
Nitrogen Deposition Modulates Litter Decomposition and Enhances Water Retention in Subtropical Forests


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study
2.2. Litter Collection and Simulated N Deposition Experiment
2.3. Sample Measurements
2.4. Statistical Analyses
3. Results
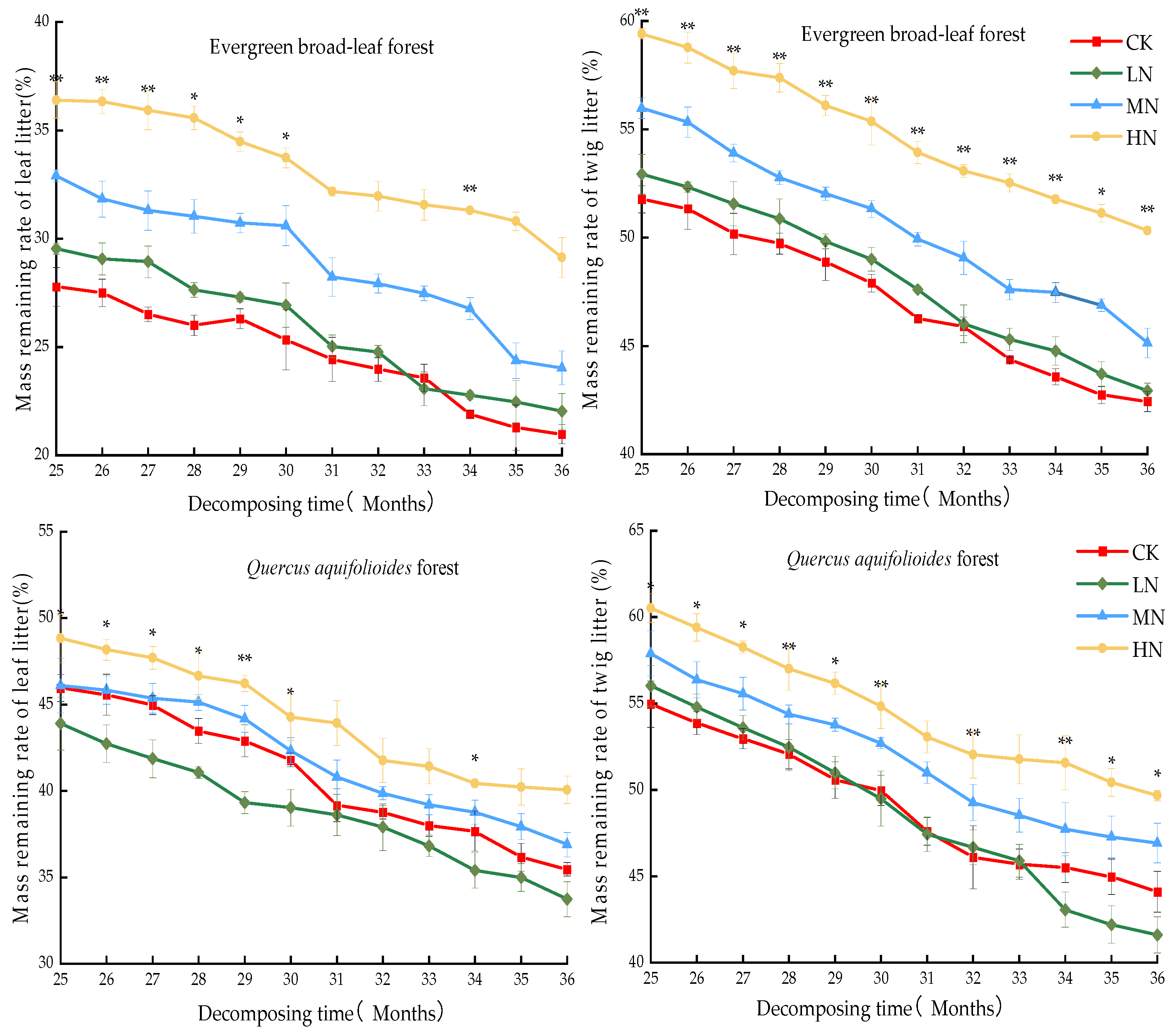
3.1. Decomposition Rate and Loss of Leaf Litter Mass
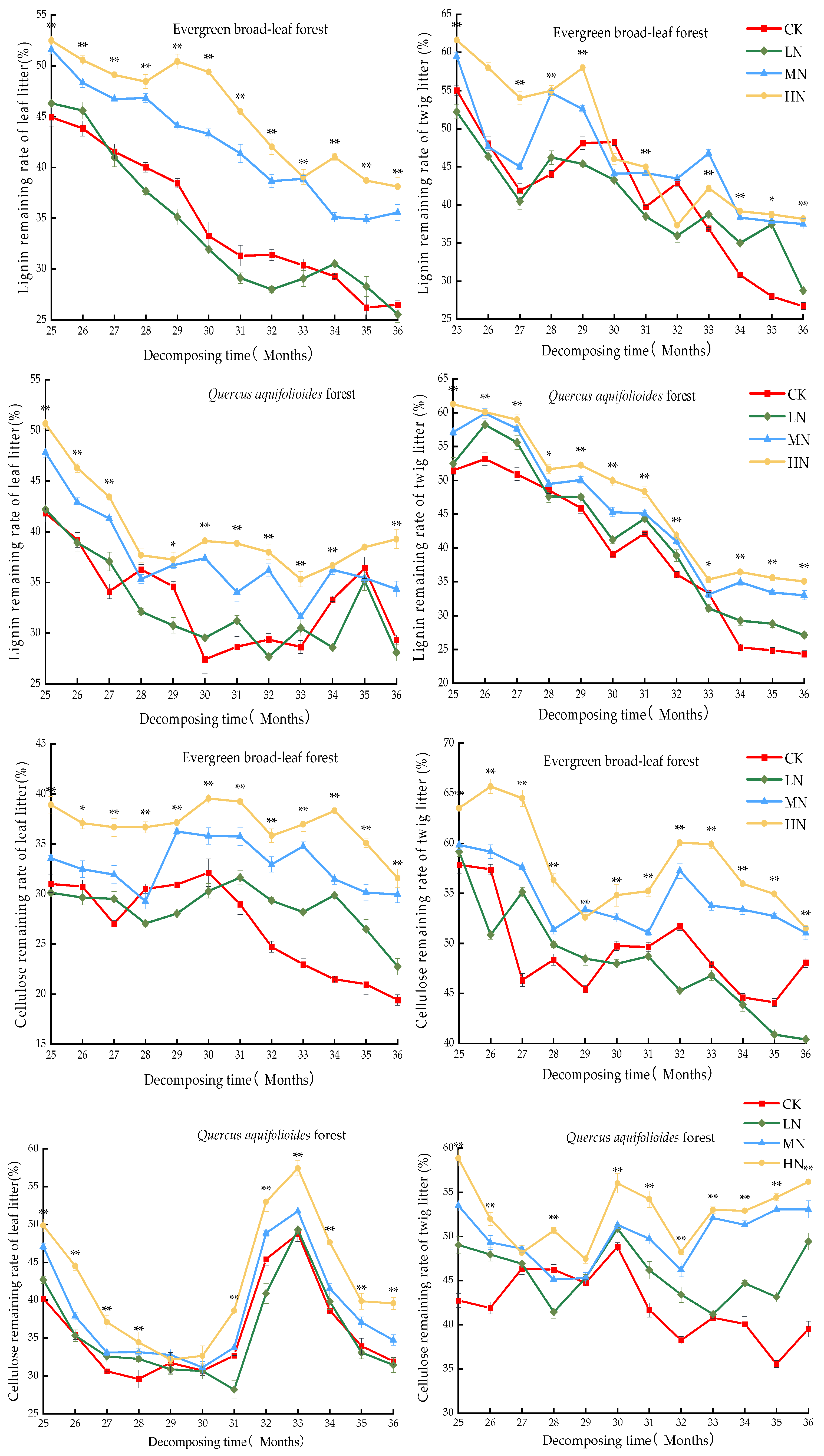
3.2. Decomposition Rate of Lignin and Cellulose in Litter
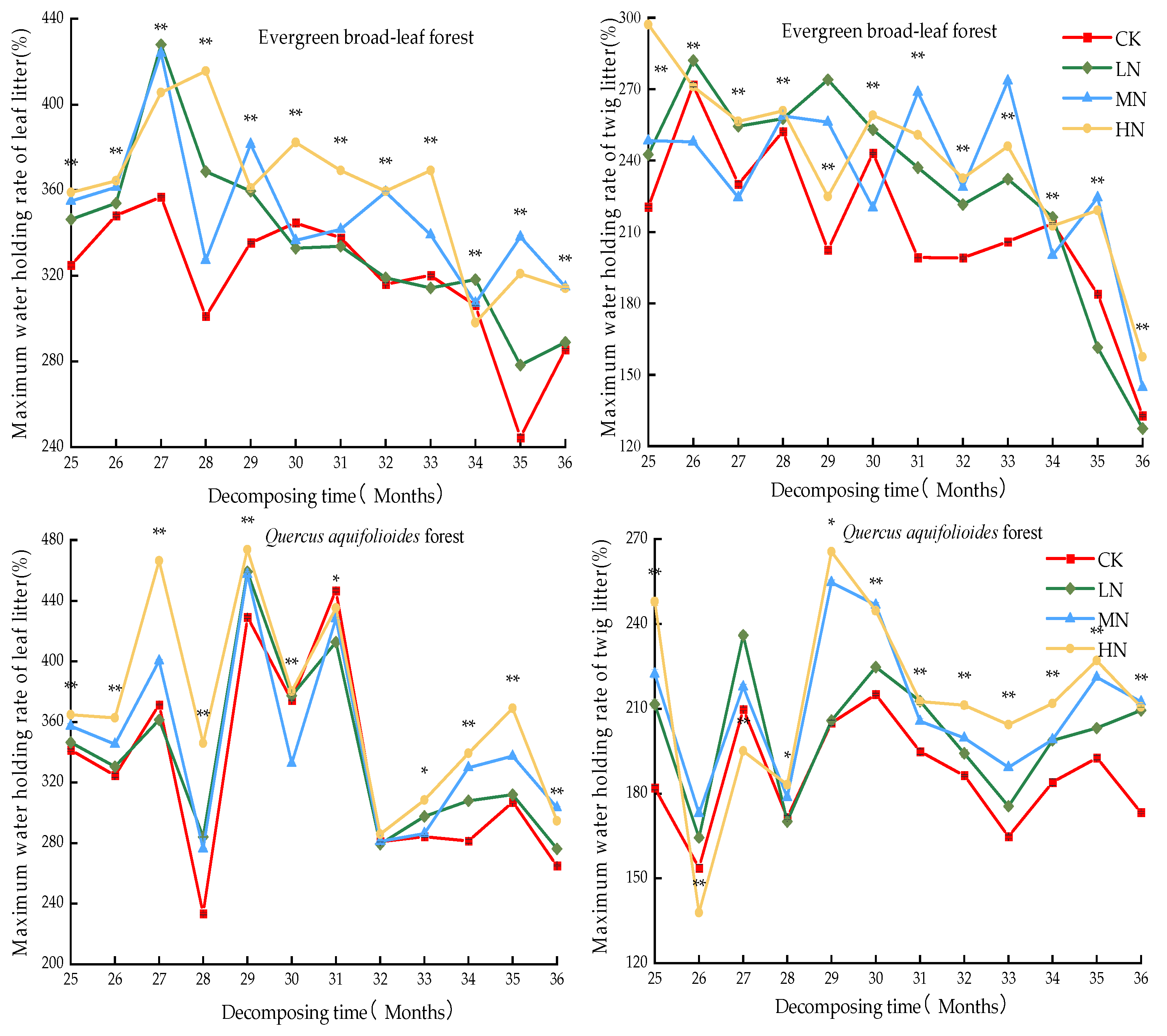
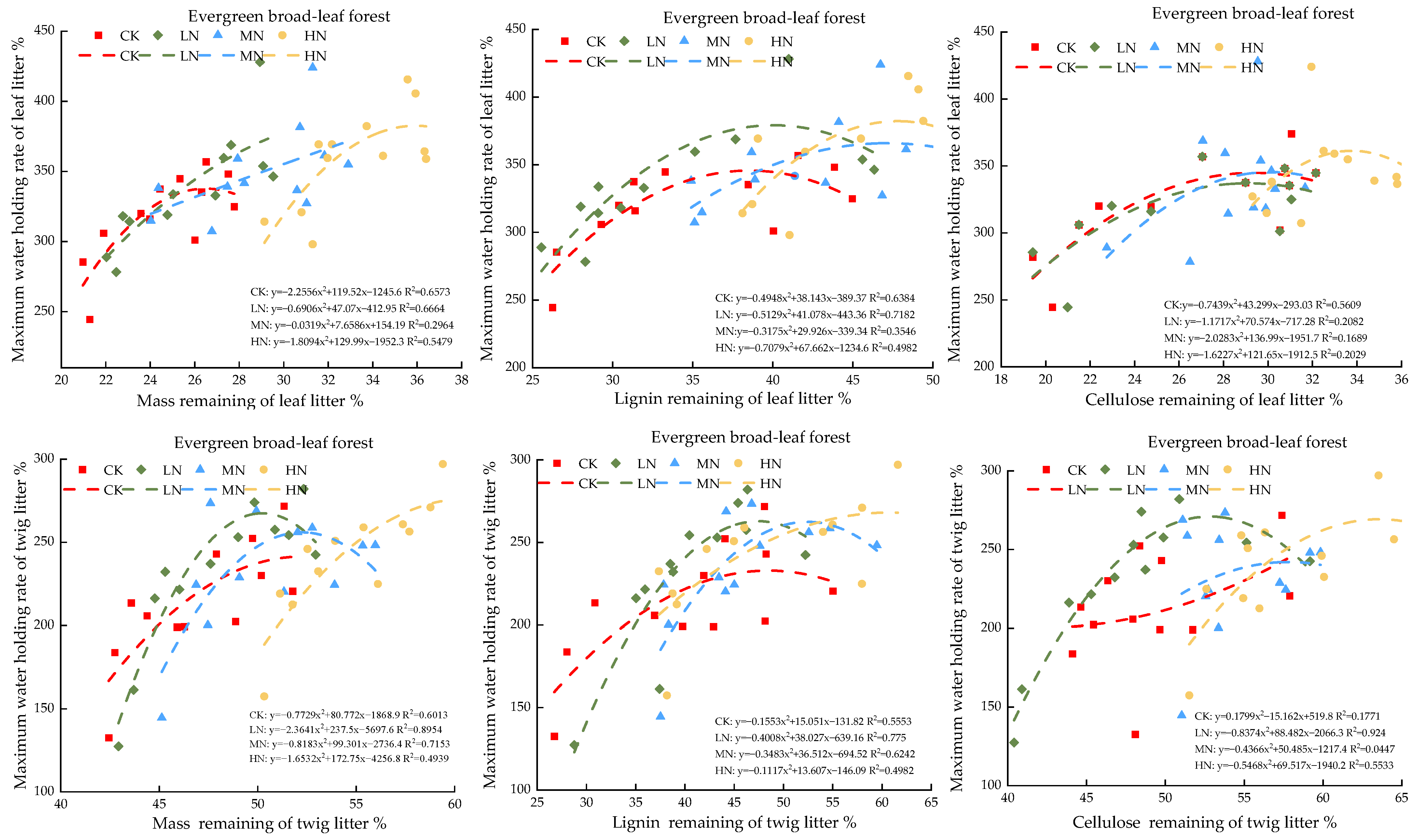
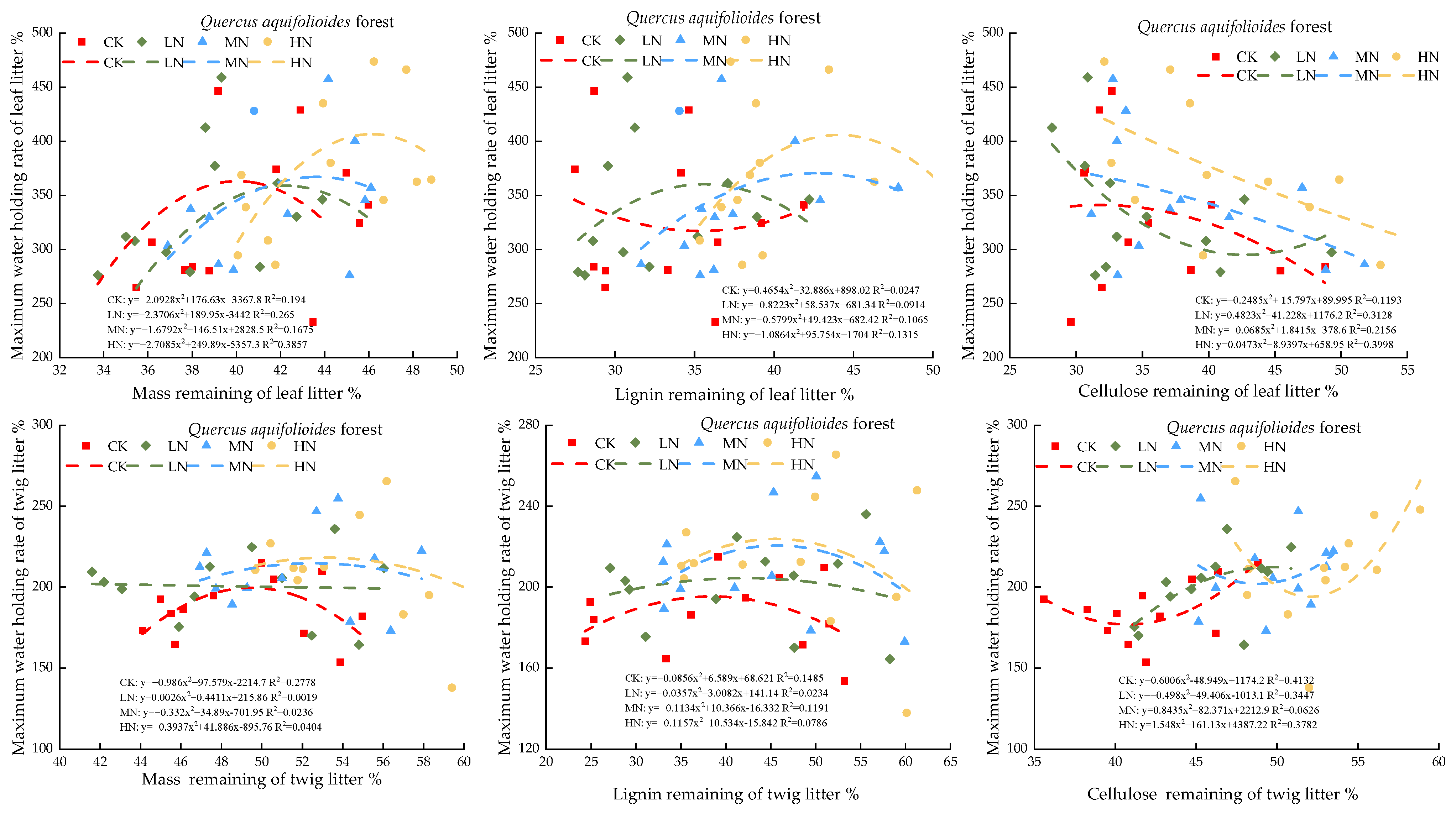
3.3. Impacts of Modeled Nitrogen Deposition on the Water Retention Ability of Leaf Litters
3.4. Analysis of the Correlation between Leaf and Twig Retention Rate and Peak Water Holding Capacity
4. Discussion
4.1. Simulation of the Effect of Nitrogen Deposition on Litter Decomposition
4.2. Simulation of the Effect of Nitrogen Deposition on Lignin and Cellulose Degradation in Litter
4.3. Simulation of the Effect of Nitrogen Deposition on the Water-Holding Characteristics of Litter
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, L.; Xu, W.; Wen, Z.; Liu, P.; Xu, H.; Liu, S.; Lu, X.; Zhong, B.; Guo, Y.; Lu, X.; et al. Modeling Global Oceanic Nitrogen Deposition from Food Systems and Its Mitigation Potential by Reducing Overuse of Fertilizers. Proc. Natl. Acad. Sci. USA 2023, 120, e2221459120. [Google Scholar] [CrossRef]
- Feng, H.; Guo, J.; Peng, C.; Kneeshaw, D.; Roberge, G.; Chang, P.; Ma, X.; Zhou, D.; Wang, W. Nitrogen Addition Promotes Terrestrial Plants to Allocate More Biomass to Aboveground Organs: A Global Meta-analysis. Glob. Change Biol. 2023, 29, 3970–3989. [Google Scholar] [CrossRef]
- Rowe, J.A.; Litton, C.M.; Lepczyk, C.A.; Popp, B.N. Impacts of Endangered Seabirds on Nutrient Cycling in Montane Forest Ecosystems of Hawai‘i. Pac. Sci. 2017, 71, 495–509. [Google Scholar] [CrossRef]
- Zhou, S.; Huang, C.; Han, B.; Xiao, Y.; Tang, J.; Xiang, Y.; Luo, C. Simulated Nitrogen Deposition Significantly Suppresses the Decomposition of Forest Litter in a Natural Evergreen Broad-Leaved Forest in the Rainy Area of Western China. Plant Soil 2017, 420, 135–145. [Google Scholar] [CrossRef]
- Song, S.; Hu, X.; Zhu, J.; Zheng, T.; Zhang, F.; Ji, C.; Zhu, J. The Decomposition Rates of Leaf Litter and Fine Root and Their Temperature Sensitivities Are Influenced Differently by Biotic Factors. Plant Soil 2021, 461, 603–616. [Google Scholar] [CrossRef]
