
Mycorrhizal Fungi Reclamation Promotes Stoichiometric Homeostasis of Re-Vegetation Types and Affects Soil Bacterial Function in Mining Subsidence of Northern Loess Plateau


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Location and Sampling Design
2.2. Determination of Soil Physicochemical Properties
2.3. Soil Microbial Sequencing
2.4. Biological Information and Data Analysis
3. Results
3.1. Ecological Stoichiometry in Root and Leaf under Re-Vegetation Types and Mycorrhizal Fungi Reclamation
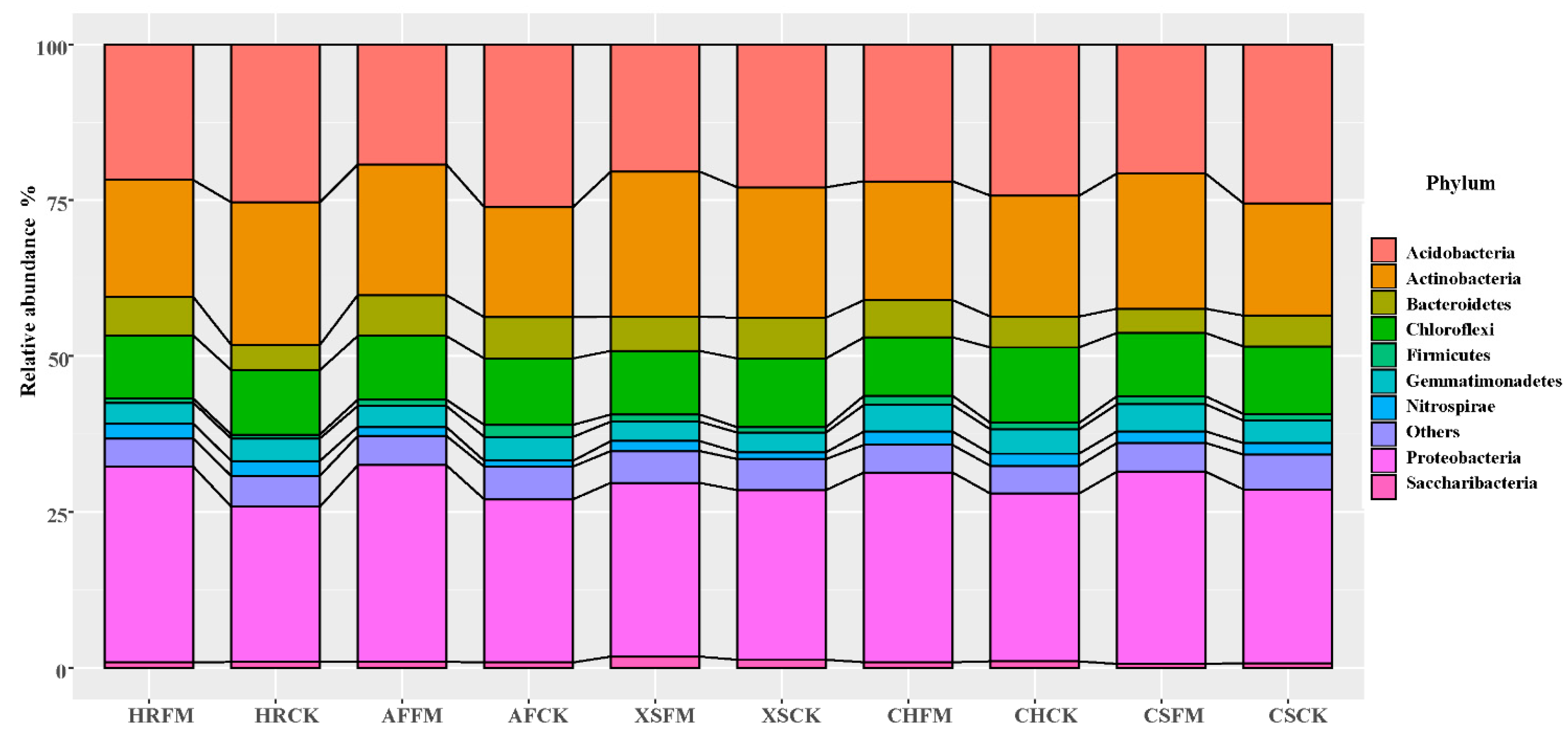
3.2. Soil Alpha Diversity Index and Community Composition under Re-Vegetation Types and Mycorrhizal Fungi Reclamation
3.3. Network Analysis and Functional Prediction of Soil Bacterial Communities
3.4. Relationships among Soil Bacteria, Physicochemical Properties, Re-Vegetation Types and Mycorrhizal Fungi Reclamation
4. Discussion
4.1. Effects of AMF and Re-Vegetation Types on Plant Stoichiometry and Homeostasis
4.2. Effects of AMF and Re-Vegetation Types on Soil Bacterial Community Composition and Function
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yuan, J.; Na, C.; Lei, Q.; Xiong, M.; Guo, J.; Hu, Z. Coal use for power generation in China. Resour. Conserv. Recycl. 2018, 129, 443–453. [Google Scholar] [CrossRef]
- Ramani, R.V. Surface Mining Technology: Progress and Prospects. Procedia Eng. 2012, 46, 9–21. [Google Scholar] [CrossRef]
- Wang, J.; Wang, P.; Qin, Q.; Wang, H. The effects of land subsidence and rehabilitation on soil hydraulic properties in a mining area in the Loess Plateau of China. Catena 2017, 159, 51–59. [Google Scholar] [CrossRef]
- Lechner, A.M.; Baumgartl, T.; Matthew, P.; Glenn, V. The impact of underground longwall mining on prime agricultural land: A review and research agenda. Land. Degrad. Dev. 2016, 27, 1650–1663. [Google Scholar] [CrossRef]
- Liu, X.; Bai, Z.; Zhou, W.; Cao, Y.; Zhang, G. Changes in soil properties in the soil profile after mining and reclamation in an opencast coal mine on the Loess Plateau, China. Ecol. Eng. 2017, 98, 228–239. [Google Scholar] [CrossRef]
- Vidal-Macua, J.J.; Nicolau, J.M.; Vicente, E.; Moreno-de las Heras, M. Assessing vegetation recovery in reclaimed opencast mines of the Teruel coalfield (Spain) using Landsat time series and boosted regression trees. Sci. Total Environ. 2020, 717, 137250. [Google Scholar] [CrossRef]
- Wang, S.; Bi, Y.; Christie, P. Effects of extracellular metabolites from a dark septate endophyte at different growth stages on maize growth, root structure and root exudates. Rhizosphere 2023, 25, 100657. [Google Scholar] [CrossRef]
