
Characterization of Soil Microbial Biomass Carbon and Nitrogen in Four Forest Types of Shushan Urban Forest Park


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Set-Up and Sample Collection
2.2. Soil Physicochemical Property Analyses
2.3. Soil MBC and MBN Analyses
2.4. Statistical Analysis
3. Results
3.1. Soil Physicochemical Properties
3.2. Soil MBC and MBN Characterizations
3.3. Impact of Forest Types and Soil Layers on Soil Properties and Microbial Biomass
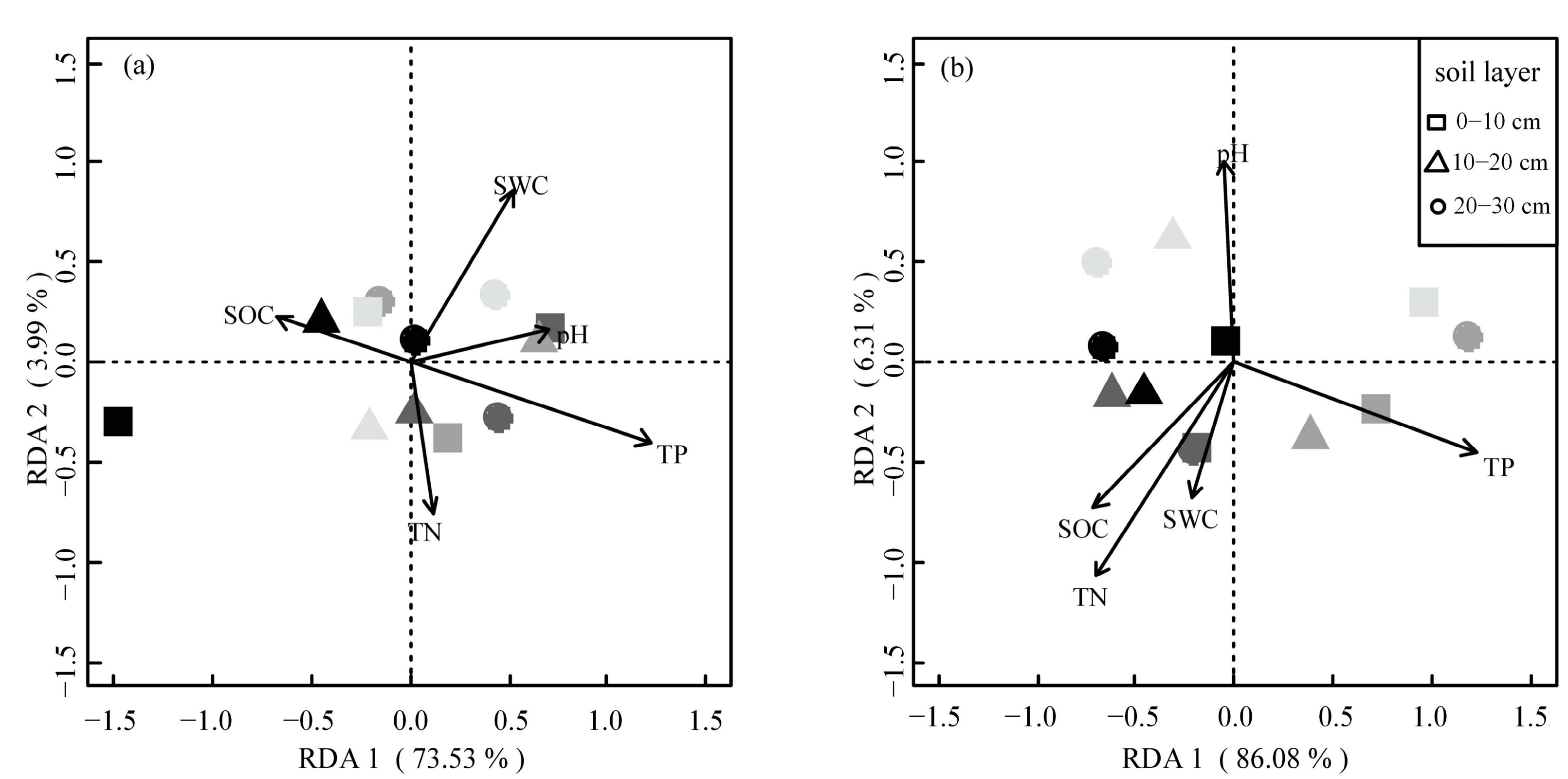
3.4. Relationships between Soil Physicochemical Properties and Microbial Biomass
4. Discussion
4.1. Effects of Different Forest Types on Soil Physicochemical Characteristics
4.2. Effects of Soil Physicochemical Factors on Soil Microbial Biomass
4.3. Forest Type and Soil Layer Interactions Influence Soil Properties and Microbial Biomass
4.4. Implications of Soil Physicochemical Properties on Soil MBC and MBN
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ordóñez, C.; Duinker, P.N. An analysis of urban forest management plans in Canada: Implications for urban forest management. Landsc. Urban Plan. 2013, 116, 36–47. [Google Scholar] [CrossRef] [Green Version]
- Steenberg, J.W.N.; Millward, A.A.; Nowakd, D.J.; Robinson, P.J.; Ellis, A. Forecasting urban forest ecosystem structure, function, and vulnerability. Environ. Manag. 2016, 59, 373–392. [Google Scholar] [CrossRef]
- Duinker, P.N.; Greig, L.A.; Kenney, W.A.; Yawney, J.K. Forecasting urban forest ecosystem structure, function, and vulnerability. Urban For. Urban Green. 2015, 14, 890–901. [Google Scholar]
- Han, Q.; Keeffe, G. Promoting climate-driven forest migration through large-scale urban afforestation. Landsc. Urban Plan. 2021, 212, 104059. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, Y.; Cheng, J.; Xue, L.; Yan, J. Effects of afforestation on carbon sequestration and global climate change: A review. For. Ecol. Manag. 2021, 482, 118802. [Google Scholar]
- Li, Z.; Tian, D.; Wang, B.; Wang, J.; Wang, S.; Chen, H.Y.H.; Xu, X.; Wang, C.; He, N.; Niu, S. Microbes drive global soil nitrogen mineralization and availability. Glob. Change Biol. 2019, 25, 1078–1088. [Google Scholar] [CrossRef]
- Yan, M.Y. Research on the Microbial Decomposition and Nutrient Release Feature of the Pinus Massoniana Forest Litters in Jinyun Mountain. Master's Thesis, South West University, Chongqing, China, 2011. [Google Scholar]
- Fan, X.G.; Jin, K.; Li, Z.J.; Rong, X.N. Soil microbial diversity under different fertilization and tillage practices: A review. J. Plant Nutr. Fertil. Sci. 2010, 16, 744–751. [Google Scholar]