- Yu, H.; Wang, J.; Wan, F.; Zhou, X.; Cai, M.; Ou, Q.; Li, W. Research progress on effects of plant litter on the decomposition of soil organic matter. J. Biosaf. 2018, 27, 88–94. [Google Scholar]
- Liu, Z.; Huang, X.; Tu, J.; Chen, C.; Ma, J.; Wang, K. Litter Reserves and Water Holding Characteristics of Different Species in Yunnan Plateau. Ecol. Environ. Sci. 2015, 24, 919–924. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, J.; Li, W.; Wu, D.; Fu, Y.; Jia, Z. Litter Reserves and Water Holding Characteristics of Rhododendron Forest in Baili Rhododendron Nature Reserve of Guizhou. J. Soil Water Conserv. 2018, 32, 167–173. [Google Scholar] [CrossRef]
- Klamerus-Iwan, A.; Lasota, J.; Błońska, E. Interspecific Variability of Water Storage Capacity and Absorbability of Deadwood. Forests 2020, 11, 575. [Google Scholar] [CrossRef]
- Chen, B.; Yang, X.; Zhao, X.; Wang, Y.; Tian, C.; Liu, Y.; Liu, P. Hydrological Effects of Six Natural Pure Forests Litters and Soil in Northern Mountain of Hebei Province. J. Soil Water Conserv. 2012, 26, 196–202. [Google Scholar] [CrossRef]
- Zheng, X.; Song, Y.; Wang, K.; Zhang, Y.; Pan, Y. Response of nutrient release and ecological stoichiometry of litter to simulated nitrogen deposition in evergreen broad-leaved forest in central Yunnan, China. Chin. J. Appl. Ecol. 2021, 32, 23–30. [Google Scholar] [CrossRef]
- Zhang, Y.; Song, Y.; Wang, K.; Yang, X.; Xing, J.; Zhang, Z. Responses of litter decomposition in two subalpine plantations to simulated nitrogen deposition in central Yunnan, China. Chin. J. Appl. Ecol. 2020, 31, 2523–2532. [Google Scholar] [CrossRef]
- Mao, Y.; Wang, K.; Song, Y.; Zhang, X.; Liang, Y.; Xiao, W. Responses of soil bacterial diversity and community structure to N sedimentation in alpine forests in central Yunnan. J. Cent. South Univ. For. Technol. 2023, 43, 125–137. [Google Scholar] [CrossRef]
- Zhang, N.; Song, Y.; Wang, K. Response of Enzyme Activity Characteristics of Forest Soil Aggregates to Nitrogen Deposition in Central Yunnan Based on Dry and Wet Screening Method. J. Soil Water Conserv. 2023, 37, 246–253. [Google Scholar] [CrossRef]
- Huang, X.; Chen, J.; Wang, D.; Deng, M.; Wu, M.; Tong, B.; Liu, J. Simulated Atmospheric Nitrogen Deposition Inhibited the Leaf Litter Decomposition of Cinnamomum migao H.W. Li in Southwest China. Sci. Rep. 2021, 11, 1748. [Google Scholar] [CrossRef]