- Bi, Y.; Wang, X.; Cai, Y.; Christie, P. Arbuscular mycorrhizal colonization increases plant above-belowground feedback in a northwest Chinese coal mining-degraded soil by increasing photosynthetic carbon assimilation and allocation to maize. Environ. Sci. Pollut. R 2022, 29, 72612–72627. [Google Scholar] [CrossRef]
- Bohrer, S.L.; Limb, R.F.; Daigh, A.L.; Volk, J.M.; Wick, A.F. Fine and coarse-scale patterns of vegetation diversity on reclaimed surface mine-land over a 40-year chronosequence. Environ. Manag. 2017, 59, 431–439. [Google Scholar] [CrossRef]
- Jia, X.; Zhao, C.; Wang, Y.; Zhu, Y.; Shao, M.A. Traditional dry soil layer index method overestimates soil desiccation severity following conversion of cropland into forest and grassland on China’s Loess Plateau. Agric. Ecosyst. Environ. 2019, 291, 106794. [Google Scholar] [CrossRef]
- Tripathi, N.; Singh, R.S.; Hills, C.D. Soil carbon development in rejuvenated Indian coal mine spoil. Ecol. Eng. 2016, 90, 482–490. [Google Scholar] [CrossRef]
- Siles, J.A.; Margesin, R. Insights into microbial communities mediating the bioremediation of hydrocarbon-contaminated soil from an Alpine former military site. Appl. Microbiol. Biot. 2018, 102, 4409–4421. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Martinez, M.; Preece, C.; Corbera, J.; Cano, O.; Garcia-Porta, J.; Sardans, J.; Janssens, I.A.; Sabater, F.; Penuelas, J. Bryophyte C:N:P stoichiometry, biogeochemical niches and elementome plasticity driven by environment and coexistence. Ecol. Lett. 2021, 24, 1375–1386. [Google Scholar] [CrossRef]
- Mueller, M.; Oelmann, Y.; Schickhoff, U.; Boehner, J.; Scholten, T. Himalayan treeline soil and foliar C:N:P stoichiometry indicate nutrient shortage with elevation. Geoderma 2017, 291, 21–32. [Google Scholar] [CrossRef]
- Liu, G.; Ye, X.; Huang, Z.; Dong, M.; Cornelissen, J.H.C. Leaf and root nutrient concentrations and stoichiometry along aridity and soil fertility gradients. J. Veg. Sci. 2019, 30, 291–300. [Google Scholar] [CrossRef]
- Jansa, J.; Forczek, S.T.; Rozmos, M.; Puschel, D.; Bukovska, P.; Hrselova, H. Arbuscular mycorrhiza and soil organic nitrogen: Network of players and interactions. Chem. Biol. Technol. Agric. 2019, 6, 10. [Google Scholar] [CrossRef]
- Meng, L.B.; Zhang, A.Y.; Wang, F.; Han, X.G.; Wang, D.J.; Li, S.M. Arbuscular mycorrhizal fungi and rhizobium facilitate nitrogen uptake and transfer in soybean/maize intercropping system. Front. Plant Sci. 2015, 6, 339. [Google Scholar] [CrossRef]
- Mooshammer, M.; Wanek, W.; Zechmeister-Boltenstern, S.; Richter, A. Stoichiometric imbalances between terrestrial decomposer communities and their resources: Mechanisms and implications of microbial adaptations to their resources. Front. Microbiol. 2014, 5, 22. [Google Scholar] [CrossRef]
