
Combining Tree-Ring Width and Density to Separate the Effects of Climate Variation and Insect Defoliation

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
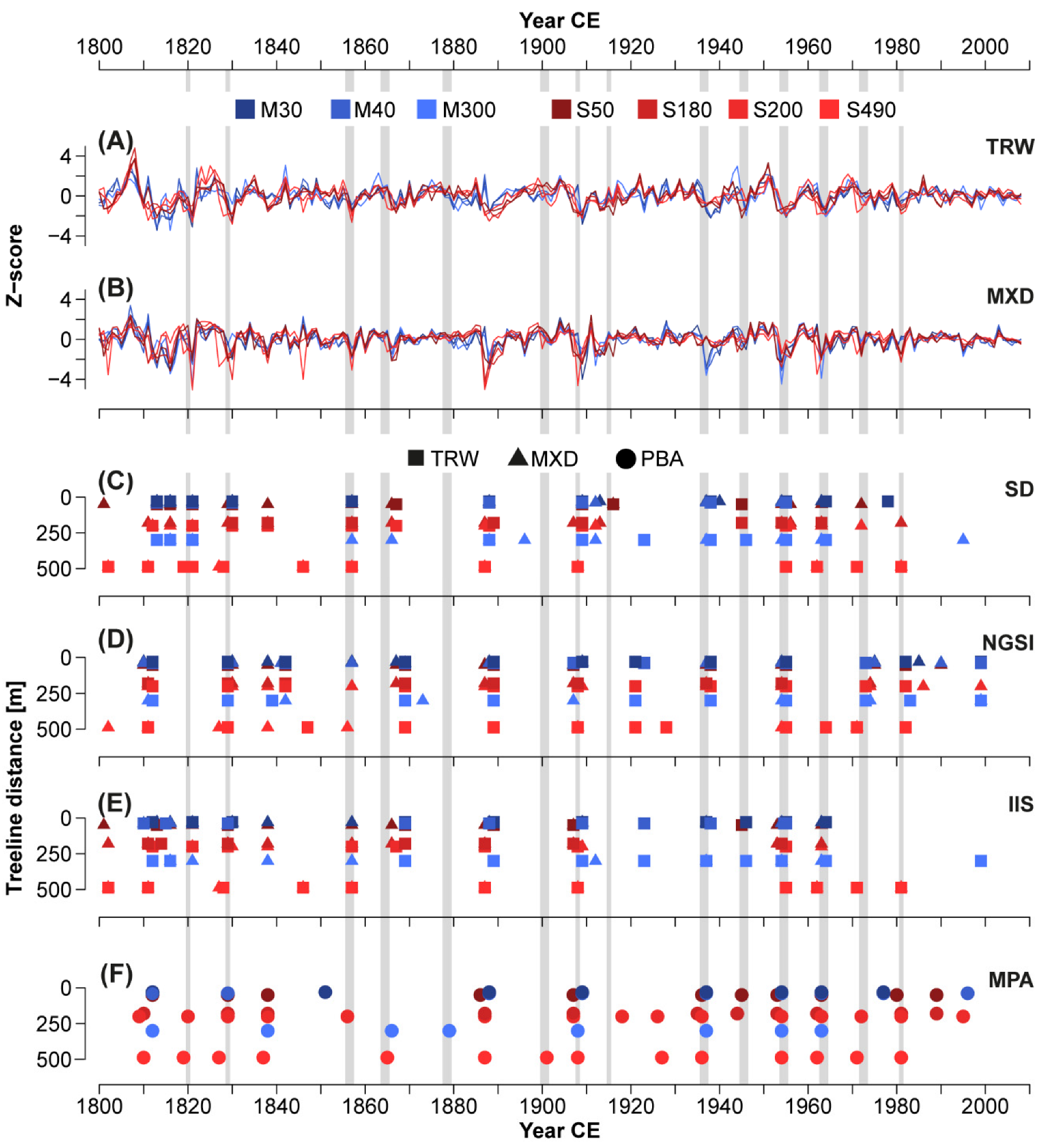
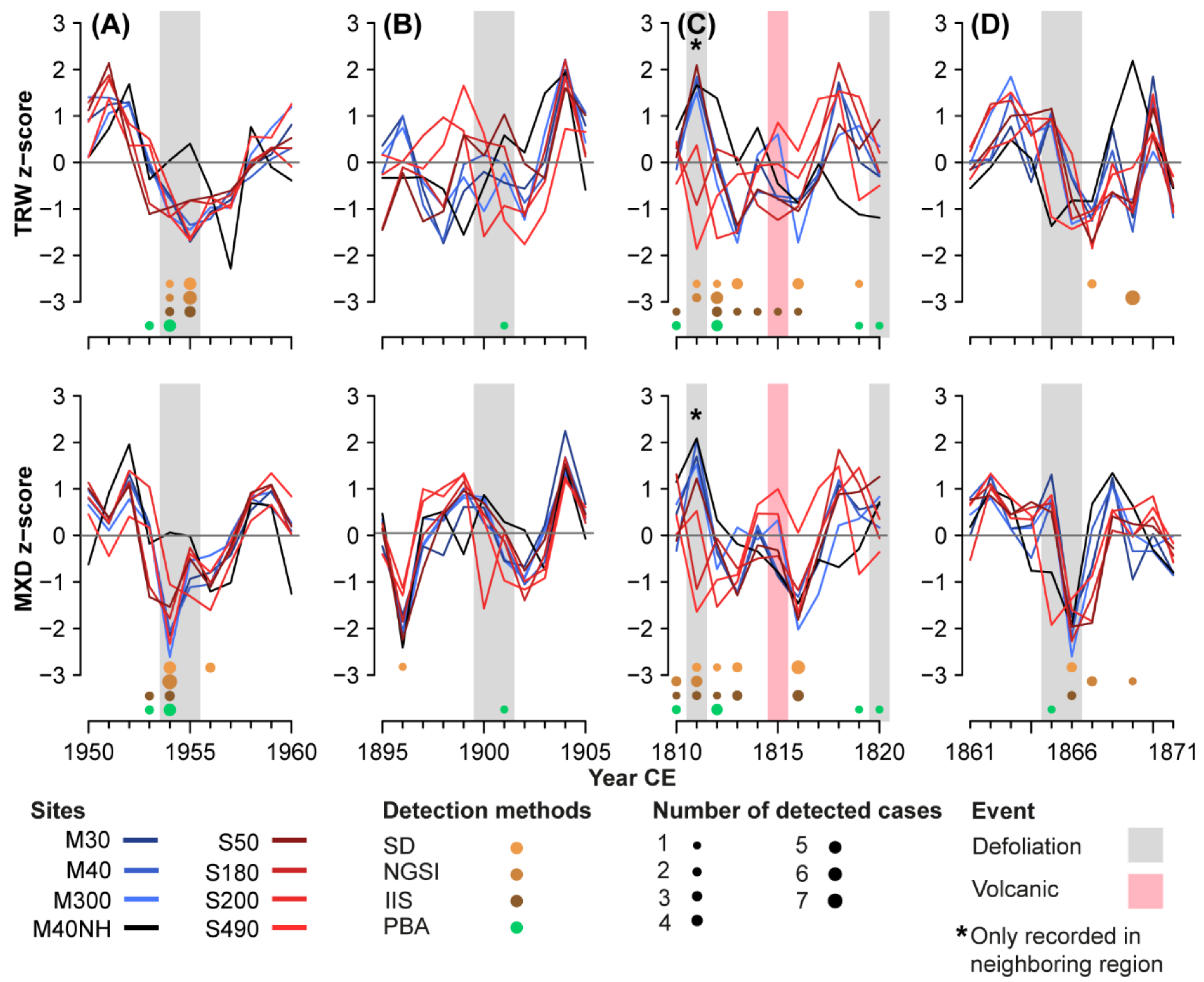
- Standard deviation (SD): The first method selects strong negative outliers exceeding 1.5 SD in both TRW and the MXD index chronologies;
- Normalized growth-suppression index (NGSI): This method was applied to single TRW and MXD larch series using the pine chronology from M40NH as a non-host reference to represent variability unrelated to LBM outbreaks. Outbreak duration was set to five years to account for the cyclicity of the LBM and avoid the identification of several events within the same cycle. The method detects outbreaks by selecting events that exceed an NGSI threshold value of −1.28, which has seen use across different species [43]. Two event years are defined per outbreak: the year in which the most single trees were defoliated according to their NGSI, and the year of the strongest outbreak-related growth departure represented by a minimum of the mean NGSI. Since the year of the minimum mean NGSI showed a higher agreement with the external defoliation data, these years were chosen for further analysis. This analysis was performed using the “dfoliatR” package in R [44];
- Impulse Indicator Saturation (IIS): Here, a regression model to detect structural breaks in time series is saturated with a maximum number of indicators, from which the non-significant ones are removed. 0.1 was chosen as the significance level for the t-tests, and the non-host pine chronology was used as an additional regressor in the model to support differentiating LBM- and non-LBM-related growth reductions. If several consecutive years were detected by this method, the one with the lowest coefficient (i.e., the strongest negative outlier) among them was chosen. This method is mainly used in econometrics but has also been successfully applied to tree-ring data [45]. We used the “gets” package in R [46];
- Our new process-based approach (PBA, Figure 2): Based on the characteristic LBM pattern in tree rings (cf. Figure 1C–E), we developed a stepwise procedure for LBM detection. First, the TRW series are analyzed to identify values exceeding 0.5 SD below the chronology mean in two consecutive years. TRW has been shown to be strongly affected for only one year in some