- Chen, F.; Wang, G.; Fang, X.; Wan, S.; Zhang, Y.; Liang, C. Nitrogen Deposition Effect on Forest Litter Decomposition Is Interactively Regulated by Endogenous Litter Quality and Exogenous Resource Supply. Plant Soil 2019, 437, 413–426. [Google Scholar] [CrossRef]
- Hu, J.; Zhou, S.; Tie, L.; Liu, X.; Liu, X.; Zhao, A.; Lai, J.; Xiao, L.; You, C.; Huang, C. Effects of Nitrogen Addition on Soil Faunal Abundance: A Global Meta-Analysis. Glob. Ecol. Biogeogr. 2022, 31, 1655–1666. [Google Scholar] [CrossRef]
- Peguero, G.; Folch, E.; Liu, L.; Ogaya, R.; Peñuelas, J. Divergent Effects of Drought and Nitrogen Deposition on Microbial and Arthropod Soil Communities in a Mediterranean Forest. Eur. J. Soil Biol. 2021, 103, 103275. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.Y.H. Mapping Global Nitrogen Deposition Impacts on Soil Respiration. Sci. Total Environ. 2023, 871, 161986. [Google Scholar] [CrossRef] [PubMed]
- Forsmark, B.; Nordin, A.; Maaroufi, N.I.; Lundmark, T.; Gundale, M.J. Low and High Nitrogen Deposition Rates in Northern Coniferous Forests Have Different Impacts on Aboveground Litter Production, Soil Respiration, and Soil Carbon Stocks. Ecosystems 2020, 23, 1423–1436. [Google Scholar] [CrossRef]
- Payne, R.J.; Dise, N.B.; Field, C.D.; Dore, A.J.; Caporn, S.J.; Stevens, C.J. Nitrogen Deposition and Plant Biodiversity: Past, Present, and Future. Front. Ecol. Environ. 2017, 15, 431–436. [Google Scholar] [CrossRef]
- Lafuente, A.; Recio, J.; Ochoa-Hueso, R.; Gallardo, A.; Pérez-Corona, M.E.; Manrique, E.; Durán, J. Simulated Nitrogen Deposition Influences Soil Greenhouse Gas Fluxes in a Mediterranean Dryland. Sci. Total Environ. 2020, 737, 139610. [Google Scholar] [CrossRef] [PubMed]
- Nie, Y.; Han, X.; Chen, J.; Wang, M.; Shen, W. The Simulated N Deposition Accelerates Net N Mineralization and Nitrification in a Tropical Forest Soil. Biogeosciences 2019, 16, 4277–4291. [Google Scholar] [CrossRef]
- Yuan, X.; Niu, D.; Guo, D.; Fu, H. Responses of Soil Carbon and Nitrogen Mineralization to Nitrogen Addition in a Semiarid Grassland: The Role of Season. CATENA 2023, 220, 106719. [Google Scholar] [CrossRef]
- Liu, G.; Sun, J.; Tian, K.; Xiao, D.; Yuan, X. Long-term Responses of Leaf Litter Decomposition to Temperature, Litter Quality and Litter Mixing in Plateau Wetlands. Freshw. Biol. 2017, 62, 178–190. [Google Scholar] [CrossRef]
