
Impact of Climate Change on the Habitat Suitability of Monochamus saltuarius Gebler (Coleoptera; Cerambycidae) and Its Natural Enemies in China

Abstract
:1. Introduction
2. Materials and Methods
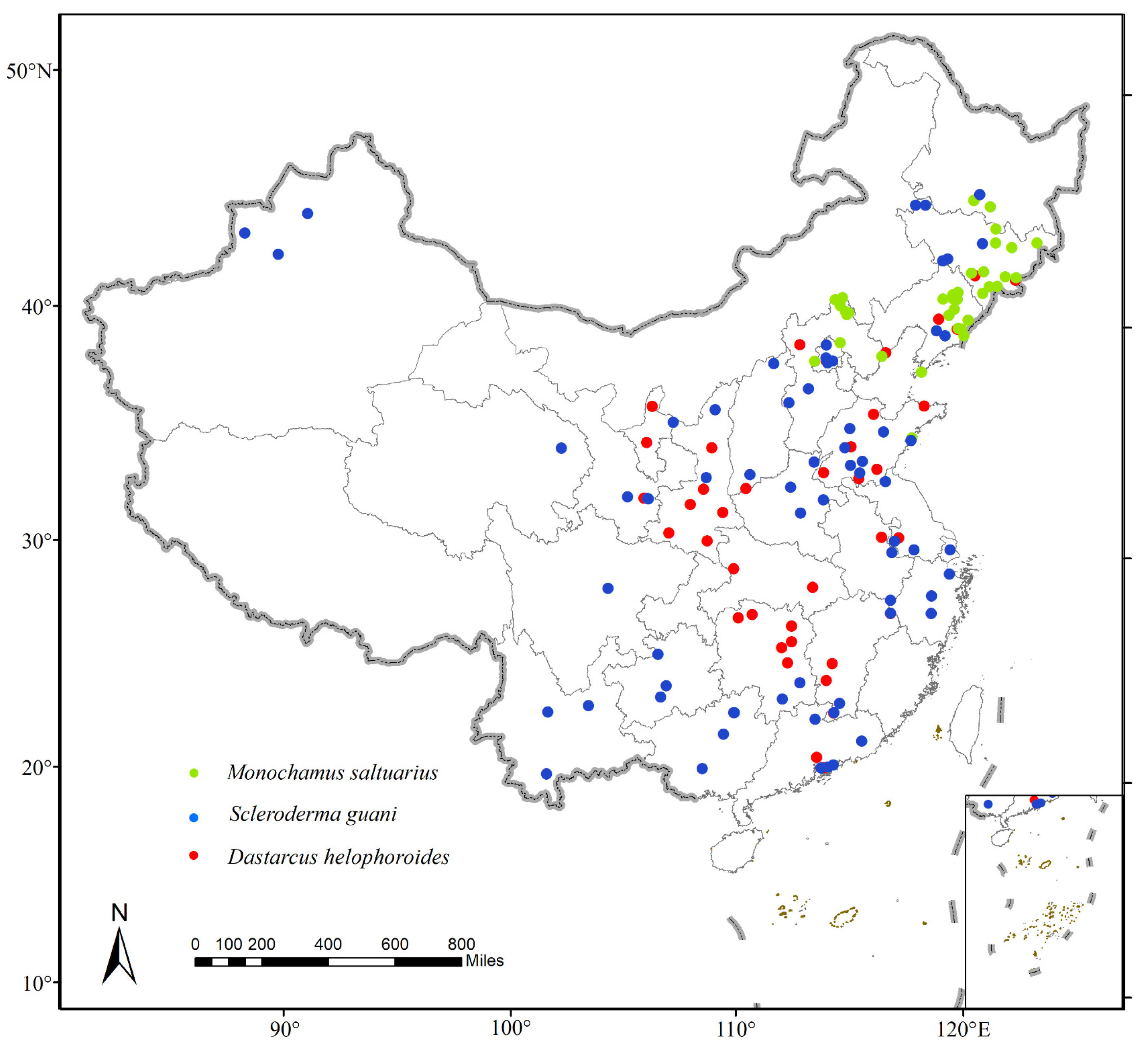
2.1. Species Location Data
2.2. Acquisition and Selection of Bioclimate Variables
2.3. Optimization of Maxent Model Parameters
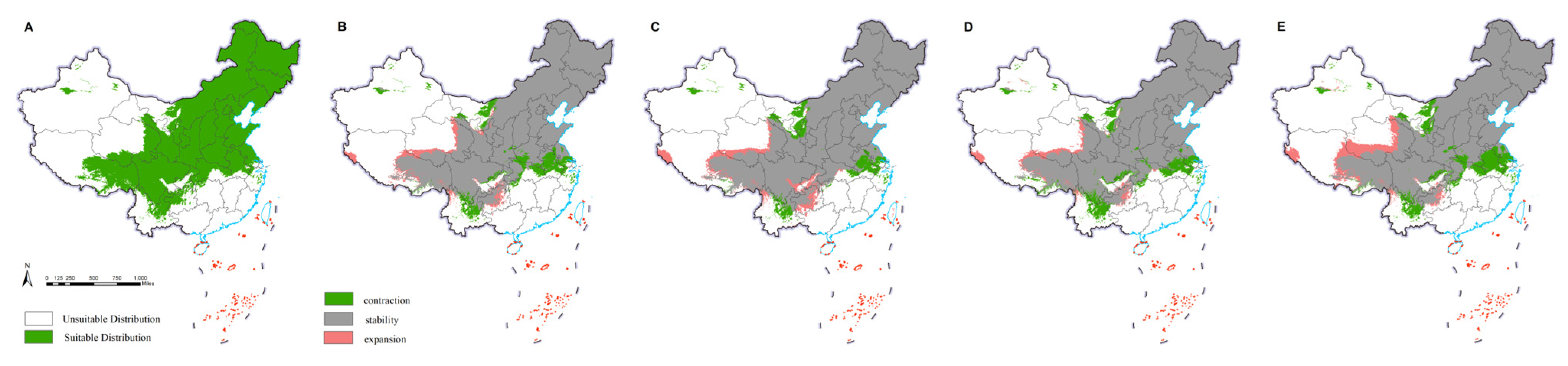
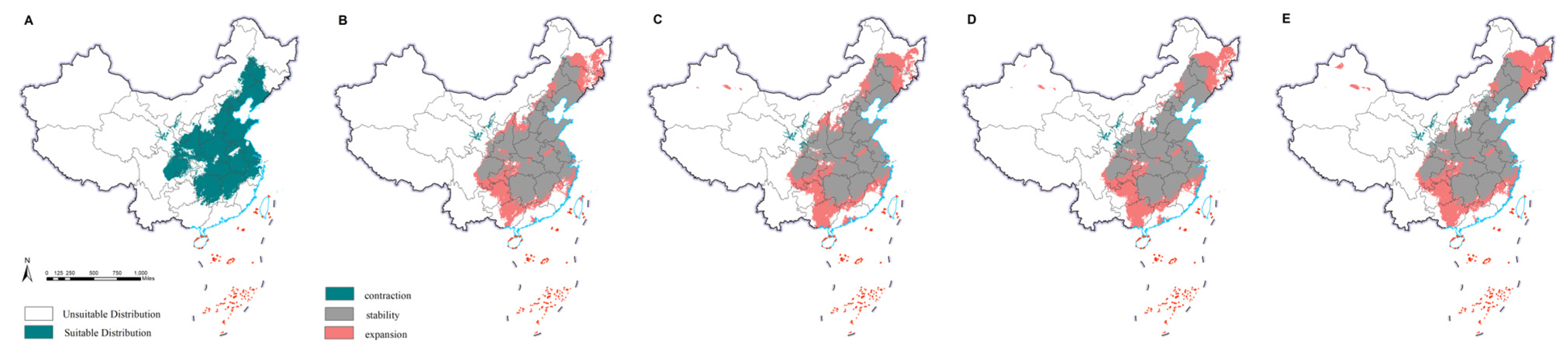
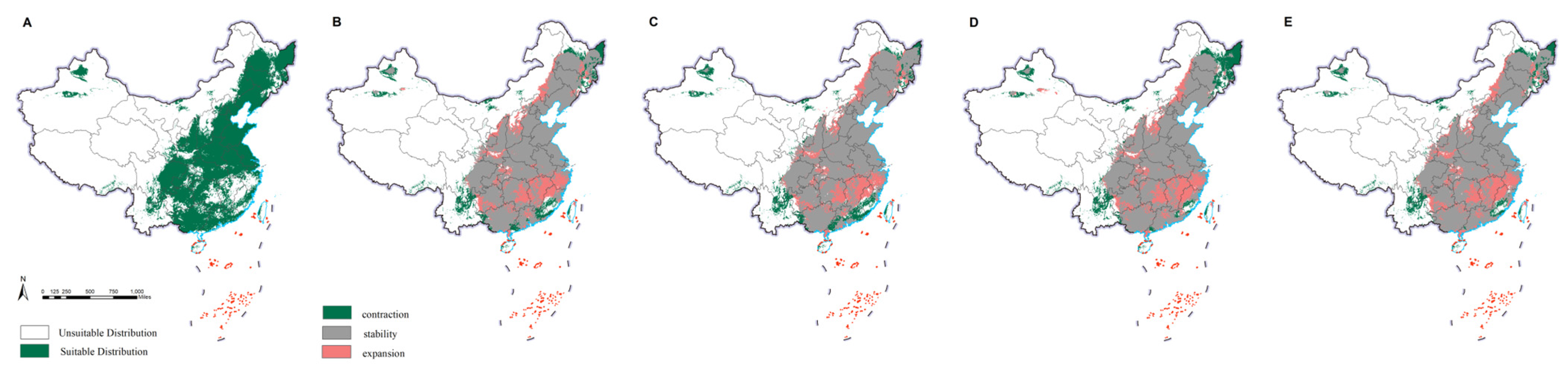
2.4. Changes in the Potential Distributions of M. saltuarius and Natural Enemies
2.5. Calculation of Fitness Overlap between M. saltuarius and Natural Enemies
3. Results
3.1. Model Optimization Results
3.2. Projected Changes in Species Distributions
3.3. Quantitative Analysis of Species Overlap under Current Climate Change
4. Discussion
4.1. Model Prediction Reliability
4.2. Ecological and Environmental Factors Influencing the Distribution of M. saltuarius and Its Natural Enemies
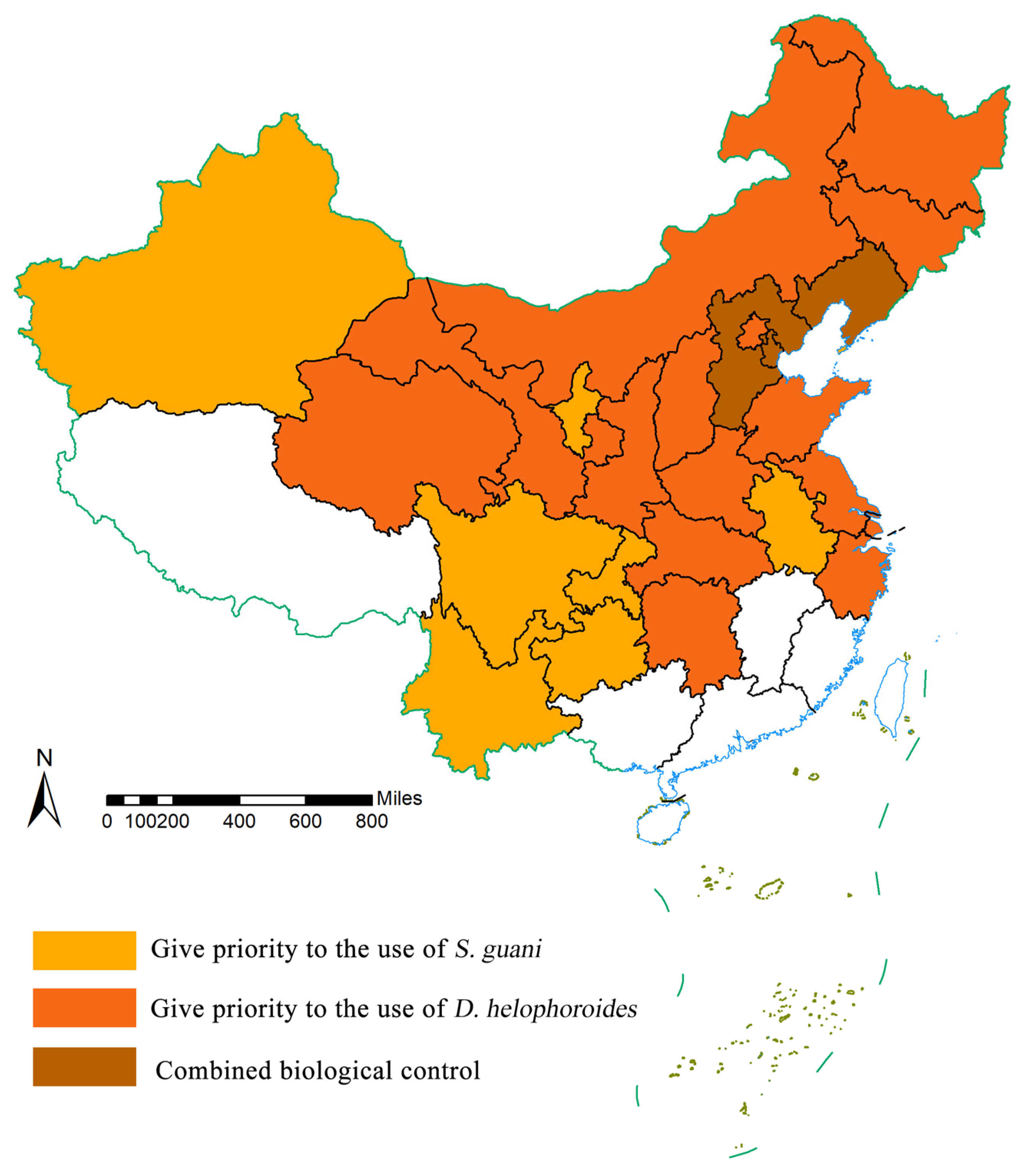
4.3. Future Control Strategy for M. saltuarius Based on Model Predictions
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, M.; Li, H.; Sheng, R.C.; Sun, H.; Sun, S.H.; Chen, F.M. The First Record of Monochamus saltuarius (Coleoptera; Cerambycidae) as Vector of Bursaphelenchus xylophilus and Its New Potential Hosts in China. Insects 2020, 11, 636. [Google Scholar] [CrossRef] [PubMed]
- Yusuke, N.; Katsumi, T. Transmission of Bursaphelenchus mucronatus Mamiya et Enda (Nematoda, Aphelenchoididae) from Monochamus saltuarius (Gebler) (Coleoptera, Cerambycidae) to pine branches via vector’s oviposition wounds in comparison to B. xylophilus (Steiner et Buhrer) Nickle carried by M. alternatus Hope. J. For. Res. 2022, 27, 236–243. [Google Scholar] [CrossRef]
- Zhao, S.G.; Chen, G.F.; Qu, H.C.; Xu, S.L.; Lu, Y.H.; Wang, J. Advances in studies on biological and ecological characteristics of Monochamus saltuarius. For. Pest Dis. 2021, 40, 37–43. [Google Scholar]
- Lim, J.; Jung, S.Y.; Lim, J.S.; Jang, J.; Kim, K.M.; Lee, Y.M.; Lee, B.W. A review of host plants of Cerambycidae (Coleoptera: Chrysomeloidea) with new host records for fourteen cerambycids, including the Asian longhorn beetle (Anoplophora glabripennis Motschulsky), in Korea. Korean J. Appl. Entomol. 2014, 53, 111–133. [Google Scholar] [CrossRef]