- Qiu, T.T.; Liu, G.B.; Wang, G.L.; Sun, L.P.; Yao, X. Changes of soil microbial biomass carbon and their impact factors for Pinus tabuliformis plantations at different development stages on the Loess Plateau, China. Chin. J. Appl. Ecol. 2016, 27, 681–687. [Google Scholar]
- Zak, D.R.; Holmes, W.E.; White, D.C.; Peacock, A.D.; Tilman, D. Plant diversity, soil microbial communities, and ecosystem function: Are there any links? Ecology 2003, 84, 2042–2050. [Google Scholar] [CrossRef] [Green Version]
- Li, F.S.; Fang, X.; Xiang, W.H.; Sun, W.J.; Zhang, S.J. Soil Microbial Biomass Carbon and Nitrogen Concentrations in Four Subtropical Forests in Hilly Region of Central Hunan Province, China. Sci. Silvae Sin. 2014, 50, 8–16. [Google Scholar]
- Zhao, T.; Yan, H.; Jang, Y.L.; Huang, Y.M.; An, S.S. Effects of vegetation types on soil microbial biomass C,N,P on the Loess Hilly Area. Acta Ecol. Sin. 2013, 33, 5615–5622. [Google Scholar] [CrossRef]
- Liang, C.; Amelung, W.; Lehmann, J.; Kästner, M. Quantitative assessment of microbial necromass contribution to soil organic matter. Global Change Biology. 2019, 25, 3578–3590. [Google Scholar] [CrossRef] [PubMed]
- Wardle, D.A. A comparative assessment of factors which influence microbial biomass carbon and nitrogen levels in soil. Biol. Rev. 1992, 67, 321–358. [Google Scholar] [CrossRef]
- Groffman, P.M.; McDowell, W.H.; Myers, J.C.; Pastor, J.; Carpenter, S.R.; Nadelhoffer, K.J. Soil microbial biomass and activity in tropical riparian forests. Soil Biol. Biochem. 2001, 33, 339–348. [Google Scholar] [CrossRef]
- Zeng, D.H.; Hu, Y.L.; Chang, S.X.; Fan, Z.P.; Li, Y.L. Land cover change effects on soil chemical and biological properties after planting Mongolian pine (Pinus sylvestris var. mongolica) in sandy lands in Keerqin, northeastern China. Plant Soil 2009, 317, 121–133. [Google Scholar] [CrossRef]
- Burton, J.; Chen, C.R.; Xu, Z.H.; Mathers, N.J.; Xu, D.Z. Gross nitrogen transformations in adjacent native and plantation forests of subtropical Australia. Soil Biol. Biochem. 2007, 39, 426–433. [Google Scholar] [CrossRef]
- Liu, S.R.; Li, X.M.; Niu, L.M. The degradation of soil fertility in pure larch plantations in the northeastern of China. Ecol. Eng. 1998, 10, 75–86. [Google Scholar] [CrossRef]
- Behera, N.; Sahani, U. Soil microbial biomass and activity in response to Eucalyptus plantation and natural regeneration on tropical soil. For. Ecol. Manag. 2003, 174, 1–11. [Google Scholar] [CrossRef]