- Bhardwaj, Y.; Sharma, M.P.; Pandey, J.; Dubey, S.K. Variations in microbial community in a tropical dry deciduous forest across the season and topographical gradient assessed through signature fatty acid biomarkers. Ecol. Res. 2020, 35, 139–153. [Google Scholar] [CrossRef]
- Liu, L.; Ma, L.; Zhu, M.; Liu, B.; Liu, X.; Shi, Y. Rhizosphere microbial community assembly and association networks strongly differ based on vegetation type at a local environment scale. Front. Microbiol. 2023, 14, 1129471. [Google Scholar] [CrossRef]
- Zhong, Z.; Wang, X.; Zhang, X.; Zhang, W.; Xu, Y.; Ren, C.; Han, X.; Yang, G. Edaphic factors but not plant characteristics mainly alter soil microbial properties along a restoration chronosequence of Pinus tabulaeformis stands on Mt. Ziwuling, China. For. Ecol. Manag. 2019, 453, 117625. [Google Scholar] [CrossRef]
- Rodriguez-Caballero, G.; Caravaca, F.; Fernandez-Gonzalez, A.J.; Alguacil, M.M.; Fernandez-Lopez, M.; Roldan, A. Arbuscular mycorrhizal fungi inoculation mediated changes in rhizosphere bacterial community structure while promoting revegetation in a semiarid ecosystem. Sci. Total Environ. 2017, 584, 838–848. [Google Scholar] [CrossRef] [PubMed]
- De Gruyter, J.; Weedon, J.T.; Elst, E.M.; Geisen, S.; van der Heijden, M.G.A.; Verbruggen, E. Arbuscular mycorrhizal inoculation and plant response strongly shape bacterial and eukaryotic soil community trajectories. Soil Biol. Biochem. 2022, 165, 108524. [Google Scholar] [CrossRef]
- Zhao, F.Z.; Ren, C.J.; Han, X.H.; Yang, G.H.; Wang, J.; Doughty, R. Changes of soil microbial and enzyme activities are linked to soil C, N and P stoichiometry in afforested ecosystems. For. Ecol. Manag. 2018, 427, 289–295. [Google Scholar] [CrossRef]
- Zhou, Z.; Wang, C.; Jiang, L.; Luo, Y. Trends in soil microbial communities during secondary succession. Soil Biol. Biochem. 2017, 115, 92–99. [Google Scholar] [CrossRef]
- Wang, Z.G.; Bi, Y.L.; Jiang, B.; Zhakypbek, Y.; Peng, S.P.; Liu, W.W.; Liu, H. Arbuscular mycorrhizal fungi enhance soil carbon sequestration in the coalfields, northwest China. Sci. Rep. 2016, 6, 34336. [Google Scholar] [CrossRef]
- Bi, Y.L.; Guo, Y.; Christie, P. Mining subsidence area reconstruction with N-2-fixing plants promotes arbuscular mycorrhizal fungal biodiversity and microbial biomass C:N:P stoichiometry of cyanobacterial biocrusts. For. Ecol. Manag. 2021, 503, 119763. [Google Scholar] [CrossRef]
- Zeng, Q.; Jia, P.; Wang, Y.; Wang, H.; Li, C.; An, S. The local environment regulates biogeographic patterns of soil fungal communities on the Loess Plateau. Catena 2019, 183, 104220. [Google Scholar] [CrossRef]