- Pang, J.L.; Li, J.; Dong, Y.Q. Feeding preference of Monochamus saltuarius Gebler (Coleoptera: Cerambycidae) for Pinus koraiensis, Pinus tabulaeformis and Larix kaempferi. For. Pest Dis. 2020, 39, 19–22. [Google Scholar]
- Jiang, S.W.; Wu, H.; Li, D.B.; Luo, Z.Q.; He, S.; Song, Y.S. Analysis on disaster characteristics of pine wood nematode in Northeast China. For. Pest Dis. 2022, 41, 9–15. [Google Scholar]
- Hanks, L.M. Influence of the larval host plant on reproductive strategies of cerambycid beetles. Annu. Rev. Entomol. 1999, 44, 483–505. [Google Scholar] [CrossRef]
- Lee, H.R.; Lee, S.C.; Lee, D.H.; Choi, W.S.; Jung, C.S.; Jeon, J.H.; Kim, J.E.; Park, I.K. Identification of the Aggregation-sex Pheromone Produced by Male Monochamus saltuarius, a Major Insect Vector of the Pine Wood Nematode. J. Chem. Ecol. 2017, 43, 670–678. [Google Scholar] [CrossRef]
- Ye, J.R.; Wu, X.Q. Research progress of pine wilt disease. For. Pest Dis. 2022, 41, 1–10. [Google Scholar]
- Carnegie, A.J.; Venn, T.; Lawson, S.; Nagel, M.; Wardlaw, T.; Cameron, N.; Last, I. An analysis of pest risk and potential economic impact of pine wilt disease to Pinus plantations in Australia. Aust. For. 2018, 81, 24–36. [Google Scholar] [CrossRef]
- Zheng, Y.N.; Shi, Y.; Li, Y.; Fan, L.C.; Wang, J.; Wang, W.T. Feeding preference of Monochamus saltuarius (Coleoptera: Cerambycidae) adults for four host pine trees. Acta Entomol. Sin. 2021, 64, 1478–1482. [Google Scholar]
- Zou, Y.; Ge, X.Z.; Zhou, Y.T.; Guo, S.W.; Wang, T.; Zong, S.X. Risk of Monochamus saltuarius spreading in China. Chin. J. Appl. Entomol. 2023, 60, 287–297. [Google Scholar]
- Han, J.; Kim, H.K.; Kang, W.J.; Kim, G. Feeding and oviposition preference of the Sakhalin pine sawyer Monochamus saltuarius (Coleoptera: Cerambycidae) for various tree species. Entomol. Res. 2016, 46, 331–336. [Google Scholar] [CrossRef]
- Yusuke, N.; Katsumi, T. Oviposition sites constructed in Pinus densiflora phloem by Monochamus alternatus and Monochamus saltuarius (Coleoptera, Cerambycidae). J. For. Res. 2023, 28, 78–81. [Google Scholar]
- Golec, J.R.; Li, F.; Cao, L.; Wang, X.; Duan, J.J. Mortality factors of Anoplophora glabripennis (Coleoptera: Cerambycidae) infesting Salix and Populus in central, northwest, and northeast China. Biol. Control 2018, 126, 198–208. [Google Scholar] [CrossRef]