- Singh, M.K.; Ghoshal, N. Variation in soil microbial biomass in the dry tropics: Impact of land-use change. Soil Res. 2014, 52, 299–306. [Google Scholar] [CrossRef]
- Hua, F.; Bruijnzeel, L.A.; Meli, P.; Martin, P.A.; Zhang, J.; Nakagawa, S.; Miao, X.; Wang, W.; McEvoy, C.; Peña-Arancibia, J.L.; et al. The biodiversity and ecosystem service contributions and trade-offs of forest restoration approaches. Science 2022, 376, 839–844. [Google Scholar] [CrossRef]
- Rousk, J.; Baath, E.; Brookes, P.C. Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J. 2010, 4, 1340–1351. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N.; Jackson, R. The diversity and biogeography of soil bacterial communities. Proc. Natl. Acad. Sci. USA 2006, 103, 626–631. [Google Scholar] [CrossRef] [PubMed]
- Liang, G.H.; Wu, P.; Xiong, X.; Wu, X.Y.; Chu, G.W. Responses of Soil pH Value and Soil Microbial Biomass Carbon and Nitrogen to Simulated Acid Rain in Three Successional Subtropical Forests at Dinghushan Nature Reserve. Ecol. Environ. Sci. 2015, 24, 911–918. [Google Scholar]
- Griffiths, R.I.; Thomson, B.C.; James, P.; Bell, T.; Bailey, M.J. The bacterial biogeography of British soils. Environ. Microbiol. 2011, 13, 1642–1654. [Google Scholar] [CrossRef]
- Xiao, R.H.; Man, X.L.; Ding, L.Z. Effects of slope position on soil microbial biomass carbon and nitrogen in natural Pinus sylvestris var. mongolia forest in the cold temperature zone. J. Beijing For. Univ. 2020, 42, 31–39. [Google Scholar]
- Day, M.Y.; Chuang, W.C. The contribution of soil organic matter fractions to carbon and nitrogen mineralization and microbial community size and structure. Soil Biol. Biochem. 2016, 37, 1726–1737. [Google Scholar]
- Li, H.; Liu, J.; Yang, L.; Zheng, H.; Zhang, J. Effects of simulated climate warming on soil microbial biomass carbon, nitrogen and phosphorus of alpine forest. Chin. J. Appl. Environ. Biol. 2016, 22, 426–432. [Google Scholar]
- Pan, F.J.; Zhang, W.; Liang, Y.M.; Liu, S.J.; Wang, K.L. Increased associated effects of topography and litter and soil nutrients on soil enzyme activities and microbial biomass along vegetation successions in karst ecosystem, southwestern China. Environ. Sci. Pollut. Res. 2018, 25, 16979–16990. [Google Scholar] [CrossRef] [PubMed]
- Daniel, C.S.; Matthew, G.B.; James, M.B.; Linda, L.K. Plant community richness and microbial interactions structure bacterial communities in soil. Ecol. A Publ. Ecol. Soc. Am. 2015, 96, 134–142. [Google Scholar]