- Ilek, A.; Kucza, J.; Szostek, M. The Effect of Stand Species Composition on Water Storage Capacity of the Organic Layers of Forest Soils. Eur. J. For. Res. 2015, 134, 187–197. [Google Scholar] [CrossRef]
- Xing, Z.; Yan, D.; Wang, D.; Liu, S.; Dong, G. Experimental Analysis of the Effect of Forest Litter Cover on Surface Soil Water Dynamics under Continuous Rainless Condition in North China. Kuwait J. Sci. 2018, 45, 75–83. [Google Scholar]
- Du, J.; Niu, J.; Gao, Z.; Chen, X.; Zhang, L.; Li, X.; Van Doorn, N.S.; Luo, Z.; Zhu, Z. Effects of Rainfall Intensity and Slope on Interception and Precipitation Partitioning by Forest Litter Layer. CATENA 2019, 172, 711–718. [Google Scholar] [CrossRef]
- Zhou, Q.; Keith, D.M.; Zhou, X.; Cai, M.; Cui, X.; Wei, X.; Luo, Y. Comparing the Water-Holding Characteristics of Broadleaved, Coniferous, and Mixed Forest Litter Layers in a Karst Region. Mt. Res. Dev. 2018, 38, 220–229. [Google Scholar] [CrossRef]
- Bai, Y.; Zhou, Y.; Du, J.; Zhang, X.; Di, N. Effects of a Broadleaf-Oriented Transformation of Coniferous Plantations on the Hydrological Characteristics of Litter Layers in Subtropical China. Glob. Ecol. Conserv. 2021, 25, e01400. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, X.; Lu, X. The Composition, Seasonal Variation, and Potential Sources of the Atmospheric Wet Sulfur (S) and Nitrogen (N) Deposition in the Southwest of China. Environ. Sci. Pollut. Res. 2016, 23, 6363–6375. [Google Scholar] [CrossRef]
- Leng, Q.; Cui, J.; Zhou, F.; Du, K.; Zhang, L.; Fu, C.; Liu, Y.; Wang, H.; Shi, G.; Gao, M.; et al. Wet-Only Deposition of Atmospheric Inorganic Nitrogen and Associated Isotopic Characteristics in a Typical Mountain Area, Southwestern China. Sci. Total Environ. 2018, 616–617, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Yan, T.; Zhai, L.; Yen, H.; Liu, J.; Li, W.; Liu, H. Nitrogen Transport/Deposition from Paddy Ecosystem and Potential Pollution Risk Period in Southwest China. Water 2022, 14, 539. [Google Scholar] [CrossRef]
- Yu, G.; Jia, Y.; He, N.; Zhu, J.; Chen, Z.; Wang, Q.; Piao, S.; Liu, X.; He, H.; Guo, X.; et al. Stabilization of Atmospheric Nitrogen Deposition in China over the Past Decade. Nat. Geosci. 2019, 12, 424–429. [Google Scholar] [CrossRef]
- Truba, M.; Sosnowski, J. The Effect of Tytanit on Fibre Fraction Content in Medicago x varia T. Martyn and Trifolium pratense L. Cell Walls. Agriculture 2022, 12, 191. [Google Scholar] [CrossRef]
