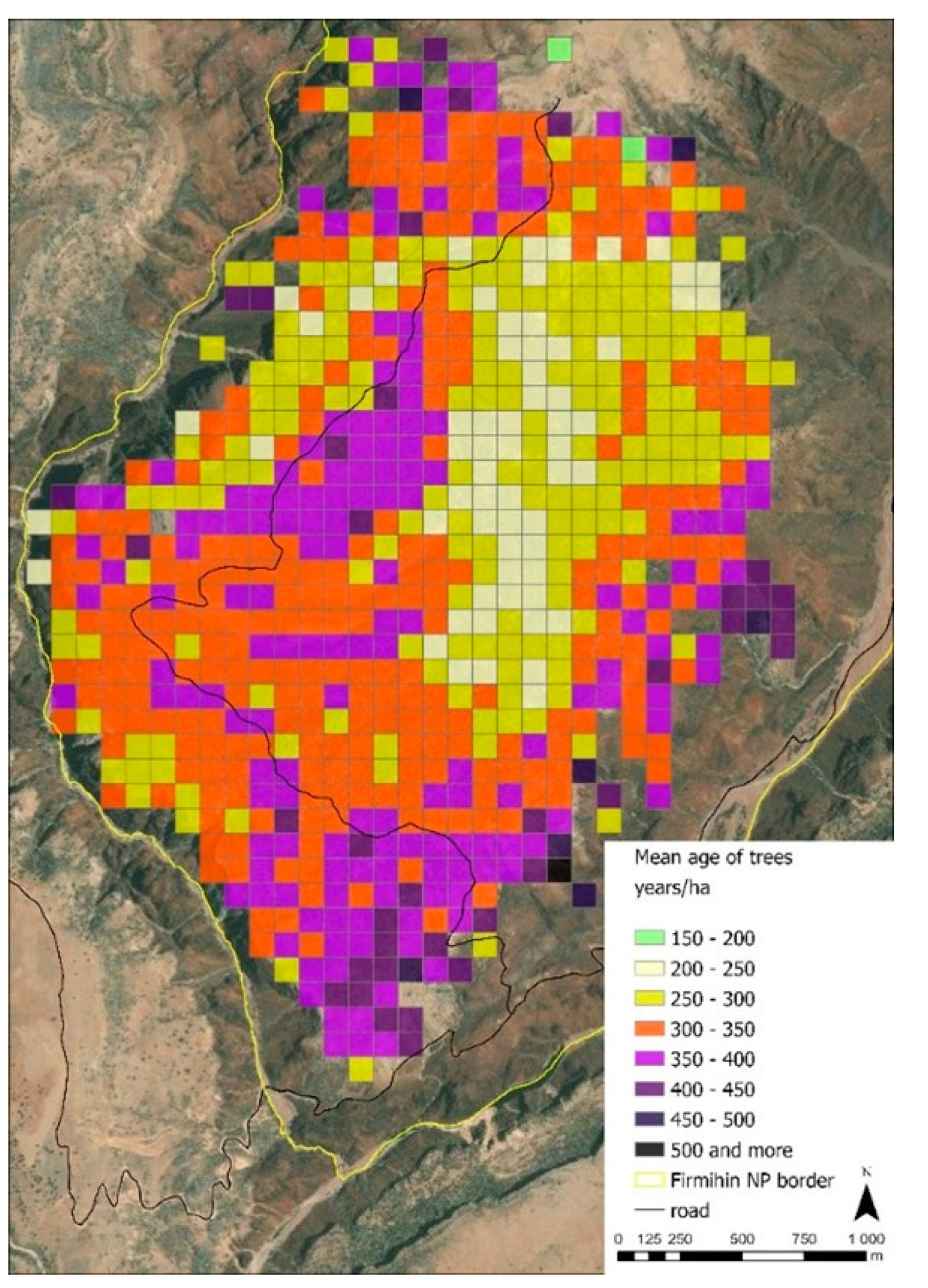
2.3.3. Spatial Age Distribution Analysis
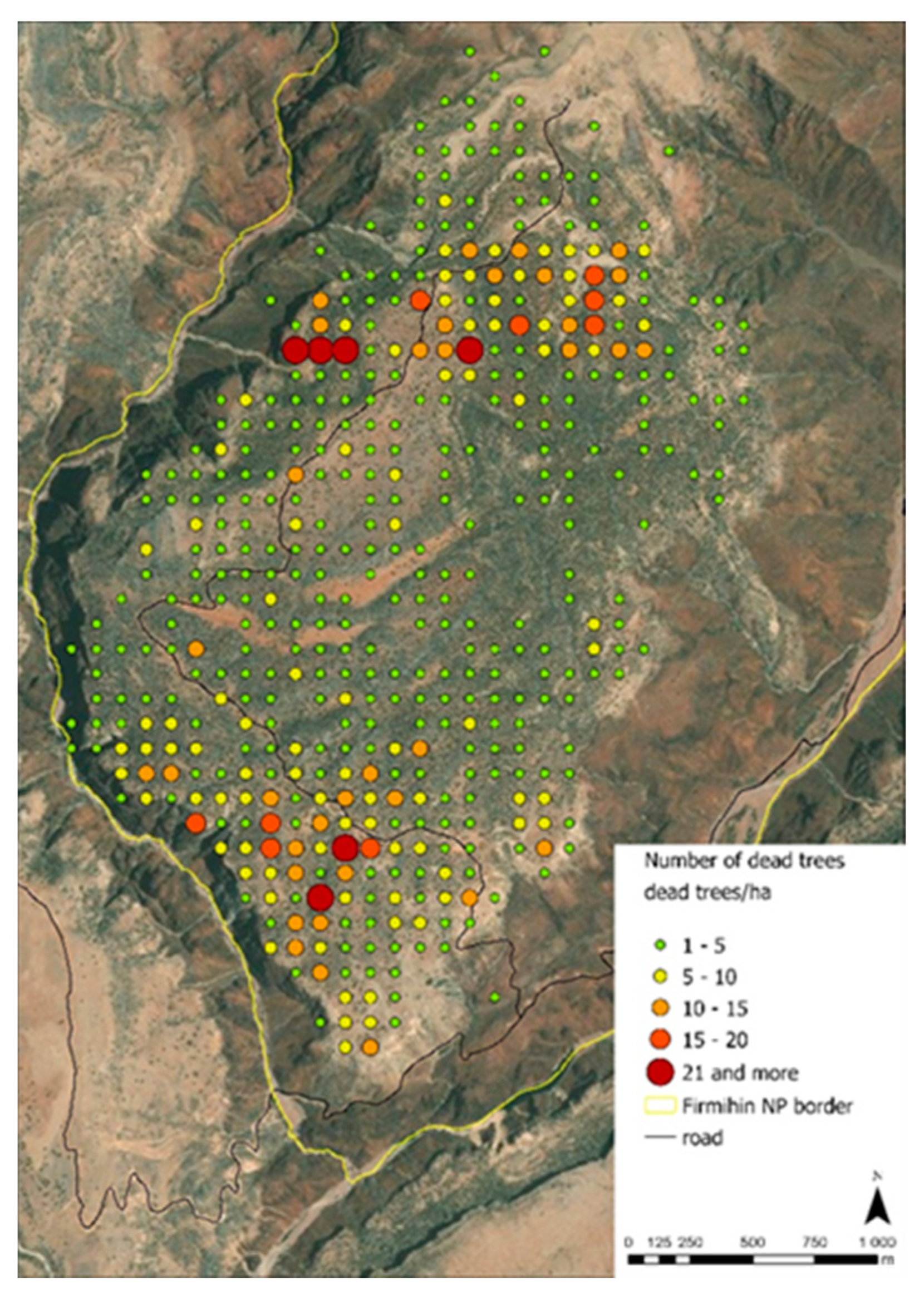
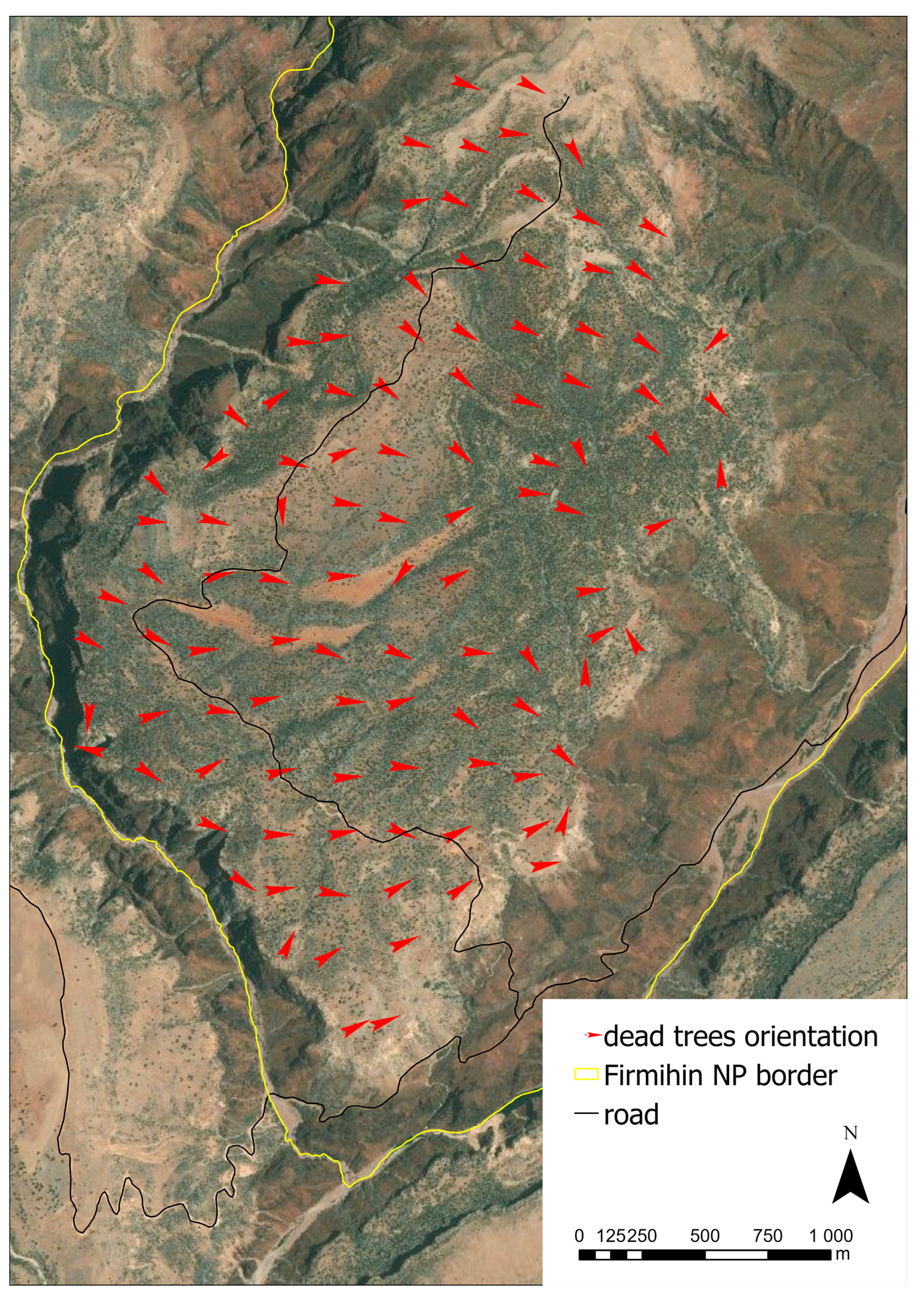
2.3.4. Inventory of Uprooted Trees
3. Results
3.1. Inventory of the Firmihin Forest
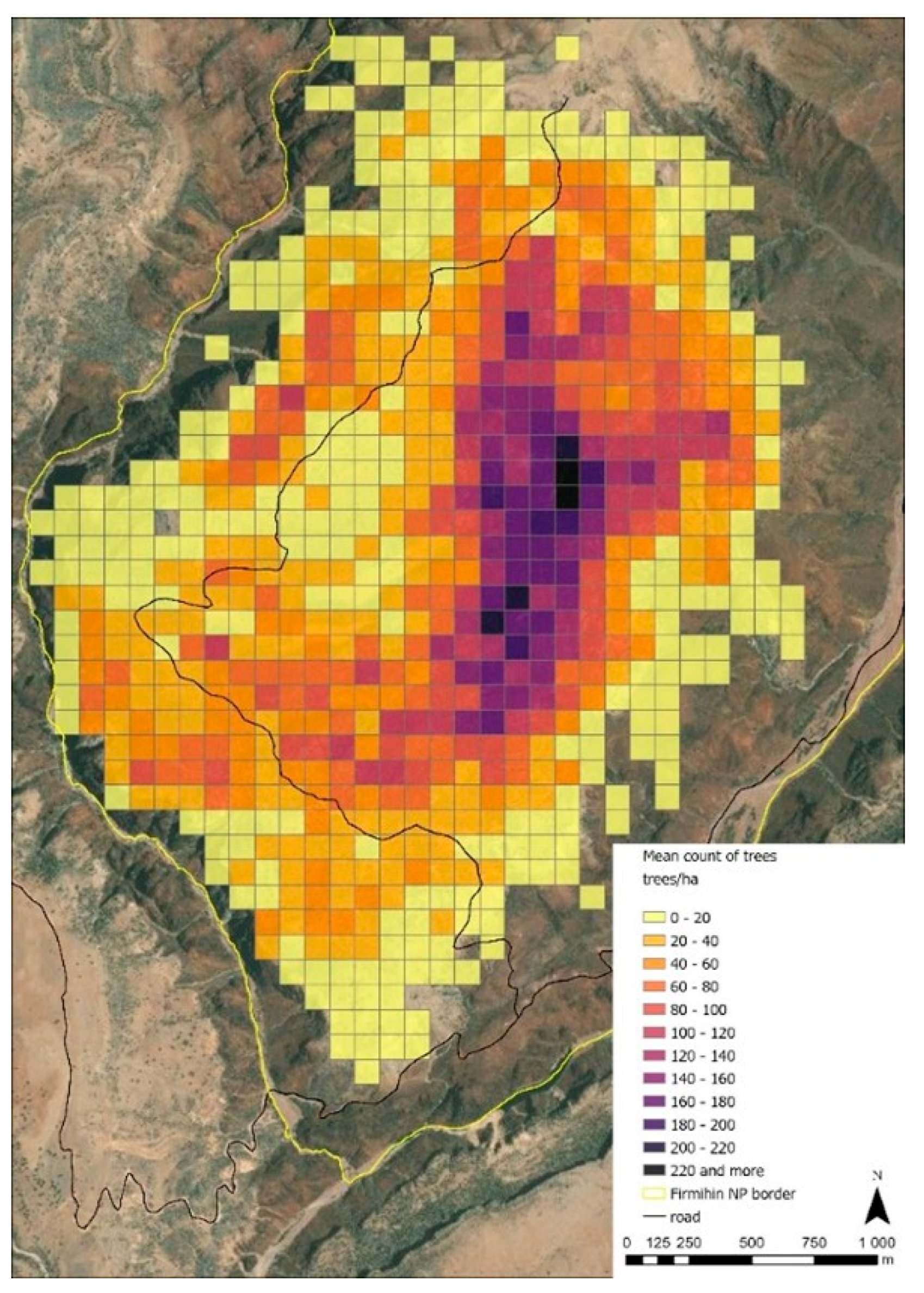
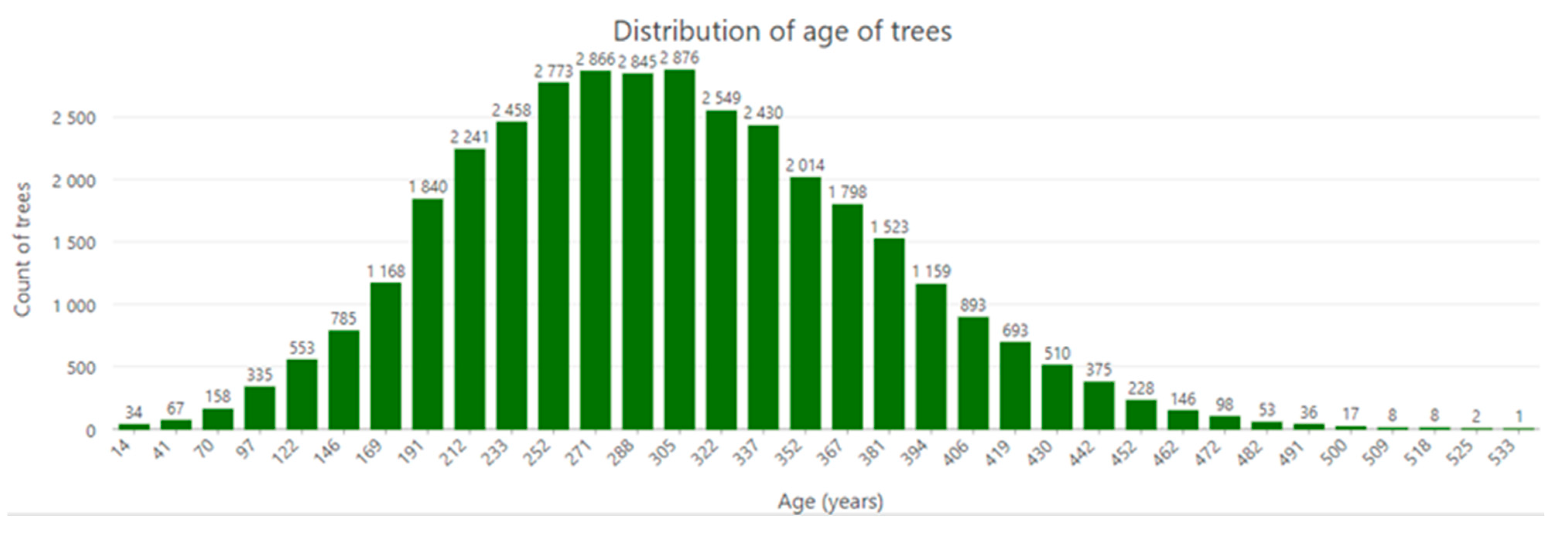
3.2. Tree Density and Age Structure
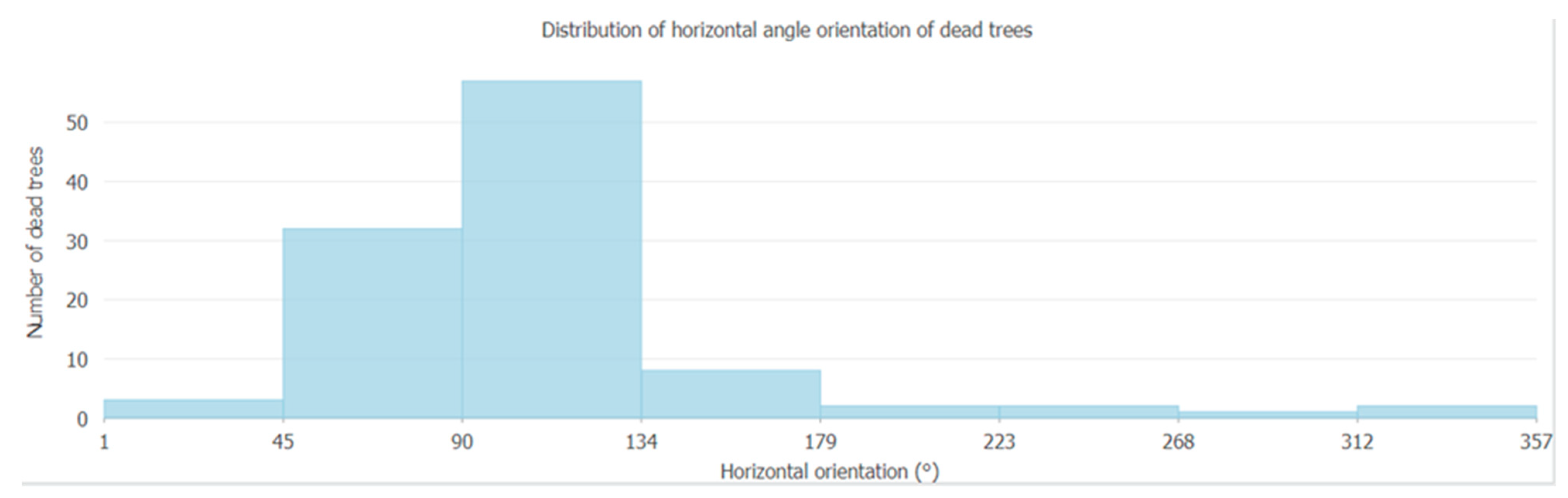
3.3. Evaluation of Uprooted Trees
4. Discussion
4.1. Previous Inventories
4.2. Population Structure and Natural Regeneration
4.3. Evaluation of Dead Trees and the Link to Global Warming
4.4. Significance of Dracaena Conservation
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maděra, P.; Forrest, A.; Hanáček, P.; Vahalík, P.; Gebauer, R.; Plichta, R.; Jupa, R.; Van Rensburg, J.J.; Morris, M.; Nadezhdina, N.; et al. What We Know and What We Do Not Know about Dragon Trees? Forests 2020, 11, 236. [Google Scholar] [CrossRef] [Green Version]
- Denk, T.; Güner, H.T.; Grimm, G.W. From mesic to arid: Leaf epidermal features suggest preadaptation in Miocene dragon trees (Dracaena). Rev. Palaeobot. Palynol. 2014, 200, 211–228. [Google Scholar] [CrossRef]
- Balfour, I.B. Botany of Socotra. Natural History Museum Library: London, UK, 1888. [Google Scholar] [CrossRef] [Green Version]
- Casson, L. The Periplus Maris Erythraei: Text with Introduction, Translation, and Commentary; Princeton University Press: Princeton, NJ, USA, 1989. [Google Scholar]