- Sugihara, S.; Funakawa, S.; Kilasara, M.; Kosaki, T. Effect of land management and soil texture on seasonal variations in soil microbial biomass in dry tropical agroecosystems in Tanzania. Appl. Soil Ecol. 2010, 44, 80–88. [Google Scholar] [CrossRef]
- Endreny, T.A. Strategically growing the urban forest will improve our world. Nat. Commun. 2018, 9, 1160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, H.M.; Tang, G.G. Characteristics of the deciduous broad-leaf forest of Tilia breviradiata in the hill terrain of Dashushan Mountain. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2009, 033, 35–40. [Google Scholar]
- Cheng, H.M. Geographical composition of vascular plants in Dashu mountain in Hefei, Anhui province. Plant Sci. J. 2011, 29, 288–295. [Google Scholar]
- Cheng, H.M.; Tian, K.; Tian, X.J. Species diversity in successional communities of vegetation in isolated island-liked hilly fragment of Dashu Mountain in Hefei, Anhui. Chin. J. Ecol. 2015, 34, 1830–1837. [Google Scholar]
- Zhang, Y.Q.; Hou, L.Y.; Li, Z.C.; Zhao, D.X.; Song, L.G.; Shao, G.D.; Ai, J.J.; Sun, Q.W. Leguminous supplementation increases the resilience of soil microbial community and nutrients in Chinese fir plantations. Sci. Total Environ. 2020, 703, 134917.1–134917.11. [Google Scholar] [CrossRef]
- Su, D.H.; Zhou, J.W.; Yin, Z.C.; Feng, H.B.; Zheng, X.M.; Han, X.; Hou, Q.Q. A calculation method of unsaturated soil water content based on thermodynamic equilibrium. Hydrol. Earth Syst. Sci. 2023, 44, 1–12. [Google Scholar]
- Wu, J.; Joergensen, R.G.; Pommerening, B. Measurement of soil microbial biomass C by fumigation-extraction—An automated procedure. Soil Biol. Biochem. 1990, 22, 1167–1169. [Google Scholar] [CrossRef]
- Vance, E.D.; Brooks, P.C.; Jenkinson, D.S. An Extraction Method for Measuring Soil Microbial Biomass. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Brookes, P.C.; Landman, A.; Pruden, G.; Jenkonson, D.S. Chloroform fumigation and release of soil N: A rapid direct extraction method to measure microbial biomass N in soil. Soil Biol. Biochem. 1985, 17, 837–842. [Google Scholar] [CrossRef]
- Snow, G.L. ggplot2: Elegant Graphics for Data Analysis. Am. Stat. 2011, 65, 204. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O'Hara, B.; Simpson, G.L.; Solymos, P.; Stevens, H.; Wagner, H.H. Vegan: Community Ecology Package. R Package Version 2.2-1 2015. [Google Scholar]
- Li, Y.; Zhao, Y.; Li, Y.; Wang, X.; Li, C. Effects of afforestation on soil properties in China: A meta-analysis. Land Use Policy 2018, 76, 52–61. [Google Scholar]
