
Keystone Soil Microbial Modules Associated with Priming Effect under Nitrogen- and Glucose-Addition Treatments

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Soil Sampling
2.2. Laboratory-Based Sample Processing and Gas Sampling
2.3. CO2 Analysis and PE
2.4. Soil Physicochemical Analysis
2.5. Microbial Biomass and Enzyme Activity
2.6. Analysis of Phospholipid Fatty Acids (PLFAs)
2.7. DNA Extraction, PCR Amplification, and Sequencing
2.8. Processing of Sequencing Data
2.9. Statistical Analysis
3. Results
3.1. Physicochemical Properties of Soil
3.2. Cumulative CO2 Emission and PE
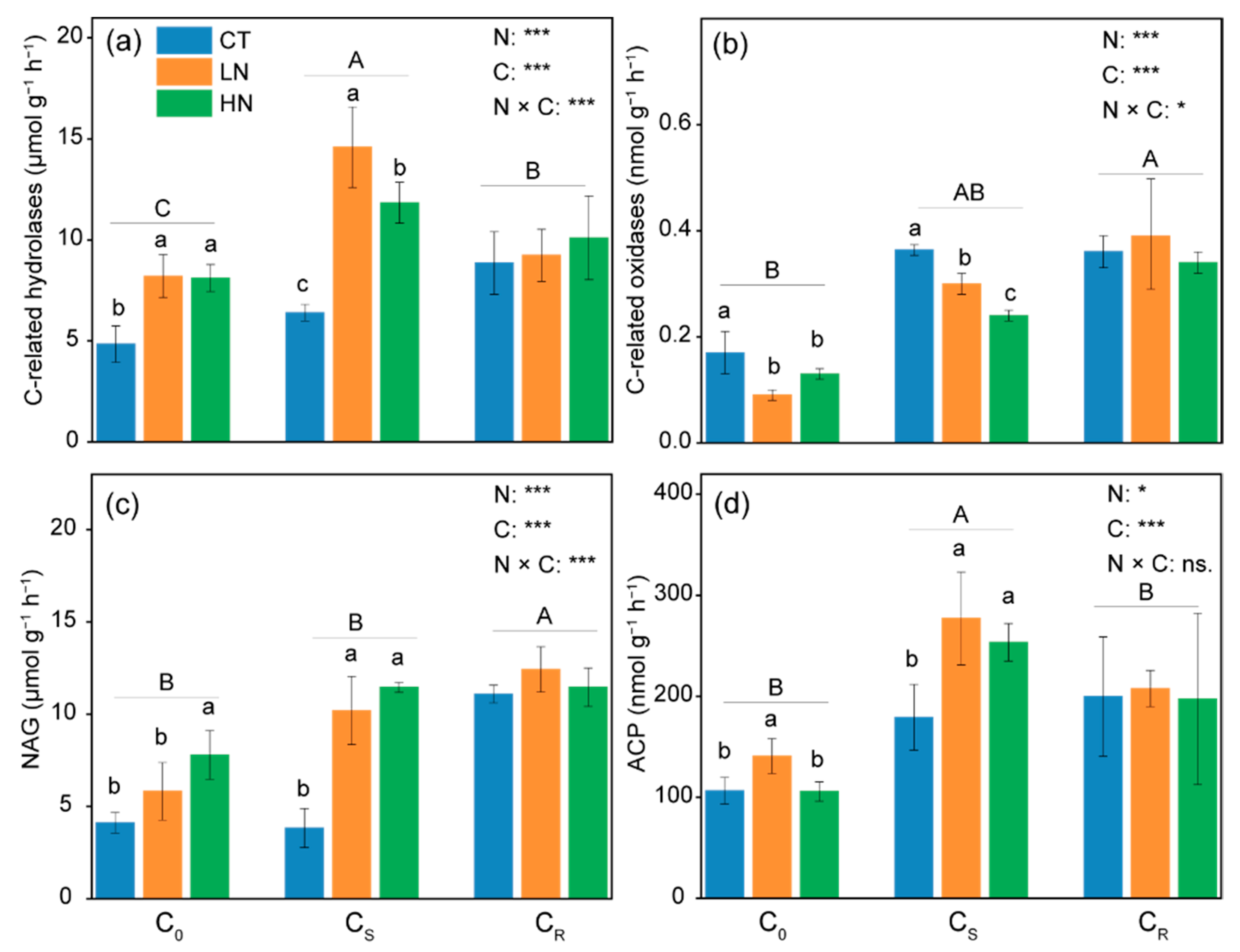
3.3. Extracellular Enzyme Activity and Microbial Biomass in Soil
3.4. Distribution of 13C-PLFA in Microbial Groups
3.5. Diversity and Community Composition of Soil Bacteria and Fungi
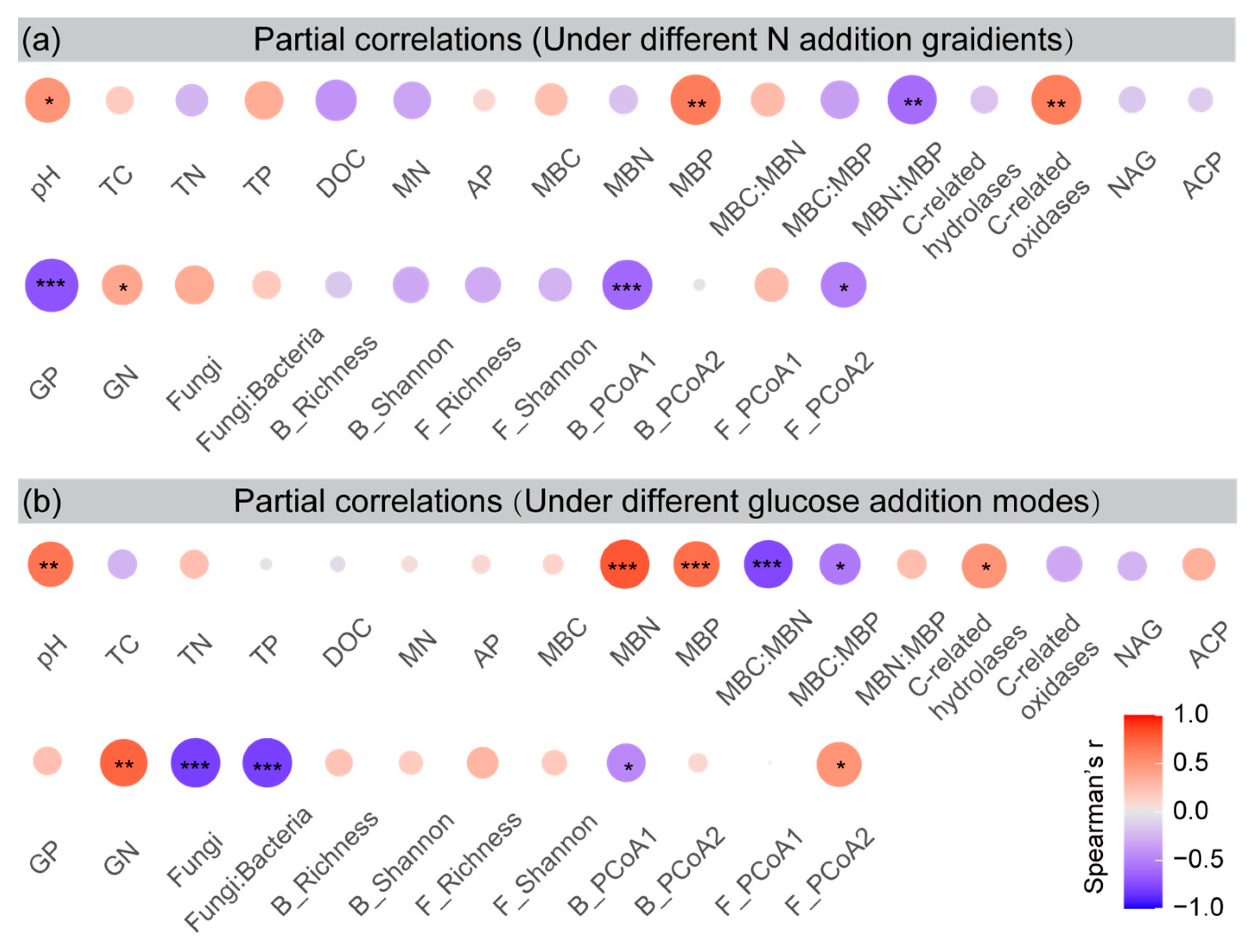
3.6. Predictors of Cumulative PE
3.7. Microbial Assemblies and Their Relationships with Cumulative PE
3.8. Co-Occurrence of Bacteria and Fungi Using Network Analysis
4. Discussion
4.1. Effects of N- and C-Addition on Cumulative CO2 Emission and PE
4.2. Drivers of Cumulative PE under N-Addition Treatment
4.3. Drivers of Cumulative PE under Different C-Addition Modes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kuzyakov, Y.; Friedel, J.K.; Stahr, K. Review of mechanisms and quantification of priming effects. Soil Biol. Biochem. 2000, 32, 1485–1498. [Google Scholar] [CrossRef]
- Dijkstra, F.A.; Carrillo, Y.; Pendall, E.; Morgan, J.A. Rhizosphere priming: A nutrient perspective. Front. Microbiol. 2013, 4, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fanin, N.; Alavoine, G.; Bertrand, I. Temporal dynamics of litter quality, soil properties and microbial strategies as main drivers of the priming effect. Geoderma 2020, 377, 114576. [Google Scholar] [CrossRef]
- Bastida, F.; García, C.; Fierer, N.; Eldridge, D.J.; Bowker, M.A.; Abades, S.; Alfaro, F.D.; Asefaw Berhe, A.; Cutler, N.A.; Gallardo, A.; et al. Global ecological predictors of the soil priming effect. Nat. Commun. 2019, 10, 3481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Razanamalala, K.; Razafifimbelo, T.; Maron, P.A.; Ranjard, L.; Chemidlin, N.; Lelievre, M.; Dequiedt, S.; Ramaroson, V.H.; Marsden, C.; Becquer, T.; et al. Soil microbial diversity drives the priming effect along climate gradients: A case study in Madagascar. ISME J. 2018, 12, 451–462. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Cheng, L.; Feng, J.; Mei, K.; Zeng, Q.; Zhu, B.; Chen, Y. Nitrogen addition stimulates priming effect in a subtropical forest soil. Soil Biol. Biochem. 2021, 160, 108339. [Google Scholar] [CrossRef]