- Cheung, C.P.S.; DeVantier, L.M. Socotra—A Natural History of the Islands and their People. In Odyssey Books and Guides; Van Damme, K., Ed.; Airphoto International Ltd.: Hong Kong, China, 2006; 408p. [Google Scholar]
- Miller, A.G.; Morris, M.; Diccon, A.; Atkinson, R. Ethnoflora of the Soqotra Archipelago, 1st ed.; Royal Botanic Garden: Edinburgh, UK, 2004; p. 759. [Google Scholar]
- Al-Okaishi, A. Local Management System of Dragon’s Blood Tree (Dracaena cinnabari Balf. F.) Resin in Firmihin Forest, Socotra Island, Yemen. Forests 2020, 11, 389. [Google Scholar] [CrossRef] [Green Version]
- Bent, T. Southern Arabia; Smith, Elder and Co.: London, UK, 1900; pp. 351–352. [Google Scholar]
- Attorre, F.; Francesconi, F.; Taleb, N.; Scholte, P.; Saed, A.; Alfo, M.; Bruno, F. Will dragonblood survive the next period of climate change? Current and future potential distribution of Dracaena cinnabari (Socotra, Yemen). Biol. Conserv. 2007, 138, 430–439. [Google Scholar] [CrossRef]
- Rezende, M.; Maděra, P.; Vahalík, P.; Van Damme, K.; Habrová, H.; Riccardi, T.; Attorre, F.; De Sanctis, M.; Sallemi, G.; Malatesta, L. Identifying Suitable Restoration and Conservation Areas for Dracaena cinnabari Balf. f. In Socotra, Yemen. Forests 2022, 13, 1276. [Google Scholar] [CrossRef]