- Wang, H.; Wang, W.; Zhang, B.; Yang, G.; Wang, X. Effects of long-term reforestation on soil properties in the Loess Plateau, China. Forests 2019, 10, 28. [Google Scholar]
- Zhang, Y.; Zhao, Y.; Liu, J.; Wang, Y.; Li, S. Coniferous plants affect soil acidification and nutrient cycling in a subtropical forest ecosystem. For. Ecol. Manag. 2021, 491, 119179. [Google Scholar]
- Liu, Z.; Li, Y.; Li, X.; Zhang, X. Effects of vegetation type on soil acidification in the mid-subtropical region of China. Catena 2019, 177, 252–259. [Google Scholar]
- Ding, L.; Wang, X.; Wang, L.; Wang, H.; Guan, D. Effects of vegetation type and forest age on soil water-holding capacity in the Loess Plateau of China. Forests 2018, 9, 537. [Google Scholar]
- Liu, J.; Zhang, Y.; Wang, Y.; Li, S.; Zhao, Y. Effects of vegetation type on soil water content and water use efficiency in a subtropical forest ecosystem. J. For. Res. 2020, 31, 2283–2294. [Google Scholar]
- Jia, M.; Liu, J.; Zhang, Y.; Wang, Y.; Li, S. Comparison of soil carbon and nitrogen characteristics in different forest types in a subtropical region of China. For. Ecol. Manag. 2019, 451, 117530. [Google Scholar]
- Zhang, Y.; Zhao, Y.; Wang, Y.; Li, S.; Liu, J. Effect of vegetation type on soil organic carbon and total nitrogen stocks in a subtropical f49orest ecosystem. Catena 2019, 173, 445–453. [Google Scholar]
- Liu, J.; Zhang, Y.; Wang, Y.; Li, S.; Zhao, Y. Effects of vegetation type on soil nutrient content and stoichiometry in a subtropical forest ecosystem. For. Ecol. Manag. 2017, 391, 9–17. [Google Scholar]
- Wang, Y.; Liu, X.S.; Chen, F.F.; Huang, R.L.; Deng, X.J.; Jiang, Y. Seasonal dynamics of soil microbial biomass C and N of Keteleeria fortune var. cyclolepis forests with different ages. J. For. Res. 2020, 31, 2377–2384. [Google Scholar] [CrossRef] [Green Version]
- Farooq, T.H.; Chen, X.; Shakoor, A.; Rashid, M.H.U.; Kumar, U.; Alhomrani, M.; Alamri, A.S.; Ravindran, B.; Yan, W. Unraveling the Importance of Forest Structure and Composition Driving Soil Microbial and Enzymatic Responses in the Subtropical Forest Soils. Forests 2022, 13, 1535. [Google Scholar] [CrossRef]
- Yang, K.; Zhang, Y.; Liu, J.; Zhang, Q. Relationships between soil microbial biomass and soil properties in different forest types in the Qinling Mountains. Ecol. Indic. 2018, 85, 1050–1057. [Google Scholar]
