
Study on the Rooting Physiological Mechanism of Schisandra chinensis (Turcz.) Baill. Green-Branched Cuttings

Abstract
:1. Introduction
2. Materials and Methods
2.1. Test Materials
2.2. Experimental Design and Methods
2.2.1. Cutting Experiment with Different Exogenous Growth Regulators
2.2.2. Cutting Experiment with Different Substrates
2.2.3. External Morphology and Anatomical Structure of the Rooting Process of S. chinensis Cuttings
2.2.4. Determination of Physiological Indicators
2.2.5. Determination of Rooting Status
2.2.6. Data Processing and Analysis
3. Results and Analysis
3.1. Effects of the Different Exogenous Growth Regulators on Rooting Ability
3.2. Effects of Different Matrix Fertilizers on Rooting
3.3. Observation of Rooting External Morphology
3.4. Anatomical Characteristics of the Cutting Roots
3.5. Dynamic Changes of Enzyme Activity
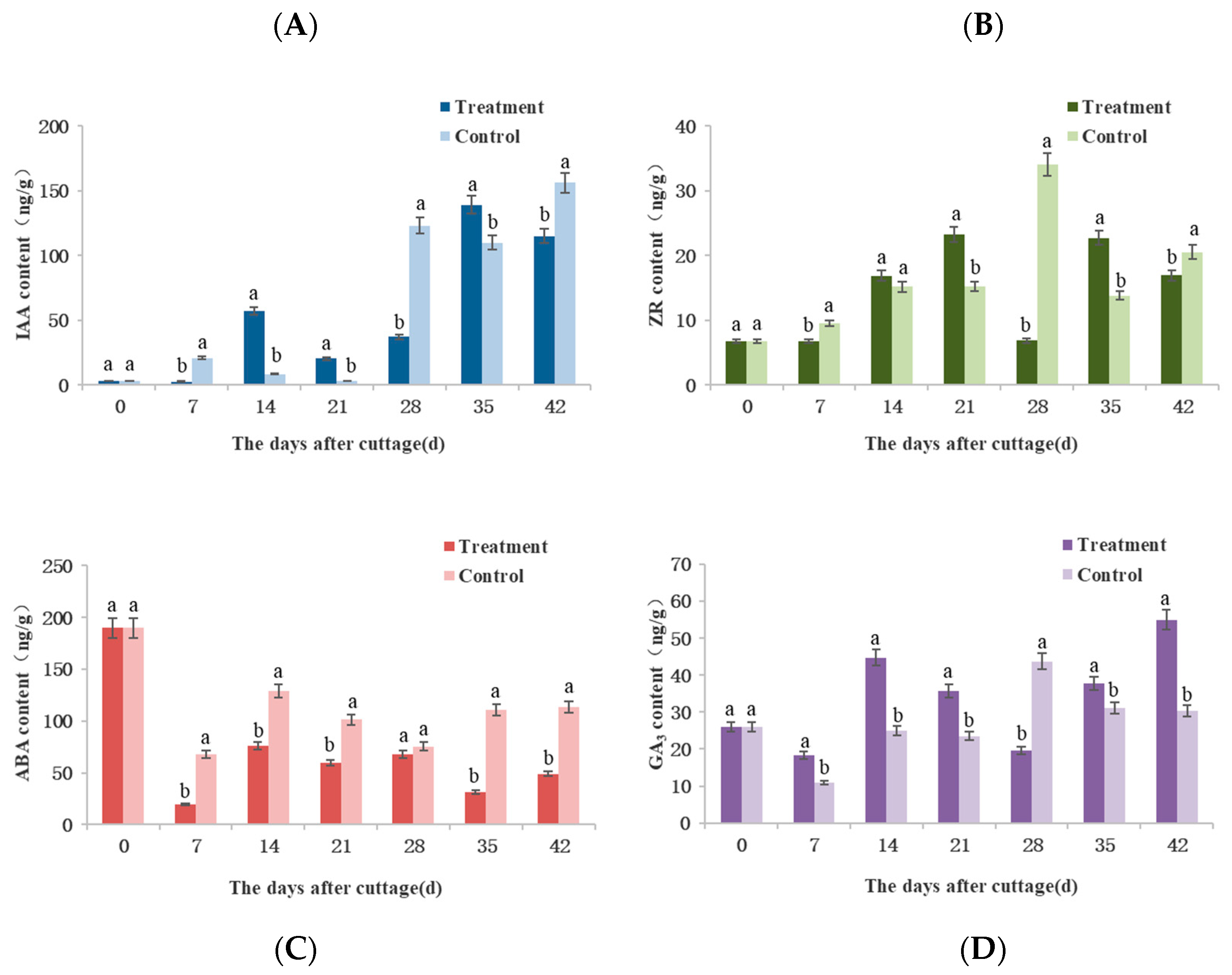
3.6. Dynamic Changes of Endogenous Growth Regulators
4. Discussion
4.1. Effects of Different Plant Growth Regulators and Substrates on the Rooting of Plant Cuttings
4.2. Anatomical Changes during Plant Cutting and Rooting
4.3. Relationship between Physiological Indexes and Rooting of S. chinensis Cuttings
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| NAA | α-Naphthalene acetic acid |
| IAA | Indole-3-acetic acid |
| ABA | Abscisic acid |
| GA3 | Gibberellic acid |
| ZR | Zeatin riboside |
| IAAO | IAA oxidase |
| POD | Peroxidase |
| PPO | Polyphenol oxidase |
| FAA | Formalin-acetic-alcohol |
References
- Ke, Y.; Jing, Q.; Zecheng, H.; Ziwei, Y.; Wenjun, W.; Huiling, H.; Yu, Y. A comprehensive review of ethnopharmacology, phytochemistry, pharmacology, and pharmacokinetics of Schisandra chinensis (Turcz.) Baill. and Schisandra sphenanthera Rehd. et Wils. J. Ethnopharmacol. 2022, 284, 114759. [Google Scholar] [CrossRef]
- Bian, Z.; Qin, Y.; Li, L.; Su, L.; Fei, C.; Li, Y.; Hu, M.; Chen, X.; Zhang, W.; Mao, C.; et al. Schisandra chinensis (Turcz.) Baill. Protects against DSS-induced colitis in mice: Involvement of TLR4/NF-κB/NLRP3 inflammasome pathway and gut microbiota. J. Ethnopharmacol. 2022, 15, 115570. [Google Scholar] [CrossRef] [PubMed]