- Fu, X.; Song, Q.; Li, S.; Shen, Y.; Yue, S. Dynamic changes in bacterial community structure are associated with distinct priming effect patterns. Soil Biol. Biochem. 2022, 169, 108671. [Google Scholar] [CrossRef]
- Zhao, F.; Wang, J.; Li, Y.; Xu, X.; He, L.; Wang, J.; Ren, C.; Guo, Y. Microbial functional genes driving the positive priming effect in forest soils along an elevation gradient. Soil Biol. Biochem. 2022, 165, 108498. [Google Scholar] [CrossRef]
- Ma, B.; Wang, H.; Dsouza, M.; Lou, J.; He, Y.; Dai, Z.; Brookes, P.C.; Xu, J.; Gilbert, J.A. Geographic patterns of co-occurrence network topological features for soil microbiota at continental scale in eastern China. ISME J. 2016, 10, 1891–1901. [Google Scholar] [CrossRef]
- Feng, J.; Zeng, X.M.; Zhang, Q.; Zhou, X.Q.; Liu, Y.R.; Huang, Q. Soil microbial trait-based strategies drive metabolic efficiency along an altitude gradient. ISME J. Commun. 2021, 1, 71. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.M.; Feng, J.; Chen, J.; Delgado-Baquerizo, M.; Zhang, Q.; Zhou, X.Q.; Yuan, Y.; Feng, S.; Zhang, K.; Liu, Y.R.; et al. Microbial assemblies associated with temperature sensitivity of soil respiration along an altitudinal gradient. Sci. Total Environ. 2022, 820, 153257. [Google Scholar] [CrossRef]
- Nottingham, A.T.; Turner, B.L.; Stott, A.W.; Tanner, E.V.J. Nitrogen and phosphorus constrain labile and stable carbon turnover in lowland tropical forest soils. Soil Biol. Biochem. 2015, 80, 26–33. [Google Scholar] [CrossRef]
- Chen, R.; Senbayram, M.; Blagodatsky, S.; Myachina, O.; Dittert, K.; Lin, X.; Blagodatskaya, E.; Kuzyakov, Y. Soil C and N availability determine the priming effect: Microbial N mining and stoichiometric decomposition theories. Glob. Chang. Biol. 2014, 20, 2356–2367. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.G.; Tang, M.; Zhu, B. Soil priming effect and its responses to nutrient addition along a tropical forest elevation gradient. Glob. Chang. Biol. 2021, 27, 2793–2806. [Google Scholar] [CrossRef]
- Zhu, Z.; Ge, T.; Luo, Y.; Liu, S.; Xu, X.; Tong, C.; Shibistova, O.; Guggenberger, G.; Wu, J. Microbial stoichiometric flexibility regulates rice straw mineralization and its priming effect in paddy soil. Soil Biol. Biochem. 2018, 121, 67–76. [Google Scholar] [CrossRef]