- Zhang, H.; Liu, J.; Guo, D.; Chen, H.; Wang, F.; Li, X. Soil microbial biomass and community structure vary across different forest types in Northeast China. For. Ecol. Manag. 2019, 432, 651–661. [Google Scholar]
- Zhang, Y.; Li, X.; Liu, J.; Yang, K.; Wang, G. Soil microbial biomass and its relationship with soil properties in different forest types in the Loess Plateau, China. Ecol. Indic. 2021, 121, 107147. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Quadrat | Elevation (m) | Longitude (°) | Latitude (°) | Slope Aspect | Mean DBH 1 (cm) | Mean Tree Height (m) | Dominant Species |
|---|---|---|---|---|---|---|---|
| P. massoniana (PM) | 258 | 117.18825 | 31.84305 | Southeast | 24.62 ± 4.37 | 28.24 ± 3.67 | Pinus massoniana Lamb. Broussonetia papyrifera (Linn.) L'Hér. ex Vent. Cudrania tricuspidata (Carr.) Bur. |
| Q. acutissima (QA) | 194 | 117.18885 | 31.84830 | Northeast | 38.15 ± 5.97 | 27.54 ± 3.23 | Quercus acutissima Carruth. Broussonetia papyrifera (Linn.) L'Hér. ex Vent. |
| L. formosana (LF) | 217 | 117.18879 | 31.85080 | Northeast | 25.63 ± 10.54 | 28.36 ± 8.16 | Liquidambar formosana Hance Sapium sebiferum (L.) Roxb. |
| secondary decidu-ous broadleaved forest (DB) | 226 | 117.18770 | 31.85227 | Northeast | 14.44 ± 4.35 | 10.29 ± 3.04 | Celtis sinensis Pers. Cudrania tricuspidata (Carr.) Bur. Broussonetia papyrifera (Linn.) L'Hér. ex Vent. |
| Soil Layer (cm) | Forest Type | pH | EC (µS/cm) | SWC (%) | SOC (g/kg) | TN (g/kg) | TP (mg/kg) |
|---|---|---|---|---|---|---|---|
| 0–10 | PM | 5.07 ± 0.64 ab | 80.25 ± 42.51 ab | 23.08 ± 7.89 b | 26.61 ± 11.41 b | 1.68 ± 0.67 b | 0.22 ± 0.12 a |
| QA | 4.37 ± 0.34 c | 108.23 ± 82.88 a | 25.02 ± 3.66 b | 35.21 ± 11.31 a | 2.38 ± 0.62 a | 0.21 ± 0.04 a | |
| LF | 5.03 ± 0.48 b | 76.67 ± 33.92 b | 28.68 ± 4.70 a | 20.38 ± 9.55 c | 1.36 ± 0.53 c | 0.15 ± 0.06 b | |
| DB | 5.55 ± 0.55 a | 96.93 ± 49.00 ab | 30.28 ± 5.13 a | 23.39 ± 8.29 bc | 1.89 ± 0.49 b | 0.20 ± 0.05 a | |
| 10–20 | PM | 5.34 ± 0.53 a | 54.67 ± 29.08 b | 26.50 ± 7.85 bc | 17.31 ± 8.50 b | 1.26 ± 0.54 b | 0.21 ± 0.16 a |
| QA | 4.58 ± 0.49 b | 103.96 ± 96.34 a | 23.73 ± 3.61 c | 21.35 ± 7.68 a | 1.54 ± 0.39 a | 0.17 ± 0.04 ab | |
| LF | 5.27 ± 0.56 a | 60.60 ± 17.72 b | 27.54 ± 4.76 ab | 14.20 ± 4.48 b | 1.07 ± 0.32 b | 0.15 ± 0.08 b | |
| DB | 5.60 ± 0.54 a | 75.18 ± 26.46 b | 29.31 ± 4.60 a | 17.41 ± 4.33 b | 1.56 ± 0.26 a | 0.18 ± 0.05 ab | |
| 20–30 | PM | 5.57 ± 0.49 ab | 59.20 ± 20.93 b | 29.28 ± 7.77 a | 14.81 ± 6.58 b | 1.14 ± 0.41 b | 0.18 ± 0.12 a |
| QA | 4.88 ± 0.51 c | 81.31 ± 51.56 a | 25.08 ± 4.91 b | 19.36 ± 8.29 a | 1.42 ± 0.41 a | 0.18 ± 0.13 a | |