- Maděra, P.; Volařík, D.; Patočka, Z.; Kalivodová, H.; Divín, J.; Rejžek, M.; Vybíral, J.; Lvončík, S.; Jeník, D.; Hanáček, P.; et al. Sustainable land use management needed to conserve the dragon´s blood tree of Socotra Island, a vulnerable endemic umbrella species. Sustainability 2019, 11, 3557. [Google Scholar] [CrossRef] [Green Version]
- Habrová, H.; Buček, A. Overview of Biotope Types of Socotra Island. J. Landsc. Ecol. 2014, 6, 60–83. [Google Scholar] [CrossRef]
- Král, K.; Pavliš, J. The First Detailed Land-Cover Map of Socotra Island by Landsat/ETM+ Data. Int. J. Remote Sens. 2006, 27, 3239–3250. [Google Scholar] [CrossRef]
- Adolt, R.; Pavlis, J. Age structure and growth of Dracaena cinnabari populations on Socotra. Trees-Struct. Funct. 2004, 18, 43–53. [Google Scholar] [CrossRef]
- Adolt, R.; Habrova, H.; Maděra, P. Crown age estimation of a monocotyledonous tree species Dracaena cinnabari using logistic regression. Trees 2012, 26, 1287–1298. [Google Scholar] [CrossRef]
- Habrova, H.; Pavlis, J. Dynamic response of woody vegetation on fencing protection in semi-arid areas; Case study: Pilot exclosure on the Firmihin Plateau, Socotra Island. Saudi J. Biol. Sci. 2017, 24, 338–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maděra, P.; Habrová, H.; Šenfeldr, M.; Kholová, I.; Lvončík, S.; Ehrenbergerová, L.; Roth, M.; Nadezhdina, N.; Němec, P.; Rosenthal, J.; et al. Growth dynamics of endemic Dracaena cinnabari Balf. F. of Socotra Island suggest essential elements for a conservation strategy. Biologia 2019, 74, 339–349. [Google Scholar] [CrossRef]