cases and to recover already in the second year after the initial outbreak, albeit still being below average [36]. The relatively small threshold was chosen to account for these different outbreak patterns and to disregard as few actual outbreaks as possible. Any selected year that occurred within four years of the previous events was removed to account for the LBM cyclicity. As LBM populations decrease by a factor of many thousands after a mass outbreak [47,48], an LBM event cannot be followed by an additional outbreak in such a short time. Next, the residuals of a linear regression between TRW and MXD for the same years were calculated. They were checked for negative outliers exceeding 1 SD below the mean along with a reduction compared to the previous year to find the strongest outlier if consecutive years crossed the threshold. The same was done for the ratios between EWW and LWW. Outlier years in both parameters were then checked for coinciding TRW deviations. At this stage, two cases were considered an outbreak: if a negative TRW deviation year coincided with an outlier of either TRW-MXD residuals or EWW/LWW ratios, this year was defined as the first year of an LBM outbreak. Occasionally, such a TRW-MXD residual or EWW/LWW ratio event occurs one year after the onset of a negative TRW deviation period. In these cases, the year with the lowest MXD value was chosen since we considered MXD to be the most characteristic parameter for the start of an outbreak period. Moreover, we removed events occurring within four years after an existing event to obtain one initial year for each outbreak period.
3. Results
4. Discussion
4.1. Differentiation of Temperature Events from LBM Outbreaks
4.2. Elevational and Valley Differences
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Esper, J.; Cook, E.R.; Schweingruber, F.H. Low-Frequency Signals in Long Tree-Ring Chronologies for Reconstructing Past Temperature Variability. Science 2002, 295, 2250–2253. [Google Scholar] [CrossRef] [Green Version]
- Esper, J.; Frank, D.C.; Timonen, M.; Zorita, E.; Wilson, R.J.S.; Luterbacher, J.; Holzkämper, S.; Fischer, N.; Wagner, S.; Nievergelt, D.; et al. Orbital Forcing of Tree-Ring Data. Nat. Clim. Chang. 2012, 2, 862–866. [Google Scholar] [CrossRef]
- Fritts, H.C. Tree Rings and Climate; Elsevier Science: Tucson, AZ, USA, 1976. [Google Scholar]