- Kumar, S.; Singh, A.K.; Ghosh, P. Distribution of soil organic carbon and glomalin related soil protein in reclaimed coal mine-land chronosequence under tropical condition. Sci. Total Environ. 2018, 625, 1341–1350. [Google Scholar] [CrossRef]
- Kabir, Z.; O’halloran, I.; Fyles, J.; Hamel, C. Seasonal changes of arbuscular mycorrhizal fungi as affected by tillage practices and fertilization: Hyphal density and mycorrhizal root colonization. Plant Soil 1997, 192, 285–293. [Google Scholar] [CrossRef]
- Mori, H.; Maruyama, F.; Kato, H.; Toyoda, A.; Dozono, A.; Ohtsubo, Y.; Nagata, Y.; Fujiyama, A.; Tsuda, M.; Kurokawa, K. Design and experimental application of a novel non-degenerate universal primer set that amplifies prokaryotic 16S rRNA genes with a low possibility to amplify eukaryotic rRNA genes. DNA Res. 2014, 21, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Hood, J.M.; Sterner, R.W. Diet Mixing: Do Animals Integrate Growth or Resources across Temporal Heterogeneity? Am. Nat. 2010, 176, 651–663. [Google Scholar] [CrossRef] [PubMed]
- Persson, J.; Fink, P.; Goto, A.; Hood, J.M.; Jonas, J.; Kato, S.J.O. To be or not to be what you eat: Regulation of stoichiometric homeostasis among autotrophs and heterotrophs. Oikos 2010, 119, 741–751. [Google Scholar] [CrossRef]
- Yu, M.F.; Tao, Y.X.; Liu, W.Z.; Xing, W.; Liu, G.H.; Wang, L.; Ma, L. C, N, and P stoichiometry and their interaction with different plant communities and soils in subtropical riparian wetlands. Environ. Sci. Pollut. Res. 2020, 27, 1024–1034. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Bi, Y.L.; Du, S.Z.; Wang, Y.; Guo, C.; Christie, P. Response of ecological stoichiometry and stoichiometric homeostasis in the plant-litter-soil system to re-vegetation type in arid mining subsidence areas. J. Arid. Environ. 2021, 184, 104298. [Google Scholar] [CrossRef]
- Lan, Z.L.; Zhang, S.L.; Xie, L.C.; Li, X.; Sial, T.A.; Shar, A.G.; Fan, J.L.; Zhang, J.G.; Dong, Q.; Fu, G.J. Effects of Artemisia ordosica on fine-scale spatial distribution of soil C, N and P and physical-chemical properties in the Mu Us Desert, China. J. Soil Sediments 2022, 22, 172–184. [Google Scholar] [CrossRef]
- Lasota, J.; Blonska, E. C:N:P stoichiometry as an indicator of Histosol drainage in lowland and mountain forest ecosystems. For. Ecosyst. 2021, 8, 39. [Google Scholar] [CrossRef]
- Elser, J.J.; Fagan, W.F.; Kerkhoff, A.J.; Swenson, N.G.; Enquist, B.J. Biological stoichiometry of plant production: Metabolism, scaling and ecological response to global change. New. Phytol. 2010, 186, 593–608. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, B.R.; An, S.S. Ecological stoichiometry in leaves, roots, litters and soil among different plant communities in a desertified region of Northern China. Catena 2018, 166, 328–338. [Google Scholar] [CrossRef]