- Mikita, T.; Janata, P.; Surový, P. Forest Stand Inventory Based on Combined Aerial and Terrestrial Close-Range Photo-grammetry. Forests 2016, 7, 165. [Google Scholar] [CrossRef] [Green Version]
- Vahalík, P.; Drápela, K.; Procházková, A.; Patočka, Z.; Balková, M.; Šenfeldr, M.; Lengálová, K.; Kalivodová, H.; Vaníčková, L.; Ehrenbergerová, L.; et al. Metrics of Growth Habit Derived from the 3D Tree Point Cloud Used for Species Determination—A New Approach in Botanical Taxonomy Tested on Dragon Tree Group Example. Forests 2020, 11, 272. [Google Scholar] [CrossRef] [Green Version]
- Kukkonen, M.; Kotivuori, E.; Maltamo, M.; Korhonen, L.; Packalen, P. Volumes by tree species can be predicted using photogrammetric UAS data, Sentinel-2 images and prior field measurements. Silva Fenn. 2021, 55, 10360. [Google Scholar] [CrossRef]
- Novo, A.; Patočka, Z.; Cibulka, M.; Vahalík, P. Use of UAV in inventory of an old orchard—Case study Světlá. Eur. J. Hortic. Sci. 2023, 88, 0060. [Google Scholar] [CrossRef]
- Scholte, P.; De Geest, P. The climate of Socotra Island (Yemen): A first-time assessment of the timing of the monsoon wind reversal and its influence on precipitation and vegetation patterns. J. Arid Environ. 2010, 74, 1507–1515. [Google Scholar] [CrossRef]
- Banu, T.; Borlea, G.; Banu, C. The Use of Drones in Forestry. J. Environ. Sci. Eng. B 2016, 5, 557–562. [Google Scholar] [CrossRef] [Green Version]