- Li, Y.; Li, B.; Zhang, X.; Chen, J.; Zhan, F.; Guo, X.; Zu, Y. Differential Water and Soil Conservation Capacity and Associated Processes in Four Forest Ecosystems in Dianchi Watershed, Yunnan Province, China. J. Soil Water Conserv. 2015, 70, 198. [Google Scholar] [CrossRef]
- Olson, J.S. Energy Storage and the Balance of Producers and Decomposers in Ecological Systems. Ecology 1963, 44, 322–331. [Google Scholar] [CrossRef]
- Zhuang, L.; Liu, Q.; Liang, Z.; You, C.; Tan, B.; Zhang, L.; Yin, R.; Yang, K.; Bol, R.; Xu, Z. Nitrogen Additions Retard Nutrient Release from Two Contrasting Foliar Litters in a Subtropical Forest, Southwest China. Forests 2020, 11, 377. [Google Scholar] [CrossRef]
- Renaudin, M.; Khlifa, R.; Legault, S.; Kembel, S.W.; Kneeshaw, D.; Moore, J.; Houle, D. Long-Term Simulated Nitrogen Deposition Has Moderate Impacts on Soil Microbial Communities across Three Bioclimatic Domains of the Eastern Canadian Forest. Forests 2023, 14, 1124. [Google Scholar] [CrossRef]
- Jing, H.; Wang, G. Temporal Dynamics of Pinus tabulaeformis Litter Decomposition under Nitrogen Addition on the Loess Plateau of China. For. Ecol. Manag. 2020, 476, 118465. [Google Scholar] [CrossRef]
- Song, X.; Li, Q.; Gu, H. Effect of Nitrogen Deposition and Management Practices on Fine Root Decomposition in Moso Bamboo Plantations. Plant Soil 2017, 410, 207–215. [Google Scholar] [CrossRef]
- Liu, R.; Zhang, Y.; Hu, X.-F.; Wan, S.; Wang, H.; Liang, C.; Chen, F. Litter Manipulation Effects on Microbial Communities and Enzymatic Activities Vary with Soil Depth in a Subtropical Chinese Fir Plantation. For. Ecol. Manag. 2021, 480, 118641. [Google Scholar] [CrossRef]
- Kong, B.; Zhou, J.; Qi, L.; Jiao, S.; Ma, L.; Geng, W.; Zhao, Y.; Gao, T.; Gong, J.; Li, K.; et al. Effects of Nitrogen Deposition on Leaf Litter Decomposition and Soil Organic Carbon Density in Arid and Barren Rocky Mountainous Regions: A Case Study of Yimeng Mountain. Forests 2023, 14, 1351. [Google Scholar] [CrossRef]
- Wu, J.; Liu, W.; Zhang, W.; Shao, Y.; Duan, H.; Chen, B.; Wei, X.; Fan, H. Long-Term Nitrogen Addition Changes Soil Microbial Community and Litter Decomposition Rate in a Subtropical Forest. Appl. Soil Ecol. 2019, 142, 43–51. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, G.; Wang, B.; Wang, J.; Xiao, J.; Li, Z. Litter Production and Its Water Holding Capability in Typical Plants Communities in the Hilly Region of the Loess Plateau. Acta Ecol. Sin. 2019, 39, 2416–2425. [Google Scholar] [CrossRef]