- Zheng, Y.N.; Zhang, Y.L.; Shi, Y.; Fan, L.C.; Li, Y. Preliminary study on the control efficiency of Monochamus saltuarius by Sclerodermus spp. Chin. J. Biol. Control 2022, 38, 306–311. [Google Scholar]
- Zheng, Y.N.; Jiang, S.W.; Huang, F.Y.; Yang, Z.Q.; Gao, G.P.; Liu, P.X.; Zhang, Y.L. Dastarcus helophoroides, Natural Enemy of Monochamus saltuarius was Found in Pine Tree Infected by Bursaphelenchus xylophilus in Liaoning, China. Chin. J. Biol. Control 2020, 36, 315–318. [Google Scholar]
- Kocmánková, E.; Trnka, M.; Juroch, J.; Dubrovský, M.; Semerádová, D.; Možný, M.; Žalud, Z. Impact of climate change on the occurrence and activity of harmful organisms. Plant Prot. Sci. 2009, 45, S48–S52. [Google Scholar] [CrossRef]
- Skendžić, S.; Zovko, M.; Živković, I.; Lešić, V.; Lemić, D. The impact of climate change on agricultural insect pests. Insects 2021, 12, 440. [Google Scholar] [CrossRef]
- Seidl, R.; Thom, D.; Kautz, M. Forest disturbances under climate change. Nat. Clim. Chang. 2017, 7, 395–402. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J. Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Franklin, J. Species distribution models in conservation biogeography: Developments and challenges. Divers. Distrib. 2013, 19, 1217–1223. [Google Scholar] [CrossRef]
- Morente-López, J.; Kass, J.M.; Lara-Romero, C. Ecological niche models as hypothesis generators of functional genetic differentiation and potential local adaptation in a Mediterranean alpine ecosystem. bioRxiv 2020. [Google Scholar] [CrossRef]
- Searcy, C.A.; Shaffer, H.B. Do Ecological Niche Models Accurately Identify Climatic Determinants of Species Ranges? Am. Nat. 2016, 187, 423–435. [Google Scholar] [CrossRef] [PubMed]
- Elith, J.; Phillips, S.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of maxent for ecologists. Divers. Distrib. 2010, 17, 43–57. [Google Scholar] [CrossRef]
- Phillips, S.; Anderson, R.H.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Araújo, M.; Luoto, M. The importance of biotic interactions for modelling species distributions under climate change. Glob. Ecol. Biogeogr. 2007, 16, 743–753. [Google Scholar] [CrossRef]
- Jiménez-Valverde, A.; Peterson, A.; Soberón, J.; Overton, J.; Aragón, P.; Lobo, J. Use of niche models in invasive species risk assessments. Biol. Invasions 2011, 13, 2785–2797. [Google Scholar] [CrossRef]