- Ren, C.; Zhao, F.; Shi, Z.; Chen, J.; Han, X.; Yang, G.; Feng, Y.; Ren, G. Differential responses of soil microbial biomass and carbon-degrading enzyme activities to altered precipitation. Soil Biol. Biochem. 2017, 115, 1–10. [Google Scholar] [CrossRef]
- Zhou, J.; Wen, Y.; Shi, L.; Marshall, M.R.; Kuzyakov, Y.; Blagodatskaya, E.; Zang, H. Strong priming of soil organic matter induced by frequent input of labile carbon. Soil Biol. Biochem. 2021, 152, 108069. [Google Scholar] [CrossRef]
- Wang, Q.; Chen, L.; Yang, Q.; Sun, T.; Li, C. Different effects of single versus repeated additions of glucose on the soil organic carbon turnover in a temperate forest receiving long-term N addition. Geoderma 2019, 341, 59–67. [Google Scholar] [CrossRef]
- Hamer, U.; Marschner, B. Priming effects in soils after combined and repeated substrate additions. Geoderma 2005, 128, 38–51. [Google Scholar] [CrossRef]
- Wei, Y.; Xiong, X.; Ryo, M.; Badgery, W.B.; Bi, Y.; Yang, G.; Zhang, Y.; Liu, N. Repeated litter inputs promoted stable soil organic carbon formation by increasing fungal dominance and carbon use efficiency. Biol. Fertil. Soils 2022, 58, 1–13. [Google Scholar] [CrossRef]
- Malik, A.A.; Chowdhury, S.; Schlager, V.; Oliver, A.; Puissant, J.; Vazquez, P.G.; Jehmlich, N.; von Bergen, M.; Grifths, R.I.; Gleixner, G. Soil fungal:bacterial ratios are linked to altered carbon cycling. Front. Microbiol. 2016, 7, 1247. [Google Scholar] [CrossRef] [Green Version]
- Müller, K.; Marhan, S.; Kandeler, E.; Poll, C. Carbon flow from litter through soil microorganisms: From incorporation rates to mean residence times in bacteria and fungi. Soil Biol. Biochem. 2017, 115, 187–196. [Google Scholar] [CrossRef]
- Esperschütz, J.; Perez-de-Mora, A.; Schreiner, K.; Welzl, G.; Bruegger, F.; Zeyer, J.; Hagedorn, F.; Munch, J.C.; Schloter, M. Microbial food web dynamics along a soil chronosequence of a glacier forefifield. Biogeosciences 2011, 8, 3283–3294. [Google Scholar] [CrossRef] [Green Version]
- Xiong, J.; Peng, F.; Sun, H.; Xue, X.; Chu, H. Divergent responses of soil fungi functional groups to short-term warming. Microb. Ecol. 2014, 68, 708–715. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zeng, Y.; Guo, C.; Bao, Y.; Lu, G.; Reinfelder, J.R.; Dang, Z. Bacterial, archaeal, and fungal community responses to acid mine drainage-laden pollution in a rice paddy soil ecosystem. Sci. Total Environ. 2018, 616, 107–116. [Google Scholar] [CrossRef] [PubMed]