- Affolter, P.; Büntgen, U.; Esper, J.; Rigling, A.; Weber, P.; Luterbacher, J.; Frank, D. Inner Alpine Conifer Response to 20th Century Drought Swings. Eur. J. For. Res. 2010, 129, 289–298. [Google Scholar] [CrossRef] [Green Version]
- Fritts, H.C.; Smith, D.G.; Cardis, J.W.; Budelsky, C.A. Tree-Ring Characteristics Along a Vegetation Gradient in Northern Arizona. Ecology 1965, 46, 394–401. [Google Scholar] [CrossRef]
- Hartl, C.; Düthorn, E.; Tejedor, E.; Kirchhefer, A.J.; Timonen, M.; Holzkämper, S.; Büntgen, U.; Esper, J. Micro-Site Conditions Affect Fennoscandian Forest Growth. Dendrochronologia 2021, 65, 125787. [Google Scholar] [CrossRef]
- Hartl, C.; Schneider, L.; Riechelmann, D.F.C.; Kuhl, E.; Kochbeck, M.; Klippel, L.; Büntgen, U.; Esper, J. The Temperature Sensitivity along Elevational Gradients Is More Stable in Maximum Latewood Density than Tree-Ring Width. Dendrochronologia 2022, 73, 125958. [Google Scholar] [CrossRef]
- Sprengel, P.C. Von Der Substanz der Ackerkrume und Des Untergrundes. J. Tech. Ökon. Chem. 1828, 3, 42–99. [Google Scholar]
- Stine, A.R.; Huybers, P. Implications of Liebig’s Law of the Minimum for Tree-Ring Reconstructions of Climate. Environ. Res. Lett. 2017, 12, 114018. [Google Scholar] [CrossRef]
- Büntgen, U.; Liebhold, A.; Nievergelt, D.; Wermelinger, B.; Roques, A.; Reinig, F.; Krusic, P.J.; Piermattei, A.; Egli, S.; Cherubini, P.; et al. Return of the Moth: Rethinking the Effect of Climate on Insect Outbreaks. Oecologia 2020, 192, 543–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esper, J.; Benz, M.; Pederson, N. Influence of Wood Harvest on Tree-Ring Time-Series of Picea Abies in a Temperate Forest. For. Ecol. Manag. 2012, 284, 86–92. [Google Scholar] [CrossRef] [Green Version]
- Pretzsch, H. The Emergent Past: Past Natural and Human Disturbances of Trees Can Reduce Their Present Resistance to Drought Stress. Eur. J. For. Res. 2022, 141, 87–104. [Google Scholar] [CrossRef]
- Hartl-Meier, C.; Esper, J.; Liebhold, A.; Konter, O.; Rothe, A.; Büntgen, U. Effects of Host Abundance on Larch Budmoth Outbreaks in the European Alps. Agric. For. Entomol. 2017, 19, 376–387. [Google Scholar] [CrossRef]
- Speer, J.H.; Kulakowski, D. Creating a Buzz: Insect Outbreaks and Disturbance Interactions. In Dendroecology; Amoroso, M.M., Daniels, L.D., Baker, P.J., Camarero, J.J., Eds.; Ecological Studies; Springer International Publishing: Cham, Switzerland, 2017; Volume 231, pp. 231–255. ISBN 978-3-319-61668-1. [Google Scholar]