- Zhou, S.; Huang, C.; Xiang, Y.; Han, B.; Xiao, Y.; Tang, J. Effects of simulated nitrogen deposition on lignin and cellulose degradation of foliar litter in natural Evergreen broad-leaved forest in Rainy Area of Western China. Chin. J. Appl. Ecol. 2016, 27, 1368–1374. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, Y.; Wang, M.; Wang, K.; Meng, F.; Liu, L.; Zhao, Y.; Ma, L.; Zhu, Q.; Xu, W.; et al. Atmospheric Nitrogen Deposition: A Review of Quantification Methods and Its Spatial Pattern Derived from the Global Monitoring Networks. Ecotoxicol. Environ. Saf. 2021, 216, 112180. [Google Scholar] [CrossRef]
- Xu, W.; Zhao, Y.; Liu, X.; Dore, A.J.; Zhang, L.; Liu, L.; Cheng, M. Atmospheric Nitrogen Deposition in the Yangtze River Basin: Spatial Pattern and Source Attribution. Environ. Pollut. 2018, 232, 546–555. [Google Scholar] [CrossRef]
- Chen, Y.; Shen, H.; Shih, J.; Russell, A.G.; Shao, S.; Hu, Y.; Odman, M.T.; Nenes, A.; Pavur, G.K.; Zou, Y.; et al. Greater Contribution from Agricultural Sources to Future Reactive Nitrogen Deposition in the United States. Earth’s Future 2020, 8, e2019EF001453. [Google Scholar] [CrossRef]
- Xu, Y.; Fan, J.; Ding, W.; Bol, R.; Chen, Z.; Luo, J.; Bolan, N. Stage-Specific Response of Litter Decomposition to N and S Amendments in a Subtropical Forest Soil. Biol. Fertil. Soils 2016, 52, 711–724. [Google Scholar] [CrossRef]
- Wang, X.; Liao, W.; Xu, Z.; Zuo, X.; Fan, F.; Cao, S. Research Progress of Influencing Factors Affecting Forest Litter Decomposition. North. Hortic. 2022, 126–132. [Google Scholar]
- Berg, B.; Lönn, M.; Ni, X.; Sun, T.; Dong, L.; Gaitnieks, T.; Virzo De Santo, A.; Johansson, M.-B. Decomposition Rates in Late Stages of Scots Pine and Norway Spruce Needle Litter: Influence of Nutrients and Substrate Properties over a Climate Gradient. For. Ecol. Manag. 2022, 522, 120452. [Google Scholar] [CrossRef]
- Yue, K.; Peng, C.; Yang, W.; Peng, Y.; Zhang, C.; Huang, C.; Wu, F. Degradation of Lignin and Cellulose during Foliar Litter Decomposition in an Alpine Forest River. Ecosphere 2016, 7, 1–11. [Google Scholar] [CrossRef]
- He, M.; Zhao, R.; Tian, Q.; Huang, L.; Wang, X.; Liu, F. Predominant Effects of Litter Chemistry on Lignin Degradation in the Early Stage of Leaf Litter Decomposition. Plant Soil 2019, 442, 453–469. [Google Scholar] [CrossRef]
- Stevens, C.J. How Long Do Ecosystems Take to Recover from Atmospheric Nitrogen Deposition? Biol. Conserv. 2016, 200, 160–167. [Google Scholar] [CrossRef]