- Chunmiao, Y.; Xiaoqian, C.; Xian, L.; Shuyu, L.; Miaomiao, Y.; Donghua, Y. Research progress of pharmacological action of Schisandra chinensis and its application in health food. Food Ind. 2022, 43, 221–227. [Google Scholar]
- Vilasboa, J.; da Costa, C.T.; Matsuura, H.N.; Fett-Neto, A.G. Rooting of cuttings of Passiflora suberosa, a medicinal passion fruit species: Characterization and modulation by external biochemical factors. Isr. J. Plant Sci. 2020, 67, 51. [Google Scholar] [CrossRef]
- Dapeng, M.; Ruirui, J.; Shenghao, L.; Shuo, X.; Ge, Z.; Shusheng, W. Research progress on adventitious root formation mechanism of woody plants. J. Zhejiang Univ. Sci. A 2022, 39, 902–912. [Google Scholar]
- Ofodile, E.; Chima, U.; Udo, E. Effect of different growth media on foliage production and root growth in Gongronema latifolia Benth stem cuttings. Greener J. Agric. Sci. 2013, 3, 215–221. [Google Scholar] [CrossRef]
- Zhou, Y.; Feng, Y.; Zhou, Q.; Jiang, T. Chamaecyparis lawsoniana with different cutting mediums and growth regulators. J. West China For. Sci. 2017, 46, 32–37. [Google Scholar]
- OuYang, F.Q.; Fu, G.Z.; Wang, J.H.; Ma, J.W.; An, S.P.; Wang, M.Q.; Li, Y. Qualitative analysis of endogenesis hormone and polyphenol during rooting of cuttingsin norway spruce (Picea abies). Sci. Silvae Sin. 2015, 51, 155–162. [Google Scholar]
- Koyama, R.; Aparecido Ribeiro Júnior, W.; Mariani Zeffa, D.; Tadeu Faria, R.; Mitsuharu Saito, H.; Simões Azeredo Gonçalves, L.; Ruffo Roberto, S. Association of Indolebutyric Acid with Azospirillum brasilense in the Rooting of Herbaceous Blueberry Cuttings. Horticulturae 2019, 5, 68. [Google Scholar] [CrossRef] [Green Version]
- Ling, M.; Xingmin, G.; Ming, T.; Zhichen, H.; Peixian, F.; Xiaorong, X. Research progress on cutting propagation and rooting mechanism of Magnoliaceae. Mol. Plant Breed. 2023, 1–21. Available online: http://kns.cnki.net/kcms/detail/46.1068.s.20220909.1738.006.html (accessed on 14 September 2022).