- Liao, J.J.; Dou, Y.X.; An, S.S. Plant community productivity is associated with multiple ecological stoichiometry in restoration grasslands. Ecol. Eng. 2023, 187, 106845. [Google Scholar] [CrossRef]
- Su, Z.; Su, B.; Mao, S.; Shangguan, Z. Leaf C:N:P stoichiometric homeostasis of a Robinia pseudoacacia plantation on the Loess Plateau. J. For. Res. 2022, 34, 929–937. [Google Scholar] [CrossRef]
- Agren, G.I. The C: N: P stoichiometry of autotrophs—Theory and observations. Ecol. Lett. 2004, 7, 185–191. [Google Scholar] [CrossRef]
- Bai, X.; Wang, B.; An, S.; Zeng, Q.; Zhang, H. Response of forest species to C:N:P in the plant-litter-soil system and stoichiometric homeostasis of plant tissues during afforestation on the Loess Plateau, China. Catena 2019, 183, 104186. [Google Scholar] [CrossRef]
- Mariotte, P.; Canarini, A.; Dijkstra, F.A. Stoichiometric N:P flexibility and mycorrhizal symbiosis favour plant resistance against drought. J. Ecol. 2017, 105, 958–967. [Google Scholar] [CrossRef]
- Gusewell, S. N: P ratios in terrestrial plants: Variation and functional significance. New. Phytol. 2004, 164, 243–266. [Google Scholar] [CrossRef] [PubMed]
- Ci, H.; Guo, C.; Tuo, B.; Zheng, L.-T.; Xu, M.-S.; Sai, B.-L.; Yang, B.-Y.; Yang, Y.-C.; You, W.-H.; Yan, E.-R.; et al. Tree species with conservative foliar nutrient status and strong phosphorus homeostasis are regionally abundant in subtropical forests. J. Ecol. 2022, 110, 1497–1507. [Google Scholar] [CrossRef]
- Wang, X.; Wu, D.; Li, S.; Chen, T.; Chen, R.; Yin, L.; Yao, X.; Chen, G. Effects of C:N imbalance on soil microbial physiology in subtropical tree plantations associated with ectomycorrhizal and arbuscular mycorrhizal fungi. Geoderma 2022, 422, 115932. [Google Scholar] [CrossRef]
- Chaudhary, V.B.; Rua, M.A.; Antoninka, A.; Bever, J.D.; Cannon, J.; Craig, A.; Duchicela, J.; Frame, A.; Gardes, M.; Gehring, C.; et al. MycoDB, a global database of plant response to mycorrhizal fungi. Sci. Data 2016, 3, 160028. [Google Scholar] [CrossRef]
- Zeng, Q.C.; Mei, T.Y.; Wang, M.X.; Tan, W.F. Intensive citrus plantations suppress the microbial profiles of the beta-glucosidase gene. Agric. Ecosyst. Environ. 2022, 323, 107687. [Google Scholar] [CrossRef]
- Deng, Z.Y.; Wang, Y.C.; Xiao, C.C.; Zhang, D.X.; Feng, G.; Long, W.X. Effects of Plant Fine Root Functional Traits and Soil Nutrients on the Diversity of Rhizosphere Microbial Communities in Tropical Cloud Forests in a Dry Season. Forests 2022, 13, 421. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, G.; Xue, S.; Wang, G. Soil bacterial community dynamics reflect changes in plant community and soil properties during the secondary succession of abandoned farmland in the Loess Plateau. Soil Biol. Biochem. 2016, 97, 40–49. [Google Scholar] [CrossRef]