- Tie, L.; Zhang, S.; Xiong, Z.; Fu, R.; Zhou, S.; Huang, C. Effects of Simulated Nitrogen and Sulfur Deposition on Lignin Degradation during Foliar Litter Decomposition in Evergreen Broad-leaved Forest in the Rainy Area of West China. For. Res. 2019, 32, 25–31. [Google Scholar] [CrossRef]
- Tie, L.; Fu, R.; Zhang, S.; Zhou, S.; Huang, C. Effects of simulated nitrogen and sulfur deposition on cellulose degradation during foliar litter decomposition in evergreen broad-leaved forest in the Rainy Area of West China. Chin. J. Appl. Environ. Biol. 2019, 25, 16–22. [Google Scholar] [CrossRef]
- Zhang, P.; Lin, J.; Hao, J.; Li, C.; Quan, W. Decomposition Characteristics of Lignocellulosic Biomass in Subtropical Rhododendron Litters under Artificial Regulation. Metabolites 2023, 13, 279. [Google Scholar] [CrossRef]
- Zak, D.R.; Argiroff, W.A.; Freedman, Z.B.; Upchurch, R.A.; Entwistle, E.M.; Romanowicz, K.J. Anthropogenic N Deposition, Fungal Gene Expression, and an Increasing Soil Carbon Sink in the Northern Hemisphere. Ecology 2019, 100, e02804. [Google Scholar] [CrossRef]
- Jourgholami, M.; Sohrabi, H.; Venanzi, R.; Tavankar, F.; Picchio, R. Hydrologic Responses of Undecomposed Litter Mulch on Compacted Soil: Litter Water Holding Capacity, Runoff, and Sediment. CATENA 2022, 210, 105875. [Google Scholar] [CrossRef]
- Pang, Q. Study on the Effects of Litter and Soil Hydrology of Karst Forests in the South Subtropical Region. Geogr. Sci. Res. 2023, 12, 52–60. [Google Scholar] [CrossRef]
- Lisý, A.; Ház, A.; Nadányi, R.; Jablonský, M.; Šurina, I. About Hydrophobicity of Lignin: A Review of Selected Chemical Methods for Lignin Valorisation in Biopolymer Production. Energies 2022, 15, 6213. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forest Type | Stand | Altitude (m) | Age (a) | Mean Height (m) | DBH (cm) | Canopy Density | Slope (°) | Aspect | Soil Category |
|---|---|---|---|---|---|---|---|---|---|
| Evergreen broad-leaf forest | 1 | 2130 | 17 | 9.6 | 14.5 | 0.87 | 23 | NE | Argi-udic Ferrosols |
| 2 | 2132 | 15 | 12.1 | 20.7 | 0.90 | 28 | NE | Argi-udic Ferrosols | |
| 3 | 2133 | 17 | 10.8 | 18.3 | 0.85 | 30 | NE | Argi-udic Ferrosols | |
| Quercus aquifolioides | 1 | 2490 | 15 | 3.4 | 12.1 | 0.90 | 10 | SE | Hapli-udic argosols |
| 2 | 2489 | 16 | 2.5 | 9.4 | 0.88 | 12 | SE | Hapli-udic argosols | |
| 3 | 2490 | 17 | 3.1 | 8.6 | 0.92 | 13 | SE | Hapli-udic argosols |