- Smeraldo, S.; Bosso, L.; Salinas-Ramos, V.B.; Ancillotto, L.; Sánchez-Cordero, V.; Gazaryan, S.; Russo, D. Generalists yet different: Distributional responses to climate change may vary in opportunistic bat species sharing similar ecological traits. Mammal Rev. 2021, 51, 571–584. [Google Scholar] [CrossRef]
- Kwon, H.S.; Kim, B.J.; Jang, G.S. Modelling the spatial distribution of wildlife animals using presence and absence data. Contemp. Probl. Ecol. 2016, 9, 515–528. [Google Scholar] [CrossRef]
- Wu, T.; Lu, Y.; Fang, Y.; Xin, X.; Li, L.; Li, W.; Liu, X. The Beijing climate center climate system model (BCC-CSM): The main progress from cmip5 to cmip6. Geosci. Model Dev. 2019, 12, 1573–1600. [Google Scholar] [CrossRef]
- Nie, S.; Fu, S.; Cao, W. Comparison of monthly air and land surface temperature extremes simulated using CMIP5 and CMIP6 versions of the Beijing Climate Center climate model. Theor. Appl. Climatol. 2020, 140, 487–502. [Google Scholar] [CrossRef]
- Eyring, V.; Bony, S.; Meehl, G.A.; Senior, C.A.; Stevens, B.; Stouffer, R.J.; Taylor, K.E. Overview of the Coupled Model Intercomparison Project Phase 6 (CMIP6) experimental design and organization. Geosci. Model Dev. 2016, 9, 1937–1958. [Google Scholar] [CrossRef]
- O’Neill, B.C.; Tebaldi, C.; Van Vuuren, D.P. The scenario model intercomparison project (Scenario MIP) for CMIP6. Geosci. Model Dev. 2016, 9, 3461–3482. [Google Scholar] [CrossRef]
- Sillero, N.; Barbosa, A.M. Common mistakes in ecological niche models. Int. J. Geogr. Inf. Sci. 2021, 35, 213–226. [Google Scholar] [CrossRef]
- Merow, C.; Smith, M.; Silander, J. A practical guide to maxent for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Zhu, G.P.; Qiao, H.J. Effect of the Maxent model’s complexity on the prediction of species potential distributions. Biodivers. Sci. 2016, 24, 1189–1196. [Google Scholar] [CrossRef]
- Radosavljevic, A.; Anderson, R.P. Making better Maxent models of species distributions: Complexity, overfitting and evaluation. J. Biogeogr. 2014, 41, 629–643. [Google Scholar] [CrossRef]
- Moreno-Amat, E.; Saura, S.; Nieto-Lugilde, D.; Morueta-Holme, N.; Svenning, J.; García-Amorena, I. Impact of model complexity on cross-temporal transferability in maxent species distribution models: An assessment using paleo botanical data. Ecol. Model. 2015, 312, 308–317. [Google Scholar] [CrossRef]