- De Menezes, A.B.; Prendergast-Miller, M.T.; Richardson, A.E.; Toscas, P.; Farrell, M.; Macdonald, L.M.; Baker, G.; Wark, T.; Thrall, P.H. Network analysis reveals that bacteria and fungi form modules that correlate independently with soil parameters. Environ. Microbiol. 2015, 17, 2677–2689. [Google Scholar] [CrossRef]
- Banerjee, S.; Kirkby, C.A.; Schmutter, D.; Bissett, A.; Kirkegaard, J.A.; Richardson, A.E. Network analysis reveals functional redundancy and keystone taxa amongst bacterial and fungal communities during organic matter decomposition in an arable soil. Soil Biol. Biochem. 2016, 97, 188–198. [Google Scholar] [CrossRef]
- Li, B.B.; Roley, S.S.; Duncan, D.S.; Guo, J.; Quensen, J.F.; Yu, H.Q.; Tiedje, J.M. Long-term excess nitrogen fertilizer increases sensitivity of soil microbial community to seasonal change revealed by ecological network and metagenome analyses. Soil Biol. Biochem. 2021, 160, 108349. [Google Scholar] [CrossRef]
- Keith, A.; Singh, B.; Singh, B.P. Interactive priming of biochar and labile organic matter mineralization in a smectite-rich soil. Environ. Sci. Technol. 2011, 45, 9611–9618. [Google Scholar] [CrossRef]
- Jones, D.L.; Willett, V.B. Experimental evaluation of method to quantify dissolved organic nitrogen (DON) and dissolved organic carbon (DOC) in soil. Soil Biol. Biochem. 2006, 38, 991–999. [Google Scholar] [CrossRef]
- Olsen, S.R.; Sommers, L.E. Phosphorous. In Methods of Soil Analysis, Part 2, Chemical and Microbial Properties; Agronomy Society of America, Agronomy Monograph 9; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; Soil Science Society of America: Madison, WI, USA, 1982; pp. 403–430. [Google Scholar]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Turner, B.L.; Wrigh, S.J. The response of microbial biomass and hydrolytic enzymes to a decade of nitrogen, phosphorus, and potassium addition in a lowland tropical rain forest. Biogeochemistry 2014, 117, 115–130. [Google Scholar] [CrossRef]
- Saiya-Cork, K.R.; Sinsabaugh, R.L.; Zak, D.R. The effects of long-term nitrogen deposition on extracellular enzyme activity in an acer saccharum, forest soil. Soil Biol. Biochem. 2002, 34, 1309–1315. [Google Scholar] [CrossRef]
- German, D.P.; Weintraub, M.N.; Grandy, A.S.; Lauber, C.L.; Rinkes, Z.L.; Allison, S.D. Optimization of hydrolytic and oxidative enzyme methods for ecosystem studies. Soil Biol. Biochem. 2011, 43, 1387–1397. [Google Scholar] [CrossRef]