- Koyama, A.; Wallenstein, M.D.; Simpson, R.T.; Moore, J.C. Soil bacterial community composition altered by increased nutrient availability in Arctic tundra soils. Front. Microbiol. 2014, 5, 516. [Google Scholar] [CrossRef]
- Fierer, N. Embracing the unknown: Disentangling the complexities of the soil microbiome. Nat. Rev. Microbiol. 2017, 15, 579–590. [Google Scholar] [CrossRef] [PubMed]
- Gui, H.; Purahong, W.; Hyde, K.D.; Xu, J.; Mortimer, P.E. The Arbuscular Mycorrhizal Fungus Funneliformis mosseae Alters Bacterial Communities in Subtropical Forest Soils during Litter Decomposition. Front. Microbiol. 2017, 8, 1120. [Google Scholar] [CrossRef] [PubMed]
- Nuccio, E.E.; Hodge, A.; Pett-Ridge, J.; Herman, D.J.; Weber, P.K.; Firestone, M.K. An arbuscular mycorrhizal fungus significantly modifies the soil bacterial community and nitrogen cycling during litter decomposition. Environ. Microbiol. 2013, 15, 1870–1881. [Google Scholar] [CrossRef]
- Ren, C.; Zhao, F.; Kang, D.; Yang, G.; Han, X.; Tong, X.; Feng, Y.; Ren, G. Linkages of C:N:P stoichiometry and bacterial community in soil following afforestation of former farmland. For. Ecol. Manag. 2016, 376, 59–66. [Google Scholar] [CrossRef]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef] [PubMed]
- Hicks, L.C. Drying-rewetting of permanent pasture and agricultural soils induces a shift towards microbial use of more C-rich organic matter. Soil Biol. Biochem. 2023, 178, 108928. [Google Scholar] [CrossRef]
- Fan, H.; Wu, J.; Liu, W.; Yuan, Y.; Hu, L.; Cai, Q. Linkages of plant and soil C:N:P stoichiometry and their relationships to forest growth in subtropical plantations. Plant Soil 2015, 392, 127–138. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C:N | C:P | N:P | ||||
|---|---|---|---|---|---|---|
| FM | CK | FM | CK | FM | CK | |
| AF | 12.01 ± 0.29 f | 12.73 ± 0.09 f | 157.93 ± 4.01 c | 159.66 ± 5.44 c | 13.15 ± 0.12 a | 12.54 ± 0.28 ab |
| HR | 13.38 ± 0.40 ef | 14.50 ± 0.45 e | 160.73 ± 3.64 c | 160.71 ± 5.04 c | 12.02 ± 0.43 b | 11.10 ± 0.38 c |
| XS | 18.65 ± 1.11 cd | 21.96 ± 1.27 b | 178.94 ± 4.99 b | 185.03 ± 7.34 b | 9.60 ± 0.35 e | 8.44 ± 0.38 f |
| CH | 20.02 ± 1.06 c | 24.88 ± 0.68 a | 213.96 ± 13.28 a | 215.28 ± 18.27 a | 10.69 ± 0.21 cd | 8.70 ± 0.95 f |
| CS | 17.80 ± 0.95 d | 19.49 ± 0.52 c | 179.51 ± 4.60 b | 188.24 ± 1.62 b | 10.10 ± 0.28 de | 9.66 ± 0.29 e |