- Syfert, M.M.; Smith, M.J.; Coomes, D.A. The effects of sampling bias and model complexity on the predictive performance of maxent species distribution models. PLoS ONE 2013, 8, e55158. [Google Scholar] [CrossRef]
- Fithian, W.; Elith, J.; Hastie, T.; Keith, D.A. Bias correction in species distribution models: Pooling survey and collection data for multiple species. Methods Ecol. Evol. 2015, 6, 424–438. [Google Scholar] [CrossRef] [PubMed]
- Warren, D.L.; Seifert, S.N. Ecological niche modeling in Maxent: The importance of model complexity and the performance of model selection criteria. Ecol. Appl. 2011, 21, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Vignali, S.; Barras, A.G.; Arlettaz, R.; Braunisch, V. SDMtune: An R package to tune and evaluate species distribution models. Ecol. Evol. 2020, 10, 11488–11506. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Dang, Y.Q.; Wang, X. Risk Assessment of the Worldwide Expansion and Outbreak of Massicus raddei (Blessig) (Coleoptera: Cerambycidae) Based on Host Plant and Climatic Factors. Insects 2022, 13, 730. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.L. SDM toolbox: A python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. Methods Ecol. Evol. 2014, 5, 694–700. [Google Scholar] [CrossRef]
- Schoener, T. The anoles lizards of Bimini: Resource partitioning in a complex fauna. Ecology 1968, 49, 704–726. [Google Scholar] [CrossRef]
- Warren, D.; Glor, R.; Turelli, M. Environmental niche equivalency versus conservatism: Quantitative approaches to niche evolution. Evolution 2008, 62, 2868–2883. [Google Scholar] [CrossRef] [PubMed]
- Lobo, J.M.; Jiménez-Valverde, A.; Real, R. AUC: A misleading measure of the performance of predictive distribution models. Glob. Ecol. Biogeogr. 2008, 17, 145–151. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Estay, S.; Labra, F.; Sepúlveda, R.; Bacigalupe, L. Evaluating habitat suitability for the establishment of Monochamus spp. through climate-based niche modeling. PLoS ONE 2014, 9, e102592. [Google Scholar] [CrossRef]