| LF | 5.49 ± 0.51 b | 62.39 ± 15.84 b | 28.16 ± 4.98 a | 12.23 ± 4.10 b | 0.98 ± 0.30 b | 0.13 ± 0.06 b | |
| DB | 5.83 ± 0.48 a | 72.56 ± 29.33 ab | 30.61 ± 5.63 a | 15.01 ± 4.46 b | 1.50 ± 0.53 a | 0.16 ± 0.04 ab |
| Factor | Growing Season | Dormant Season | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Depth | Forest Type | Depth × Forest Type | Depth | Forest Type | Depth × Forest Type | |||||||
| F | p | F | p | F | p | F | p | F | p | F | p | |
| pH | 15.083 | 0.000 *** | 51.285 | 0.000 *** | 0.273 | 0.949 | 11.287 | 0.000 *** | 37.152 | 0.000 *** | 0.511 | 0.799 |
| Electrical Conductivity (EC) | 9.278 | 0.000 *** | 21.488 | 0.000 *** | 1.157 | 0.332 | 1.209 | 0.301 | 3.974 | 0.009 ** | 0.794 | 0.576 |
| Soil water content (SWC) | 1.169 | 0.313 | 12.743 | 0.000 *** | 0.671 | 0.673 | 1.1 | 0.335 | 0.864 | 0.461 | 0.975 | 0.444 |
| Soil organic carbon (SOC) | 40.639 | 0.000 *** | 7.727 | 0.000 *** | 1.363 | 0.232 | 4.028 | 0.02 * | 2.809 | 0.041 * | 0.823 | 0.554 |
| Total nitrogen (TN) | 39.668 | 0.000 *** | 17.1 | 0.000 *** | 1.493 | 0.183 | 15.212 | 0.000 *** | 31.807 | 0.000 *** | 0.198 | 0.977 |
| Total phosphorus (TP) | 2.874 | 0.059 | 42.426 | 0.000 *** | 1.117 | 0.354 | 6.206 | 0.003 ** | 28.548 | 0.000 *** | 0.286 | 0.943 |
| Microbial biomass carbon (MBC) | 29.813 | 0.000 *** | 31.087 | 0.000 *** | 1.766 | 0.109 | 2.372 | 0.096 | 6.454 | 0.000 *** | 1.699 | 0.124 |
| Microbial biomass nitrogen (MBN) | 33.964 | 0.000 *** | 7.359 | 0.000 *** | 5.326 | 0.000 *** | 13.595 | 0.000 *** | 22.789 | 0.000 *** | 3.812 | 0.001 *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, M.; Cui, J.; Liu, H.; Xu, X. Characterization of Soil Microbial Biomass Carbon and Nitrogen in Four Forest Types of Shushan Urban Forest Park. Forests 2023, 14, 1498. https://doi.org/10.3390/f14071498
Wang M, Cui J, Liu H, Xu X. Characterization of Soil Microbial Biomass Carbon and Nitrogen in Four Forest Types of Shushan Urban Forest Park. Forests. 2023; 14(7):1498. https://doi.org/10.3390/f14071498
Chicago/Turabian StyleWang, Mimi, Jun Cui, Haiyang Liu, and Xiaoniu Xu. 2023. "Characterization of Soil Microbial Biomass Carbon and Nitrogen in Four Forest Types of Shushan Urban Forest Park" Forests 14, no. 7: 1498. https://doi.org/10.3390/f14071498
APA StyleWang, M., Cui, J., Liu, H., & Xu, X. (2023). Characterization of Soil Microbial Biomass Carbon and Nitrogen in Four Forest Types of Shushan Urban Forest Park. Forests, 14(7), 1498. https://doi.org/10.3390/f14071498