- Chen, B.; Li, J.; Zhang, J.; Fan, H.; Li, Q. Optimizing the Propagation of Cinnamomum micranthum by Cuttings. Life Sci. 2014, 11, 928–931. [Google Scholar]
- Xiang, Z.; Qingying, L.; Qingbin, J.; Chonglu, Z.; Jie, Z. Effects of substrates and rooting regulators on rooting of cuttings in Manglietia conifera Dandy. J. Nanjing For. Univ. 2019, 62, 23–30. [Google Scholar] [CrossRef]
- Jie, W.; Xinguo, L.; Shuoyu, A. Effects of different substrate ratios on the growth of sour pomelo seedlings. Acta Trop. Crops 2018, 39, 443–447. [Google Scholar]
- Mullah, A.; Linyu, L.; Ziyan, F.; Long, Z.; Biao, L. Study on rooting characteristics and morphological anatomy of Lonicera tatarica cuttings. Shandong Agric. Sci. 2018, 50, 48–51. [Google Scholar]
- Gelei, M.; Dongtao, L.; Zhijuan, T.; Jifeng, D.; Jianqiu, H.; Yumei, Z. Effects of Different Substrate Ratios on the Growth and Physiology of Sequoia sempervirens Container Seedlings. Compost. Sci. Util. 2019, 27, 169–177. [Google Scholar] [CrossRef]
- Mengyi, L.; Yanlin, L.; Lu, X.; Ye, F.; Dexin, G.; Xiaoying, Y. Study on cutting rooting characteristics and anatomy of Acer rubrum. J. Hunan Agr. Univ. Sci. 2019, 45, 362–367. [Google Scholar]
- Rongtao, Z.; Yang, L.; Shanshan, L. Anatomical study on adventitious root formation of Populus davidiana. Acta Bot. Northwest 2013, 33, 2184–2188. [Google Scholar]
- Yixuan, L.; Yanan, X.; Meiyu, J.; Xuanchun, P.; Meilan, L. Study on the influence of several factors on the growth of adventitious roots of Phaseolus vulgaris. J. Agr. Yanbian Univ. 2022, 44, 7–12. [Google Scholar]
- Gengxin, L.; Yide, M.; Jun, Q.; He, F.; Panfeng, L.; Qingxin, D.; Hongyan, D.; Lanying, D.; Lu, W. Anatomical and physiological changes of rooting from cuttings of Eucommia ulmoides Oliv.’ Huazhong 6’. Sci. Silvae Sin. 2022, 58, 113–124. [Google Scholar]
- Jin, H.; Zheng, C.; Hua, B.; Yu, C.; Li, K.; Yu, W. Anatomy and physiological activities of cutting roots of Torreya grandis. J. Zhejiang Agr. 2022, 34, 1955–1966. [Google Scholar] [CrossRef]
- Vilasboa, J.; Costa, C.; Ransan, L.; Mariath, J.; Fett-Neto, A. Microcutting Redox Profile and Anatomy in Eucalyptus spp. With Distinct Adventitious Rooting Competence. Front. Plant Sci. 2021, 11, 620832. [Google Scholar] [CrossRef] [PubMed]
- Chaohui, L. Study on Softwood Cutting Technology of Pteroceltis tatarinowii and Physiological and Biochemical Dynamics during Adventitious Root Formation; Shandong Agricultural University: Taian, China, 2014. [Google Scholar]
- Guoyu, H. Study on Cutting Propagation Technology Optimization and Its Mechanism of Fejoko; Southwest University of Science and Technology: Mianyang, China, 2021. [Google Scholar]