- Guimarães, N.; Pádua, L.; Marques, P.; Silva, N.; Peres, E.; Sousa, J.J. Forestry Remote Sensing from Unmanned Aerial Vehicles: A Review Focusing on the Data, Processing and Potentialities. Remote Sens. 2020, 12, 1046. [Google Scholar] [CrossRef] [Green Version]
- Lvončík, S.; Vahalík, P.; Bongers, F.; Peijnenburg, J.; Hušková, K.; van Rensburg, J.J.; Hamdiah, S.; Maděra, P. Development of a population of Boswellia elongata Balf. F. in Homhil nature sanctuary, Socotra island (Yemen). Rend. Lincei. Sci. Fis. E Nat. 2020, 31, 747–759. [Google Scholar] [CrossRef]
- Adolt, R.; Maděra, P.; Abraham, J.; Čupa, P.; Svátek, M.; Matula, R.; Šebesta, J.; Čermák, M.; Volařík, D.; Koutecký, T.; et al. Field survey of Dracaena cinnabari populations in Firmihin, Socotra Island: Methodology and preliminary results. J. Landsc. Ecol. 2013, 6, 7–34. [Google Scholar] [CrossRef] [Green Version]
- IUCN. The IUCN Red List of Threatened Species. Version 2022-2. 2022. Available online: https://www.iucnredlist.org (accessed on 28 December 2022).
- Riccardi, T.; Malatesta, L.; Van Damme, K.; Suleiman, A.S.; Farcomeni, A.; Rezende, M.; Vahalík, P.; Attorre, F. Environmental factors and human activity as drivers of tree cover and density on the Island of Socotra, Yemen. Rend. Lincei. Sci. Fis. E Nat. 2020, 31, 703–718. [Google Scholar] [CrossRef]