- Büntgen, U.; Frank, D.C.; Nievergelt, D.; Esper, J. Summer Temperature Variations in the European Alps, a.d. 755–2004. J. Clim. 2006, 19, 5606–5623. [Google Scholar] [CrossRef]
- Kuhl, E.; Zang, C.; Esper, J.; Riechelmann, D.F.C.; Büntgen, U.; Briesch, M.; Reinig, F.; Römer, P.; Konter, O.; Schmidhalter, M.; et al. Using Machine Learning to Provenance the Geographical Origin of Historical Construction Timbers. Ecosphere 2023, 14, e4453. [Google Scholar] [CrossRef]
- Cole-Dai, J.; Ferris, D.; Lanciki, A.; Savarino, J.; Baroni, M.; Thiemens, M.H. Cold Decade (AD 1810–1819) Caused by Tambora (1815) and Another (1809) Stratospheric Volcanic Eruption. Geophys. Res. Lett. 2009, 36, L22703. [Google Scholar] [CrossRef] [Green Version]
- Harington, C.R. The Year without a Summer: World Climate in 1816; Canadian Museum of Nature: Ottawa, ON, USA, 1992; ISBN 978-0-660-13063-7. [Google Scholar]
- Luterbacher, J.; Pfister, C. The Year without a Summer. Nat. Geosci. 2015, 8, 246–248. [Google Scholar] [CrossRef]
- Frank, D.C.; Büntgen, U.; Böhm, R.; Maugeri, M.; Esper, J. Warmer Early Instrumental Measurements versus Colder Reconstructed Temperatures: Shooting at a Moving Target. Quat. Sci. Rev. 2007, 26, 3298–3310. [Google Scholar] [CrossRef]
- Büntgen, U.; Esper, J.; Frank, D.; Nicolussi, K.; Schmidhalter, M. A 1052-Year Tree-Ring Proxy for Alpine Summer Temperatures. Clim. Dyn. 2005, 25, 141–153. [Google Scholar] [CrossRef] [Green Version]
- Büntgen, U.; Frank, D.; Liebhold, A.; Johnson, D.; Carrer, M.; Urbinati, C.; Grabner, M.; Nicolussi, K.; Levanic, T.; Esper, J. Three Centuries of Insect Outbreaks across the European Alps. New Phytol. 2009, 182, 929–941. [Google Scholar] [CrossRef]
- Saulnier, M.; Corona, C.; Stoffel, M.; Guibal, F.; Edouard, J.-L. Climate-Growth Relationships in a Larix Decidua Mill. Network in the French Alps. Sci. Total Environ. 2019, 664, 554–566. [Google Scholar] [CrossRef]
- Baltensweiler, W.; Weber, U.M.; Cherubini, P. Tracing the Influence of Larch-Bud-Moth Insect Outbreaks and Weather Conditions on Larch Tree-Ring Growth in Engadine (Switzerland). Oikos 2008, 117, 161–172. [Google Scholar] [CrossRef]
- Baltensweiler, W.; Fischlin, A. The Larch Budmoth in the Alps. In Dynamics of Forest Insect Populations; Berryman, A.A., Ed.; Springer: Boston, MA, USA, 1988; pp. 331–351. ISBN 978-1-4899-0791-2. [Google Scholar]