| P(M) *, P(V) NS, P(V*M) * | P(M)NS, P(V) NS, P(F*V) * | P(M) NS, P(V) NS, P(M*V) NS | ||||
| C:N | C:P | N:P | ||||
|---|---|---|---|---|---|---|
| FM | CK | FM | CK | FM | CK | |
| AF | 18.73 ± 0.56 f | 19.88 ± 0.56 ef | 274.88 ± 10.46 ab | 256.83 ± 12.16 bc | 14.68 ± 0.31 a | 12.98 ± 0.52 b |
| HR | 24.03 ± 0.72 de | 26.78 ± 2.73 cd | 270.50 ± 8.34 ab | 285.33 ± 27.22 a | 11.26 ± 0.14 c | 10.67 ± 0.12 d |
| XS | 31.56 ± 1.74 bc | 33.20 ± 2.58 b | 232.43 ± 11.06 d | 238.54 ± 12.31 cd | 7.37 ± 0.41 e | 6.87 ± 0.72 e |
| CH | 66.26 ± 5.84 a | 68.66 ± 5.22 a | 277.15 ± 9.13 ab | 271.62 ± 10.89 ab | 4.20 ± 0.25 f | 3.99 ± 0.13 f |
| CS | 65.15 ± 1.06 a | 69.90 ± 2.66 a | 281.08 ± 6.97 ab | 292.62 ± 7.65 a | 4.31 ± 0.08 f | 4.16 ± 0.13 f |
| P(M) *, P(V) NS, P(M*V) * | P(F)NS, P(V) NS, P(M*V) * | P(M) NS, P(V) NS, P(M*V) NS | ||||
| Shannon | Simpson | ACE | ||||
|---|---|---|---|---|---|---|
| FM | CK | FM | CK | FM | CK | |
| AF | 1.95 ± 0.02 adc | 1.93 ± 0.04 bc | 0.202 ± 0.013 ab | 0.193 ± 0.002 ab | 35.61 ± 5.19 a | 35.43 ± 2.82 a |
| HR | 1.99 ± 0.02 a | 1.94 ± 0.01 abc | 0.197 ± 0.002 ab | 0.186 ± 0.004 b | 38.49 ± 4.05 a | 34.19 ± 1.05 a |
| XS | 1.98 ± 0.03 ab | 1.96 ± 0.00 ab | 0.191 ± 0.007 b | 0.189 ± 0.002 b | 37.08 ± 2.00 a | 34.63 ± 0.77 a |
| CH | 1.95 ± 0.01 adc | 1.94 ± 0.03 abc | 0.198 ± 0.004 ab | 0.197 ± 0.008 ab | 35.77 ± 1.01 a | 34.59 ± 2.09 a |
| CS | 1.96 ± 0.02 ab | 1.90 ± 0.07 c | 0.210 ± 0.022 a | 0.197 ± 0.003 ab | 36.72 ± 0.73 a | 35.73 ± 3.54 a |
| P(M) *, P(V) NS, P(M*V) * | P(M)NS, P(V) NS, P(M*V) * | P(M) NS, P(V) NS, P(M*V) NS | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, L.; Bi, Y.; Wang, D. Mycorrhizal Fungi Reclamation Promotes Stoichiometric Homeostasis of Re-Vegetation Types and Affects Soil Bacterial Function in Mining Subsidence of Northern Loess Plateau. Forests 2023, 14, 1720. https://doi.org/10.3390/f14091720
Xiao L, Bi Y, Wang D. Mycorrhizal Fungi Reclamation Promotes Stoichiometric Homeostasis of Re-Vegetation Types and Affects Soil Bacterial Function in Mining Subsidence of Northern Loess Plateau. Forests. 2023; 14(9):1720. https://doi.org/10.3390/f14091720
Chicago/Turabian StyleXiao, Li, Yinli Bi, and Dongdong Wang. 2023. "Mycorrhizal Fungi Reclamation Promotes Stoichiometric Homeostasis of Re-Vegetation Types and Affects Soil Bacterial Function in Mining Subsidence of Northern Loess Plateau" Forests 14, no. 9: 1720. https://doi.org/10.3390/f14091720
APA StyleXiao, L., Bi, Y., & Wang, D. (2023). Mycorrhizal Fungi Reclamation Promotes Stoichiometric Homeostasis of Re-Vegetation Types and Affects Soil Bacterial Function in Mining Subsidence of Northern Loess Plateau. Forests, 14(9), 1720. https://doi.org/10.3390/f14091720