- Al-Okaishi, A. Exploring the historical distribution of Dracaena cinnabari using ethnobotanical knowledge on Socotra Island, Yemen. J. Ethnobiol. Ethnomed. 2021, 17, 22. [Google Scholar] [CrossRef]
- Scholte, P.; Miller, A.; Shamsan, A.R.; Suleiman, A.S.; Taleb, N.; Millroy, T.; Attorre, F.; Porter, R.; Carugati, C.; Pella, F. Goats: Part of the Problem or the Solution to Biodiversity Conservation on Socotra? Technical Report; SCDP: Hadibo, Socotra, Republic of Yemen, 2007; p. 11. [Google Scholar]
- Schweiger, A.H.; Irl, S.D.H.; Svenning, J.; Higgins, S.I. Dynamic management needs for long-lived, sporadically recruiting plant species in human-dominated landscapes. Plants People Planet 2020, 2, 186–200. [Google Scholar] [CrossRef]
- Sopko, P.; Falvey, R. Annual Tropical Cyclone Report; Joint Typhoon Warning Center: Washington, DC, USA, 2016; p. 2, Retrieved 31 March 2020. [Google Scholar]
- Hubálková, I. Prediction of Dragon’s Blood Tree (Dracaena Cinnabari Balf.) Stand Sample Density on Soqotra Island. J. Landsc. Ecol. 2011, 4, 5–17. [Google Scholar] [CrossRef] [Green Version]
- Habrová, H.; Cermak, Z.; Pavlis, J. Dragon’s blood tree—Threatened by overmaturity, not by extinction: Dynamics of a Dracaena cinnabari woodland in the mountains of Soqotra. Biol. Conserv. 2019, 142, 772–778. [Google Scholar] [CrossRef]
- Arens, N.C.; West, I.D. Press-Pulse: A General Theory of Mass Extinction? Paleobiology 2008, 34, 456–471. [Google Scholar] [CrossRef]