- Konter, O.; Esper, J.; Liebhold, A.; Kyncl, T.; Schneider, L.; Düthorn, E.; Büntgen, U. Tree-Ring Evidence for the Historical Absence of Cyclic Larch Budmoth Outbreaks in the Tatra Mountains. Trees 2015, 29, 809–814. [Google Scholar] [CrossRef]
- Schweingruber, F.H. Auswirkungen Des Lärchenwicklerbefalls Auf Die Jahrringstruktur Der Lärche. Schweiz. Z. Forstwes. 1979, 130, 1071–1093. [Google Scholar]
- Weidner, K.; Heinrich, I.; Helle, G.; Löffler, J.; Neuwirth, B.; Schleser, G.H.; Vos, H. Consequences of Larch Budmoth Outbreaks on the Climatic Significance of Ring Width and Stable Isotopes of Larch. Trees 2010, 24, 399–409. [Google Scholar] [CrossRef]
- Esper, J.; Büntgen, U.; Frank, D.C.; Nievergelt, D.; Liebhold, A. 1200 Years of Regular Outbreaks in Alpine Insects. Proc. Biol. Sci. 2007, 274, 671–679. [Google Scholar] [CrossRef]
- Rozenberg, P.; Pâques, L.; Huard, F.; Roques, A. Direct and Indirect Analysis of the Elevational Shift of Larch Budmoth Outbreaks along an Elevation Gradient. Front. For. Glob. Chang. 2020, 3, 86. [Google Scholar] [CrossRef]
- Moser, L.; Fonti, P.; Büntgen, U.; Esper, J.; Luterbacher, J.; Franzen, J.; Frank, D. Timing and Duration of European Larch Growing Season along Altitudinal Gradients in the Swiss Alps. Tree Physiol. 2010, 30, 225–233. [Google Scholar] [CrossRef]
- Pallardy, S. Physiology of Woody Plants; Academic Press: New York, NY, USA, 2018. [Google Scholar]
- Esper, J.; Schneider, L.; Smerdon, J.E.; Schöne, B.R.; Büntgen, U. Signals and Memory in Tree-Ring Width and Density Data. Dendrochronologia 2015, 35, 62–70. [Google Scholar] [CrossRef] [Green Version]
- Arbellay, E.; Jarvis, I.; Chavardès, R.D.; Daniels, L.D.; Stoffel, M. Tree-Ring Proxies of Larch Bud Moth Defoliation: Latewood Width and Blue Intensity Are More Precise than Tree-Ring Width. Tree Physiol. 2018, 38, 1237–1245. [Google Scholar] [CrossRef] [Green Version]
- Ryerson, D.E.; Swetnam, T.W.; Lynch, A.M. A Tree-Ring Reconstruction of Western Spruce Budworm Outbreaks in the San Juan Mountains, Colorado, U.S.A. Can. J. For. Res. 2003, 33, 1010–1028. [Google Scholar] [CrossRef]
- Weber, U.M. Dendroecological Reconstruction and Interpretation of Larch Budmoth (Zeiraphera diniana) Outbreaks in Two Central Alpine Valleys of Switzerland from 1470–1990. Trees 1997, 11, 277–290. [Google Scholar] [CrossRef]
- Rinn, F. TSAP-WinTM; Rinntech: Heidelberg, Germany, 2003. [Google Scholar]