- Frostegård, Å.; Tunlid, A.; Bååth, E. Use and misuse of PLFA measurements in soils. Soil Biol. Biochem. 2011, 43, 1621–1625. [Google Scholar] [CrossRef]
- Wang, Q.K.; Wang, S.; He, T.X.; Liu, L.; Wu, J.B. Response of organic carbon mineralization and microbial community to leaf litter and nutrient additions in subtropical forest soils. Soil Biol. Biochem. 2014, 71, 13–20. [Google Scholar] [CrossRef]
- Qiao, N.; Wang, J.; Xu, X.; Shen, Y.; Long, X.E.; Hu, Y.; Schaefer, D.; Li, S.; Wang, H.; Kuzyakov, Y. Priming alters soil carbon dynamics during forest succession. Biol. Fertil. Soils 2019, 55, 339–350. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108, 4516–4522. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Sheng, L.; Li, Y.; Jiang, H.; Lv, Z.; Qi, W.; Luo, W. Soil labile organic carbon indicating seasonal dynamics of soil organic carbon in northeast peatland. Ecol. Indic. 2022, 138, 108847. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- Nilsson, R.H.; Larsson, K.H.; Taylor, A.F.S.; Bengtsson-Palme, J.; Jeppesen, T.S.; Schigel, D.; Kennedy, P.; Picard, K.; Glöckner, F.O.; Tedersoo, L.; et al. The UNITE database for molecular identification of fungi: Handling dark taxa and parallel taxonomic classifications. Nucleic Acids Res. 2019, 47, D259–D264. [Google Scholar] [CrossRef] [PubMed]
- Revelle, W. psych: Procedures for Psychological, Psychometric, and Personality Research; Northwestern University: Evanston, IL, USA, 2021; R Package Version 2.1.3.; Available online: https://CRAN.R-project.org/package=psych (accessed on 15 October 2021).
- Langfelder, P.; Horvath, S. Fast R functions for robust correlations and hierarchical clustering. J. Stat. Softw. 2012, 46, i11. [Google Scholar] [CrossRef] [Green Version]
- Benjamini, Y.; Krieger, A.M.; Yekutieli, D. Adaptive linear step-up procedures that control the false discovery rate. Biometrika 2006, 93, 491–507. [Google Scholar] [CrossRef]
- Tu, L.H.; Hu, T.X.; Zhang, J.; Li, X.W.; Hu, H.L.; Liu, L.; Xiao, Y.L. Nitrogen addition stimulates different components of soil respiration in a subtropical bamboo ecosystem. Soil Biol. Biochem. 2013, 58, 255–264. [Google Scholar] [CrossRef]
- Wang, X.; Li, S.; Zhu, B.; Homyak, P.M.; Chen, G.; Yao, X.; Wu, D.; Yang, Z.; Lyu, M.; Yang, Y. Long-term nitrogen deposition inhibits soil priming effects by enhancing phosphorus limitation in a subtropical forest. Glob. Chang. Biol. 2023, 1–13. [Google Scholar] [CrossRef]