- Deng, X.; Xu, D.; Liao, W.; Wang, R.; Zhuo, Z. Predicting the distributions of Scleroderma guani (Hymenoptera: Bethylidae) under climate change in China. Ecol. Evol. 2022, 12, e9410. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Wang, J.; Lei, Y. Predicting Distribution of the Asian Longhorned Beetle, Anoplophora glabripennis (Coleoptera: Cerambycidae) and its natural enemies in China. Insects 2022, 13, 687. [Google Scholar] [CrossRef] [PubMed]
- Shim, J.; Jung, J.; Byeon, D.; Jung, S.; Lee, W. Evaluation of the spatial distribution of Anoplophora glabripennis (motschulsky) (Coleoptera: Cerambycidae) in South Korea combining climate and host plant distribution. J. Asia-Pac. Entomol. 2020, 23, 646–652. [Google Scholar] [CrossRef]
- Pureswaran, D.S.; Roques, A.; Battisti, A. Forest Insects and Climate Change. Curr. For. Rep. 2018, 4, 35–50. [Google Scholar] [CrossRef]
- Dang, Y.Q.; Zhang, Y.; Wang, X.; Xin, B.; Quinn, N.; Duan, J. Retrospective analysis of factors affecting the distribution of an invasive wood-boring insect using native range data: The importance of host plants. J. Pest Sci. 2021, 94, 981–990. [Google Scholar] [CrossRef]
- Wei, J.R.; Yang, Z.Q.; Niu, Y.L.; Zhao, H.B.; Tang, H. Distribution and ecological biology of Dastarcus helophoroides. For. Pest Dis. 2009, 28, 16–18. [Google Scholar]
- Zhou, Y.J.; Lu, C.D.; Shen, H.Y.; Liang, Y.; Chen, Z.H.; Zhong, J.H.; Tan, J.; Wang, X.Y.; Liang, G.H. Adaptation and Physiological Response of Dastarcus helophoroides (Fairmaire) to High Temperature Stress. Chin. J. Biol. Control 2021, 37, 1179–1188. [Google Scholar]
- Yao, D.F.; Yan, J.J.; Xu, C.H. Study on the relationship between temperature and humidity and the growth and development of Scleroderma guani Xiao et Wu. Sci. Silvae Sin. 1983, 19, 85–91. [Google Scholar]
- Tian, S.Z.; Zhang, Z.X. Study on the effective accumulated temperature of Scleroderma guani. J. Shandong For. Sci. Technol. 1983, 2, 33–36. [Google Scholar]
- Gao, X.; Zhao, D.S.; Zheng, D. Regional Differences in Surface Temperature Variation in China from 1961 to 2018. Chin. J. Atmos. Sci. 2023, 47, 995–1006. [Google Scholar]
- Liang, C.; Zhu, G.M.; Zhen, H.L.; Lin, W.; Jason, F.; Yan, P.L. Dynamical Downscaling of Temperature and Precipitation Extremes in China under Current and Future Climates. Atmos.-Ocean 2018, 56, 55–70. Available online: https://www.tandfonline.com/doi/full/10.1080/07055900.2017.1422691 (accessed on 17 September 2023).