- Holmes, R.L. Computer-Assisted Quality Control in Tree-Ring Dating and Measurement. Tree-Ring Bull. 1983, 43, 69–78. [Google Scholar]
- Björklund, J.; Arx, G.; Nievergelt, D.; Wilson, R.; van den Bulcke, J.; Günther, B.; Loader, N.J.; Rydval, M.; Fonti, P.; Scharnweber, T.; et al. Scientific Merits and Analytical Challenges of Tree–Ring Densitometry. Rev. Geophys. 2019, 57, 1224–1264. [Google Scholar] [CrossRef] [Green Version]
- Cook, E.R. A Time Series Analysis Approach to Tree Ring Standardization; University of Arizona: Tucson, AZ, USA, 1985. [Google Scholar]
- Bunn, A.G. A Dendrochronology Program Library in R (DplR). Dendrochronologia 2008, 26, 115–124. [Google Scholar] [CrossRef]
- Schulz, M.; Mudelsee, M. REDFIT: Estimating Red-Noise Spectra Directly from Unevenly Spaced Paleoclimatic Time Series. Comput. Geosci. 2002, 28, 421–426. [Google Scholar] [CrossRef]
- Lynch, A.M. What Tree-Ring Reconstruction Tells Us about Conifer Defoliator Outbreaks. In Insect Outbreaks Revisited; Blackwell Publishing Ltd.: Hoboken, NJ, USA, 2012; pp. 126–154. ISBN 978-1-118-29520-5. [Google Scholar]
- Guiterman, C.H.; Lynch, A.M.; Axelson, J.N. DfoliatR: An R Package for Detection and Analysis of Insect Defoliation Signals in Tree Rings. Dendrochronologia 2020, 63, 125750. [Google Scholar] [CrossRef]
- Schneider, L.; Smerdon, J.E.; Pretis, F.; Hartl-Meier, C.; Esper, J. A New Archive of Large Volcanic Events over the Past Millennium Derived from Reconstructed Summer Temperatures. Environ. Res. Lett. 2017, 12, 094005. [Google Scholar] [CrossRef]
- Pretis, F.; Reade, J.J.; Sucarrat, G. Automated General-to-Specific (GETS) Regression Modeling and Indicator Saturation for Outliers and Structural Breaks. J. Stat. Softw. 2018, 86, 1–44. [Google Scholar] [CrossRef]
- Baltensweiler, W.; Benz, G.; Bovey, P.; Delucchi, V. Dynamics of Larch Bud Moth Populations. Annu. Rev. Entomol. 1977, 22, 79–100. [Google Scholar] [CrossRef]
- Turchin, P.; Wood, S.N.; Ellner, S.P.; Kendall, B.E.; Murdoch, W.W.; Fischlin, A.; Casas, J.; McCauley, E.; Briggs, C.J. Dynamical Effects of Plant Quality and Parasitism on Population Cycles of Larch Budmoth. Ecology 2003, 84, 1207–1214. [Google Scholar] [CrossRef] [Green Version]
- Baltensweiler, W.; Rübli, D. Dispersal: An Important Driving Force of the Cyclic Population Dynamics of the Larch Bud Moth, Zeiraphera diniana Gn. For. Snow Landsc. Res. 1999, 74, 3–153. [Google Scholar]