- Yu, G.; Zhao, H.; Chen, J.; Zhang, T.; Cai, Z.; Zhou, G.; Li, Z.; Qiu, Z.; Wu, Z. Soil microbial community dynamics mediate the priming effects caused by in situ decomposition of fresh plant residues. Sci. Total Environ. 2020, 737, 139708. [Google Scholar] [CrossRef] [PubMed]
- Mooshammer, M.; Wanek, W.; Zechmeister-Boltenstern, S.; Richter, A. Stoichiometric imbalances between terrestrial decomposer communities and their resources: Mechanisms and implications of microbial adaptations to their resources. Front. Microbiol. 2014, 5, 22. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.; Moorhead, D.L.; Wang, X.; Xu, M.; Wang, X.; Wei, X.; Zhu, Z.; Ge, T.; Peng, S.; Zhu, B.; et al. Decreasing microbial phosphorus limitation increases soil carbon release. Geoderma 2022, 419, 115868. [Google Scholar] [CrossRef]
- Ho, A.; Di Lonardo, D.P.; Bodelier, P.L.E. Revisiting life strategy concepts in environmental microbial ecology. FEMS Microbiol. Ecol. 2017, 93, fix006. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Yu, G.; Zhang, X.; Wang, Q.; Tian, J.; Niu, S.; Tian, J.; Ge, J. Different strategies for regulating free-living N2 fixation in nutrient-amended subtropical and temperate forest soils. Appl. Soil Ecol. 2019, 136, 21–29. [Google Scholar] [CrossRef]
- Hu, X.; Liu, J.; Wei, D.; Zhou, B.; Chen, X.; Jin, J.; Liu, X.; Wang, G. Long-term application of nitrogen, not phosphate or potassium, significantly alters the diazotrophic community compositions and structures in a Mollisol in northeast China. Res. Microbiol. 2019, 170, 147–155. [Google Scholar] [CrossRef]
- Zhou, H.; Gao, Y.; Jia, X.; Wang, M.; Ding, J.; Cheng, L.; Bao, F.; Wu, B. Network analysis reveals the strengthening of microbial interaction in biological soil crust development in the Mu Us Sandy Land, northwestern China. Soil Biol. Biochem. 2020, 144, 107782. [Google Scholar] [CrossRef]
- Deng, F.; Liang, C. Revisiting the quantitative contribution of microbial necromass to soil carbon pool: Stoichiometric control by microbes and soil. Soil Biol. Biochem. 2022, 165, 108486. [Google Scholar] [CrossRef]
- Thiet, R.K.; Frey, S.D.; Six, J. Do growth yield efficiencies differ between soil microbial communities differing in fungal: Bacterial ratios? Reality check and methodological issues. Soil Biol. Biochem. 2006, 38, 837–844. [Google Scholar] [CrossRef]
- Zhou, S.; Wang, J.; Chen, L.; Wang, J.; Zhao, F. Microbial community structure and functional genes drive soil priming effect following afforestation. Sci. Total Environ. 2022, 825, 153925. [Google Scholar] [CrossRef]