- Gatenby, R.A.; Zhang, J.; Brown, J.S. First Strike–Second Strike Strategies in Metastatic Cancer: Lessons from the Evolutionary Dynamics of Extinction. Cancer Res. 2019, 79, 3174–3177. [Google Scholar] [CrossRef] [Green Version]
- Rau, E.; Gardiner, B.A.; Fischer, F.J.; Maréchaux, I.; Joetzjer, E.; Sun, I.; Chave, J. Wind Speed Controls Forest Structure in a Subtropical Forest Exposed to Cyclones: A Case Study Using an Individual-Based Model. Front. For. Glob. Chang. 2022, 5, 753100. [Google Scholar] [CrossRef]
- Maděra, P.; Van Damme, K. 2022. Socotra Archipelago (Yemen). In Imperiled: The Encyclopedia of Conservation; DellaSala, D.A., Goldstein, I.G., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 267–281. [Google Scholar] [CrossRef]
- Rejžek, M.; Svátek, M.; Šebesta, J.; Adolt, R.; Maděra, P.; Matula, R. Loss of a single tree species will lead to an overall decline in plant diversity: Effect of Dracaena cinnabari Balf. f. on the vegetation of Socotra Island. Biol. Conserv. 2016, 196, 165–172. [Google Scholar] [CrossRef]
- García, C.; Vasconcelos, R. The beauty and the beast: Endemic mutualistic interactions promote community-based conservation on Socotra Island (Yemen). J. Nat. Conserv. 2017, 35, 20–23. [Google Scholar] [CrossRef]
- González-Castro, A.; Pérez-Pérez, D.; Romero, J.; Nogales, M. Unraveling the Seed Dispersal System of an Insular “Ghost” Dragon Tree (Dracaena draco) in the Wild. Front. Ecol. Evol. 2019, 7, 39. [Google Scholar] [CrossRef] [Green Version]