- Chree, C. Some Phenomena of Sunspots and of Terrestrial Magnetism at Kew Observatory. Philos. Trans. R. Soc. Lond. Ser. A 1913, 212, 75–116. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Bieber, A. Die Bekämpfung Des Grauen Lärchenwicklers in Den Wäldern Des Oberengadins. Prism. Illus. Monatsschrift Nat. Forsch. Tech. 1946, 1, 23–27. [Google Scholar]
- Reichen, L.; Burgdorf, A.-M.; Brönnimann, S.; Franke, J.; Hand, R.; Valler, V.; Samakinwa, E.; Brugnara, Y.; Rutishauser, T. A Decade of Cold Eurasian Winters Reconstructed for the Early 19th Century. Nat. Commun. 2022, 13, 2116. [Google Scholar] [CrossRef] [PubMed]
- Büntgen, U.; Trnka, M.; Krusic, P.J.; Kyncl, T.; Kyncl, J.; Luterbacher, J.; Zorita, E.; Ljungqvist, F.C.; Auer, I.; Konter, O.; et al. Tree-Ring Amplification of the Early Nineteenth-Century Summer Cooling in Central Europe. J. Clim. 2015, 28, 5272–5288. [Google Scholar] [CrossRef] [Green Version]
- Barbey, A. Traité D’entomologie Forestière; Berger-Levrault: Nancy, France; Paris, France; Strasbourg, France, 1925. [Google Scholar]
- Esper, J.; Büntgen, U.; Luterbacher, J.; Krusic, P.J. Testing the Hypothesis of Post-Volcanic Missing Rings in Temperature Sensitive Dendrochronological Data. Dendrochronologia 2013, 31, 216–222. [Google Scholar] [CrossRef]
- Esper, J.; Schneider, L.; Krusic, P.J.; Luterbacher, J.; Büntgen, U.; Timonen, M.; Sirocko, F.; Zorita, E. European Summer Temperature Response to Annually Dated Volcanic Eruptions over the Past Nine Centuries. Bull. Volcanol. 2013, 75, 736. [Google Scholar] [CrossRef] [Green Version]
- Coaz, J. Über Das Auftreten des Grauen Lärchenwicklers (Steganoptycha pinicola Zell.) in der Schweiz und Den Angrenzenden Staaten; Stämpfli: Bern, Switzerland, 1894. [Google Scholar]
- Kress, A.; Saurer, M.; Büntgen, U.; Treydte, K.S.; Bugmann, H.; Siegwolf, R.T.W. Summer Temperature Dependency of Larch Budmoth Outbreaks Revealed by Alpine Tree-Ring Isotope Chronologies. Oecologia 2009, 160, 353–365. [Google Scholar] [CrossRef] [Green Version]
- Rolland, C.; Baltensweiler, W.; Petitcolas, V. The Potential for Using Larix Decidua Ring Widths in Reconstructions of Larch Budmoth (Zeiraphera diniana) Outbreak History: Dendrochronological Estimates Compared with Insect Surveys. Trees 2001, 15, 414–424. [Google Scholar] [CrossRef]
- MeteoSchweiz. Available online: https://www.meteoschweiz.admin.ch/service-und-publikationen/applikationen/messwerte-und-messnetze.html (accessed on 14 March 2023).