- Wang, S.S.; Meade, A.; Lam, H.M.; Luo, H.W. Evolutionary timeline and genomic plasticity underlying the lifestyle diversity in rhizobiales. Msystems 2020, 5, e00438-20. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Trevathan-Tackett, S.M.; Jiang, Z.; Cui, L.; Wu, Y.; Zhang, X.; Huang, X. Nutrient loading decreases blue carbon by mediating fungi activities within seagrass meadows. Environ. Res. 2022, 212, 113280. [Google Scholar] [CrossRef]
- Challacombe, J.F.; Hesse, C.N.; Bramer, L.M.; McCue, L.A.; Lipton, M.; Purvine, S.; Nicora, C.; Gallegos-Graves, L.V.; Porras-Alfaro, A.; Kuske, C.R. Genomes and secretomes of Ascomycota fungi reveal diverse functions in plant biomass decomposition and pathogenesis. BMC Genom. 2019, 20, 1–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Štursová, M.; Žifčáková, L.; Leigh, M.B.; Burgess, R.; Baldrian, P. Cellulose utilization in forest litter and soil: Identification of bacterial and fungal decomposers. FEMS Microbiol. Ecol. 2012, 80, 735–746. [Google Scholar] [CrossRef]
- Eichlerová, I.; Homolka, L.; Žifčáková, L.; Lisá, L.; Dobiášová, P.; Baldrian, P. Enzymatic systems involved in decomposition reflects the ecology and taxonomy of saprotrophic fungi. Fungal Ecol. 2015, 13, 10–22. [Google Scholar] [CrossRef]
- Ren, C.; Wang, J.; Bastida, F.; Delgado-Baquerizo, M.; Yang, Y.; Wang, J.; Zhong, J.; Zhou, Z.; Zhang, S.; Guo, Y.; et al. Microbial traits determine soil C emission in response to fresh carbon inputs in forests across biomes. Glob. Chang. Biol. 2022, 28, 1516–1528. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.L.; Zhu, C.; Ruan, Y.; Luo, G.W.; Wang, M.; Ling, N.; Shen, Q.R.; Guo, S. Are the microbial communities involved in glucose assimilation in paddy soils treated with different fertilization regimes for three years similar? J. Soils Sediments 2018, 18, 2476–2490. [Google Scholar] [CrossRef]
- Novy, V.; Nielsen, F.; Seiboth, B.; Nidetzky, B. The influence of feedstock characteristics on enzyme production in Trichoderma reesei: A review on productivity, gene regulation and secretion profiles. Biotechnol. Biofuels 2019, 12, 238. [Google Scholar] [CrossRef] [Green Version]
- Fang, Y.; Van Zwieten, L.; Rose, M.T.; Vasileiadis, S.; Donner, E.; Vancov, T.; Rigg, J.; Weng, Z.; Lombi, E.; Drigo, B.; et al. Unraveling microbiomes and functions associated with strategic tillage, stubble, and fertilizer management. Agric. Ecosyst. Environ. 2022, 323, 107686. [Google Scholar] [CrossRef]