- Gao, R.; Liu, L.; Zhao, L.; Cui, S. Potentially Suitable Geographical Area for Monochamus alternatus under Current and Future Climatic Scenarios Based on Optimized MaxEnt Model. Insects 2023, 14, 182. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.R. Epidemic Status of Pine Wilt Disease in China and Its Prevention and Control Techniques and Counter Measures. Sci. Silvae Sin. 2019, 55, 1–10. [Google Scholar]
- David, G.; Giffard, B.; Piou, D.; Roques, A.; Jactel, H. Potential effects of climate warming on the survivorship of adult Monochamus galloprovincialis. Agric. For. Entomol. 2016, 19, 192–199. [Google Scholar] [CrossRef]
- Liu, Q.; Wu, Z.W.; Lin, S.T.; Li, S.; Fang, Z.B. Spatial point pattern analysis of pine wilt disease occurrence and its influence factors. Chin. J. Appl. Ecol. 2022, 33, 2530–2538. [Google Scholar]
- Xu, X.G.; Yu, R.B.; Yang, C.X.; Liu, H.J.; Zhou, Z.L.; Shen, Y.J. Prediction of invasion risk of pine wilt disease based on GIS spatial technology and MaxEnt model in western Sichuan. J. Beijing For. Univ. 2023, 45, 104–115. [Google Scholar]
- Zheng, Y.N.; Wang, J.; Wang, W.T.; Li, Y.; Fan, L.C.; Shi, Y.; Zhang, Y.L. Parasitic Effect of Dastarcus helophoroides of Monochamus alternatus Biotype on Monochamus saltuarius. Chin. J. Biol. Control 2022, 38, 587–594. [Google Scholar]
- Cui, Y.S.; Liu, Y.P.; Song, Y.S.; Xu, F.Y.; Xu, K.Q. Control of Monochamus alternatus with natural enemies. For. Pest Dis. 2011, 30, 31–33. [Google Scholar]
- Guo, Y.L.; Zhao, F.Z.; Qiao, H.J.; Wang, R.; Wei, H.Y.; Wang, L.S.; Li, X. Challenges and Development Trend of Species Distribution Model. Adv. Earth Sci. 2020, 35, 1292–1305. [Google Scholar]
- Mi, C.; Huettmann, F.; Li, X.; Jiang, Z.; Du, W.; Sun, B. Effects of climate and human activity on the current distribution of amphibians in China. Conserv. Biol. J. Soc. Conserv. Biol. 2022, 36, e13964. [Google Scholar] [CrossRef]
- Carvajal Acosta, A.N.; Mooney, K. Effects of geographic variation in host plant resources for a specialist herbivore’s contemporary and future distribution. Ecosphere 2021, 12, e03822. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Name | Variable Codes | Environment Variable Meaning (Unit) | Permutation Importance (%) |
|---|---|---|---|
| Monochamus saltuarius | bio1 | Annual mean temperature (°C) | 45.2 |
| bio18 | Precipitation of the warmest quarter (mm) | 23.0 | |
| bio8 | Mean temperature of the wettest quarter (°C) | 20.3 | |
| bio19 | Precipitation of the coldest quarter (mm) | 9.9 | |
| bio7 | Temperature annual range (bio5-bio6) (°C) | 1.6 | |
| Dastarcus helophoroides | bio4 | Temperature seasonality (standard deviation × 100) | 54.5 |
| bio1 | Annual mean temperature (°C) | 31.9 | |
| bio16 | Precipitation of the wettest quarter (mm) | 4.7 | |
| bio19 | Precipitation of the coldest quarter (mm) | 4.6 | |
| bio18 | Precipitation of the warmest quarter (mm) | 2.4 | |
| bio2 | Mean diurnal range (°C) | 1.3 | |
| bio5 | Maximum temperature of the warmest month (°C) | 0.6 | |
| Scleroderma guani | bio4 | Temperature seasonality (standard deviation × 100) | 56.9 |
| bio10 | Mean temperature of the warmest quarter (°C) | 16.2 | |
| bio2 | Mean diurnal range (°C) | 8.2 | |
| bio18 | Precipitation of the warmest quarter (mm) | 7.2 | |
| bio17 | Precipitation of the driest quarter (mm) | 5.3 | |