- Haishan, A.; Jiaying, Z.; Xueying, Z.; Boqiang, Z.; Liqing, Z. Changes of endogenous hormone levels during adventitious root formation in green cuttings of southern highbush blueberry (Vaccinium corymbosum L.) cv. ‘O’Neal’. J. Shanghai Agr. Sci. 2022, 6, 36–50. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Combination | NAA (ppm) | ABT (ppm) | Time (h) | |
|---|---|---|---|---|
| Treatment A | A1B1C1 | 25 | 225 | 1.5 |
| Treatment B | A1B2C2 | 25 | 250 | 2.0 |
| Treatment C | A1B3C3 | 25 | 275 | 2.5 |
| Treatment D | A2B1C2 | 50 | 225 | 2.0 |
| Treatment E | A2B2C3 | 50 | 250 | 2.5 |
| Treatment F | A2B3C1 | 50 | 275 | 1.5 |
| Treatment G | A3B1C3 | 75 | 225 | 2.5 |
| Treatment H | A3B2C1 | 75 | 250 | 1.5 |
| Treatment I | A3B3C2 | 75 | 275 | 2.0 |
| Treatment J | Control | 0 | 0 | 0 |
| Treatment | Rooting Rate | Average Root Length (cm) | Average Number of Roots (Strip) |
|---|---|---|---|
| A1B1C1 | 0.30 ± 0.09 c | 2.95 ± 1.35 b | 3.00 ± 2.00 b |
| A1B2C2 | 0.40 ± 0.08 b | 3.33 ± 0.50 ab | 3.00 ± 1.00 b |
| A1B3C3 | 0.34 ± 0.14 b | 3.25 ± 0.46 ab | 3.00 ± 1.00 b |
| A2B1C2 | 0.30 ± 0.15 c | 2.15 ± 0.20 c | 2.67 ± 0.58 c |
| A2B2C3 | 0.27 ± 0.15 d | 2.78 ± 1.12 b | 4.00 ± 1.73 a |
| A2B3C1 | 0.33 ± 0.07 b | 3.72 ± 1.11 a | 3.67 ± 2.08 ab |
| A3B1C3 | 0.60 ± 0.11 a | 3.54 ± 0.79 a | 3.67 ± 0.58 ab |
| A3B2C1 | 0.34 ± 0.09 b | 3.30 ± 1.11 ab | 3.67 ± 2.08 ab |
| A3B3C2 | 0.23 ± 0.01 d | 3.16 ± 1.36 b | 2.67 ± 1.16 c |
| Control | 0.00 ± 0.00 e | 0.00 ± 0.00 d | 0.00 ± 0.00 d |
| Matrix Number | Rooting Rate | Average Root Length (cm) | Average Number of Roots (Strip) |
|---|---|---|---|
| A | 0.79 ± 0.05 a | 3.79 ± 0.16 a | 8.00 ± 0.58 a |
| B | 0.32 ± 0.03 c | 2.28 ± 0.35 c | 3.67 ± 0.33 c |
| C | 0.74 ± 0.09 a | 3.15 ± 0.42 b | 5.67 ± 0.20 b |
| D | 0.42 ± 0.10 b | 3.11 ± 0.70 ab | 4.00 ± 1.15 c |
| E | 0.49 ± 0.01 b | 3.32 ± 0.39 ab | 5.33 ± 0.88 b |
| F | 0.74 ± 0.12 a | 2.84 ± 0.35 bc | 3.67 ± 0.33 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, D.; Zhao, X.; Ai, J.; Wang, Z.; Shi, G.; Liu, Y.; Li, X.; Zhao, C. Study on the Rooting Physiological Mechanism of Schisandra chinensis (Turcz.) Baill. Green-Branched Cuttings. Forests 2023, 14, 1365. https://doi.org/10.3390/f14071365
Sun D, Zhao X, Ai J, Wang Z, Shi G, Liu Y, Li X, Zhao C. Study on the Rooting Physiological Mechanism of Schisandra chinensis (Turcz.) Baill. Green-Branched Cuttings. Forests. 2023; 14(7):1365. https://doi.org/10.3390/f14071365
Chicago/Turabian StyleSun, Dan, Xilin Zhao, Jun Ai, Zhenxing Wang, Guangli Shi, Yumeng Liu, Xiang Li, and Chengcheng Zhao. 2023. "Study on the Rooting Physiological Mechanism of Schisandra chinensis (Turcz.) Baill. Green-Branched Cuttings" Forests 14, no. 7: 1365. https://doi.org/10.3390/f14071365
APA StyleSun, D., Zhao, X., Ai, J., Wang, Z., Shi, G., Liu, Y., Li, X., & Zhao, C. (2023). Study on the Rooting Physiological Mechanism of Schisandra chinensis (Turcz.) Baill. Green-Branched Cuttings. Forests, 14(7), 1365. https://doi.org/10.3390/f14071365