| Forest Type | Litter Type | Treatment | Regression Equation | Determination Coefficient | Decomposition Coefficient | Time of Half Decomposition | Time of 95% Decomposition |
|---|---|---|---|---|---|---|---|
| R2 | (k, kg·kg−1·a−1) | (T50%, a) | (T95%, a) | ||||
| Evergreen broad-leaf forest | Leaf litter | CK | y = 29.105 e −0.316 t | 0.959 ** | 0.316 | 2.194 | 9.480 |
| LN | y = 31.101 e −0.355 t | 0.969 ** | 0.355 | 1.953 | 8.439 | ||
| MN | y = 34.475 e −0.333 t | 0.935 ** | 0.332 | 2.088 | 9.023 | ||
| HN | y = 37.844 e −0.242 t | 0.964 ** | 0.242 | 2.864 | 12.379 | ||
| Twig litter | CK | y = 53.376 e −0.236 t | 0.989 ** | 0.236 | 2.937 | 12.694 | |
| LN | y = 54.664 e −0.242 t | 0.990 ** | 0.242 | 2.864 | 12.379 | ||
| MN | y = 57.190 e −0.229 t | 0.992 ** | 0.229 | 3.027 | 13.082 | ||
| HN | y = 60.557 e −0.187 t | 0.994 ** | 0.187 | 3.707 | 16.020 | ||
| Quercus aquifolioides | Leaf litter | CK | y = 47.913 e −0.303 t | 0.981 ** | 0.303 | 2.288 | 9.887 |
| LN | y = 44.852 e −0.274 t | 0.988 ** | 0.274 | 2.530 | 10.933 | ||
| MN | y = 48.168 e −0.264 t | 0.972 ** | 0.264 | 2.626 | 11.347 | ||
| HN | y = 50.304 e −0.246 t | 0.972 ** | 0.246 | 2.818 | 12.178 | ||
| Twig litter | CK | y = 56.156 e −0.256 t | 0.973 ** | 0.256 | 2.708 | 11.702 | |
| LN | y = 58.294 e −0.341 t | 0.990 ** | 0.341 | 2.033 | 8.785 | ||
| MN | y = 58.962 e −0.244 t | 0.984 ** | 0.244 | 2.841 | 12.278 | ||
| HN | y = 61.345 e −0.220 t | 0.982 ** | 0.220 | 3.151 | 13.617 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xing, J.; Hu, C.; Song, C.; Wang, K.; Song, Y. Nitrogen Deposition Modulates Litter Decomposition and Enhances Water Retention in Subtropical Forests. Forests 2024, 15, 522. https://doi.org/10.3390/f15030522
Xing J, Hu C, Song C, Wang K, Song Y. Nitrogen Deposition Modulates Litter Decomposition and Enhances Water Retention in Subtropical Forests. Forests. 2024; 15(3):522. https://doi.org/10.3390/f15030522
Chicago/Turabian StyleXing, Jinmei, Chun Hu, Chenggong Song, Keqin Wang, and Yali Song. 2024. "Nitrogen Deposition Modulates Litter Decomposition and Enhances Water Retention in Subtropical Forests" Forests 15, no. 3: 522. https://doi.org/10.3390/f15030522
APA StyleXing, J., Hu, C., Song, C., Wang, K., & Song, Y. (2024). Nitrogen Deposition Modulates Litter Decomposition and Enhances Water Retention in Subtropical Forests. Forests, 15(3), 522. https://doi.org/10.3390/f15030522