- Baltensweiler, W. Why the Larch Bud-Moth Cycle Collapsed Why the Larch Bud-Moth Cycle Collapsed in the Subalpine Larch-Cembran Pine Forests in the Year 1990 for the First Time since 1850. Oecologia 1993, 94, 62–66. [Google Scholar] [CrossRef]
- Johnson, D.M.; Büntgen, U.; Frank, D.C.; Kausrud, K.; Haynes, K.J.; Liebhold, A.M.; Esper, J.; Stenseth, N.C. Climatic Warming Disrupts Recurrent Alpine Insect Outbreaks. Proc. Natl. Acad. Sci. USA 2010, 107, 20576–20581. [Google Scholar] [CrossRef] [PubMed]
- Bjørnstad, O.N.; Peltonen, M.; Liebhold, A.M.; Baltensweiler, W. Waves of Larch Budmoth Outbreaks in the European Alps. Science 2002, 298, 1020–1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wermelinger, B.; Forster, B.; Nievergelt, D. Cycles and Importance of the Larch Budmoth. WSL Fact Sheet 2018, 61, 12. [Google Scholar]
- Johnson, D.M.; Bjørnstad, O.N.; Liebhold, A.M. Landscape Geometry and Travelling Waves in the Larch Budmoth. Ecol. Lett. 2004, 7, 967–974. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Species | Region | Exp. | Elev. [m a.s.l.] | Period | Rep. | MSL | AGR [mm] | Mean MXD [g/cm3] | AR1 TRW | AR1 MXD | Rbar TRW | Rbar MXD |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| M30 | Larix decidua | Matter Valley | NW | 2270 | 1396–2017 | 30 | 342 | 0.55 | 0.79 | 0.25 | 0.14 | 0.66 | 0.4 |
| M40 | Larix decidua | Matter Valley | NE | 2262 | 1430–2017 | 24 | 331 | 0.59 | 0.8 | 0.25 | 0.16 | 0.73 | 0.43 |
| M300 | Larix decidua | Matter Valley | N | 2000 | 1705–2017 | 22 | 154 | 1.45 | 0.9 | 0.35 | 0.12 | 0.76 | 0.39 |
| S50 | Larix decidua | Simplon Valley | W | 2150 | 1542–2009 | 24 | 218 | 0.65 | 0.82 | 0.41 | 0.18 | 0.56 | 0.37 |
| S180 | Larix decidua | Simplon Valley | W | 2020 | 1672–2009 | 22 | 212 | 0.67 | 0.85 | 0.54 | 0.42 | 0.69 | 0.47 |
| S200 | Larix decidua | Simplon Valley | NE | 1900 | 1582–2009 | 23 | 205 | 0.79 | 0.9 | 0.45 | 0.2 | 0.49 | 0.4 |
| S490 | Larix decidua | Simplon Valley | W | 1713 | 1641–2009 | 26 | 169 | 0.81 | 0.93 | 0.57 | 0.3 | 0.6 | 0.34 |
| M40NH | Pinus cembra | Matter Valley | NE | 2262 | 1596–2017 | 31 | 264 | 0.77 | 0.6 | 0.23 | 0.12 | 0.57 | 0.28 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kunz, M.; Esper, J.; Kuhl, E.; Schneider, L.; Büntgen, U.; Hartl, C. Combining Tree-Ring Width and Density to Separate the Effects of Climate Variation and Insect Defoliation. Forests 2023, 14, 1478. https://doi.org/10.3390/f14071478
Kunz M, Esper J, Kuhl E, Schneider L, Büntgen U, Hartl C. Combining Tree-Ring Width and Density to Separate the Effects of Climate Variation and Insect Defoliation. Forests. 2023; 14(7):1478. https://doi.org/10.3390/f14071478
Chicago/Turabian StyleKunz, Marcel, Jan Esper, Eileen Kuhl, Lea Schneider, Ulf Büntgen, and Claudia Hartl. 2023. "Combining Tree-Ring Width and Density to Separate the Effects of Climate Variation and Insect Defoliation" Forests 14, no. 7: 1478. https://doi.org/10.3390/f14071478
APA StyleKunz, M., Esper, J., Kuhl, E., Schneider, L., Büntgen, U., & Hartl, C. (2023). Combining Tree-Ring Width and Density to Separate the Effects of Climate Variation and Insect Defoliation. Forests, 14(7), 1478. https://doi.org/10.3390/f14071478