- Kalivodová, H.; Culek, M.; Čermák, M.; Maděra, P.; Habrová, H. Potential importance of Socotra dragon’s blood tree cloud forests and woodlands for capturing horizontal precipitation. Rend. Lincei. Sci. Fis. E Nat. 2020, 31, 607–621. [Google Scholar] [CrossRef]













Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vahalík, P.; Van Damme, K.; Nétek, R.; Habrová, H.; Tulková, J.; Lengálová, K.; Zejdová, L.; Avoiani, E.; Maděra, P. UAV Inventory of the Last Remaining Dragon Tree Forest on Earth. Forests 2023, 14, 766. https://doi.org/10.3390/f14040766
Vahalík P, Van Damme K, Nétek R, Habrová H, Tulková J, Lengálová K, Zejdová L, Avoiani E, Maděra P. UAV Inventory of the Last Remaining Dragon Tree Forest on Earth. Forests. 2023; 14(4):766. https://doi.org/10.3390/f14040766
Chicago/Turabian StyleVahalík, Petr, Kay Van Damme, Rostislav Nétek, Hana Habrová, Jana Tulková, Klára Lengálová, Lucie Zejdová, Elizaveta Avoiani, and Petr Maděra. 2023. "UAV Inventory of the Last Remaining Dragon Tree Forest on Earth" Forests 14, no. 4: 766. https://doi.org/10.3390/f14040766
APA StyleVahalík, P., Van Damme, K., Nétek, R., Habrová, H., Tulková, J., Lengálová, K., Zejdová, L., Avoiani, E., & Maděra, P. (2023). UAV Inventory of the Last Remaining Dragon Tree Forest on Earth. Forests, 14(4), 766. https://doi.org/10.3390/f14040766