- Li, S.; Wang, S.; Fan, M.; Wu, Y.; Shangguan, Z. Interactions between biochar and nitrogen impact soil carbon mineralization and the microbial community. Soil Tillage Res. 2020, 196, 104437. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Property | C0 | CS | CR | Two-Way ANOVA Analysis | Among Group | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CT | LN | HN | CT | LN | HN | CT | LN | HN | N | C | N × C | C0 | CS | CR | |
| pH | 3.31 a | 3.25 b | 3.24 b | 3.33 a | 3.30 a | 3.26 b | 3.32 a | 3.26 b | 3.20 c | *** | *** | * | B | A | B |
| TC (g kg−1) | 51.02 b | 55.52 a | 52.74 b | 51.36 b | 55.49 a | 52.18 b | 57.00 | 52.92 | 53.37 | * | ns. | *** | A | A | A |
| TN (g kg−1) | 3.85 c | 4.98 a | 4.31 b | 3.68 c | 4.96 a | 4.58 b | 3.97 c | 4.22 b | 4.74 a | *** | ns. | *** | A | A | A |
| TP (g kg−1) | 0.91 | 0.80 | 0.81 | 0.89 | 0.79 | 0.82 | 0.88 | 0.82 | 0.84 | ns. | ns. | ns. | A | A | A |
| DOC (mg kg−1) | 48.22 | 45.13 | 50.48 | 43.91 b | 51.70 a | 54.90 a | 45.41 c | 50.57 b | 56.45 a | *** | ns. | ns. | A | A | A |
| MN (mg kg−1) | 227.65 a | 178.70 b | 260.44 a | 208.72 b | 176.32 b | 236.10 a | 203.47 b | 169.85 b | 232.28 a | *** | * | ns. | A | B | B |
| AP (mg kg−1) | 5.22 a | 2.70 b | 2.50 b | 4.43 a | 1.95 b | 2.60 b | 3.49 a | 2.19 b | 2.20 b | *** | ** | * | A | B | B |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, M.; Zeng, Q.; Liu, Y.; Liu, C.; Zhang, Q.; Mei, K.; Yuan, X.; Zhang, X.; Chen, Y. Keystone Soil Microbial Modules Associated with Priming Effect under Nitrogen- and Glucose-Addition Treatments. Forests 2023, 14, 1207. https://doi.org/10.3390/f14061207
Xu M, Zeng Q, Liu Y, Liu C, Zhang Q, Mei K, Yuan X, Zhang X, Chen Y. Keystone Soil Microbial Modules Associated with Priming Effect under Nitrogen- and Glucose-Addition Treatments. Forests. 2023; 14(6):1207. https://doi.org/10.3390/f14061207
Chicago/Turabian StyleXu, Min, Quanxin Zeng, Yuanyuan Liu, Chengchung Liu, Qiufang Zhang, Kongcan Mei, Xiaochun Yuan, Xiaoqing Zhang, and Yuehmin Chen. 2023. "Keystone Soil Microbial Modules Associated with Priming Effect under Nitrogen- and Glucose-Addition Treatments" Forests 14, no. 6: 1207. https://doi.org/10.3390/f14061207
APA StyleXu, M., Zeng, Q., Liu, Y., Liu, C., Zhang, Q., Mei, K., Yuan, X., Zhang, X., & Chen, Y. (2023). Keystone Soil Microbial Modules Associated with Priming Effect under Nitrogen- and Glucose-Addition Treatments. Forests, 14(6), 1207. https://doi.org/10.3390/f14061207