| bio19 | Precipitation of the coldest quarter (mm) | 3.1 | |
| bio8 | Mean temperature of the wettest quarter (°C) | 2.3 | |
| bio15 | Precipitation seasonality (coefficient of variation) | 0.9 |
| Parameters | Species Name | RM | FC | Train TSS | Train AUC | Test TSS | Test AUC |
|---|---|---|---|---|---|---|---|
| Default | Monochamus saltuarius | 1 | LQPH | 0.9886 | 0.9972 | 0.9826 | 0.9968 |
| Dastarcus helophoroides | 0.9793 | 0.9936 | 0.8544 | 0.9723 | |||
| Scleroderma guani | 0.9090 | 0.9792 | 0.9657 | 0.9893 | |||
| Optimization | Monochamus saltuarius | 0.7 | LQPH | 0.9890 | 0.9974 | 0.9873 | 0.9975 |
| Dastarcus helophoroides | 0.5 | QPH | 0.9834 | 0.9944 | 0.9143 | 0.9835 | |
| Scleroderma guani | 0.5 | LQPHT | 0.9273 | 0.9896 | 0.9873 | 0.9966 |
| Species | SSPs | N | N1 | N2 (%) | N3 (%) | Trend |
|---|---|---|---|---|---|---|
| Monochamus saltuarius | current | 1 | 0.497 | reduction | ||
| SSP1-2.6 | 0.956 | 0.475 | 2.30 | 4.48 | ||
| SSP2-4.5 | 0.987 | 0.490 | 3.06 | 3.69 | ||
| SSP3-7.0 | 0.950 | 0.472 | 2.05 | 4.53 | ||
| SSP5-8.5 | 0.963 | 0.478 | 3.75 | 5.60 | ||
| Dastarcus helophoroides | current | 1 | 0.201 | expansion | ||
| SSP1-2.6 | 1.444 | 0.291 | 9.22 | 0.28 | ||
| SSP2-4.5 | 1.484 | 0.299 | 10.05 | 0.31 | ||
| SSP3-7.0 | 1.462 | 0.295 | 9.75 | 0.44 | ||
| SSP5-8.5 | 1.518 | 0.306 | 10.85 | 0.40 | ||
| Scleroderma guani | current | 1 | 0.313 | expansion | ||
| SSP1-2.6 | 1.067 | 0.334 | 6.13 | 4.04 | ||
| SSP2-4.5 | 1.055 | 0.330 | 5.83 | 4.12 | ||
| SSP3-7.0 | 1.035 | 0.324 | 5.95 | 4.85 | ||
| SSP5-8.5 | 1.077 | 0.337 | 5.99 | 3.58 |
| Province | M. saltuarius + S. guani | M. saltuarius + D. helophoroides |
|---|---|---|
| Anhui | 0.77 | 0.75 |
| Beijing | 0.92 | 0.95 |
| Gansu | 0.82 | 0.86 |
| Hebei | 0.72 | 0.72 |
| Henan | 0.70 | 0.76 |
| Heilongjiang | 0.80 | 0.87 |
| Hubei | 0.68 | 0.72 |
| Hunan | 0.82 | 0.90 |
| Jilin | 0.79 | 0.81 |
| Jiangsu | 0.57 | 0.63 |
| Liaoning | 0.89 | 0.89 |
| Inner Mongolia | 0.80 | 0.86 |
| Ningxia | 0.91 | 0.88 |
| Qinghai | - | 0.87 |
| Shandong | 0.76 | 0.77 |
| Shanxi | 0.64 | 0.67 |
| Shaanxi | 0.67 | 0.75 |
| Shanghai | 0.96 | 0.97 |
| Tianjin | 0.92 | 0.92 |
| Zhejiang | 0.85 | 0.86 |
| Guizhou | 0.50 | - |
| Sichuan | 0.60 | - |
| Xinjiang | 0.75 | - |
| Yunnan | 0.61 | - |
| Chongqing | 0.72 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Zhou, Y.; Xu, T.; Zong, S. Impact of Climate Change on the Habitat Suitability of Monochamus saltuarius Gebler (Coleoptera; Cerambycidae) and Its Natural Enemies in China. Forests 2024, 15, 33. https://doi.org/10.3390/f15010033
Zhang X, Zhou Y, Xu T, Zong S. Impact of Climate Change on the Habitat Suitability of Monochamus saltuarius Gebler (Coleoptera; Cerambycidae) and Its Natural Enemies in China. Forests. 2024; 15(1):33. https://doi.org/10.3390/f15010033
Chicago/Turabian StyleZhang, Xuemei, Yuting Zhou, Tian Xu, and Shixiang Zong. 2024. "Impact of Climate Change on the Habitat Suitability of Monochamus saltuarius Gebler (Coleoptera; Cerambycidae) and Its Natural Enemies in China" Forests 15, no. 1: 33. https://doi.org/10.3390/f15010033
APA StyleZhang, X., Zhou, Y., Xu, T., & Zong, S. (2024). Impact of Climate Change on the Habitat Suitability of Monochamus saltuarius Gebler (Coleoptera; Cerambycidae) and Its Natural Enemies in China. Forests, 15(1), 33. https://doi.org/10.3390/f15010033