
Soil Respiration after Bark Beetle Infestation along a Vertical Transect in Mountain Spruce Forest


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Soil Respiration Measurements
2.3. Statistical Analysis
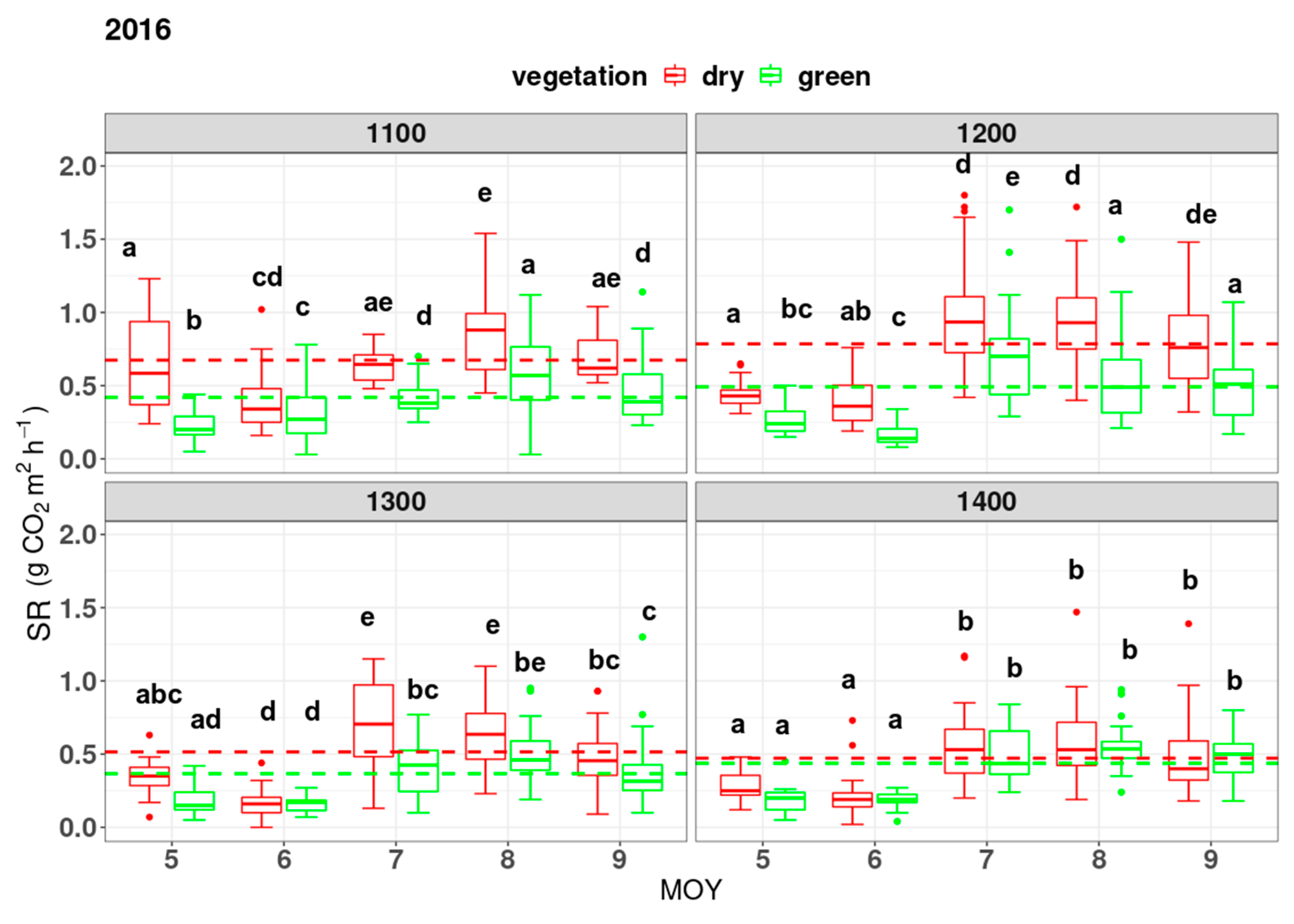
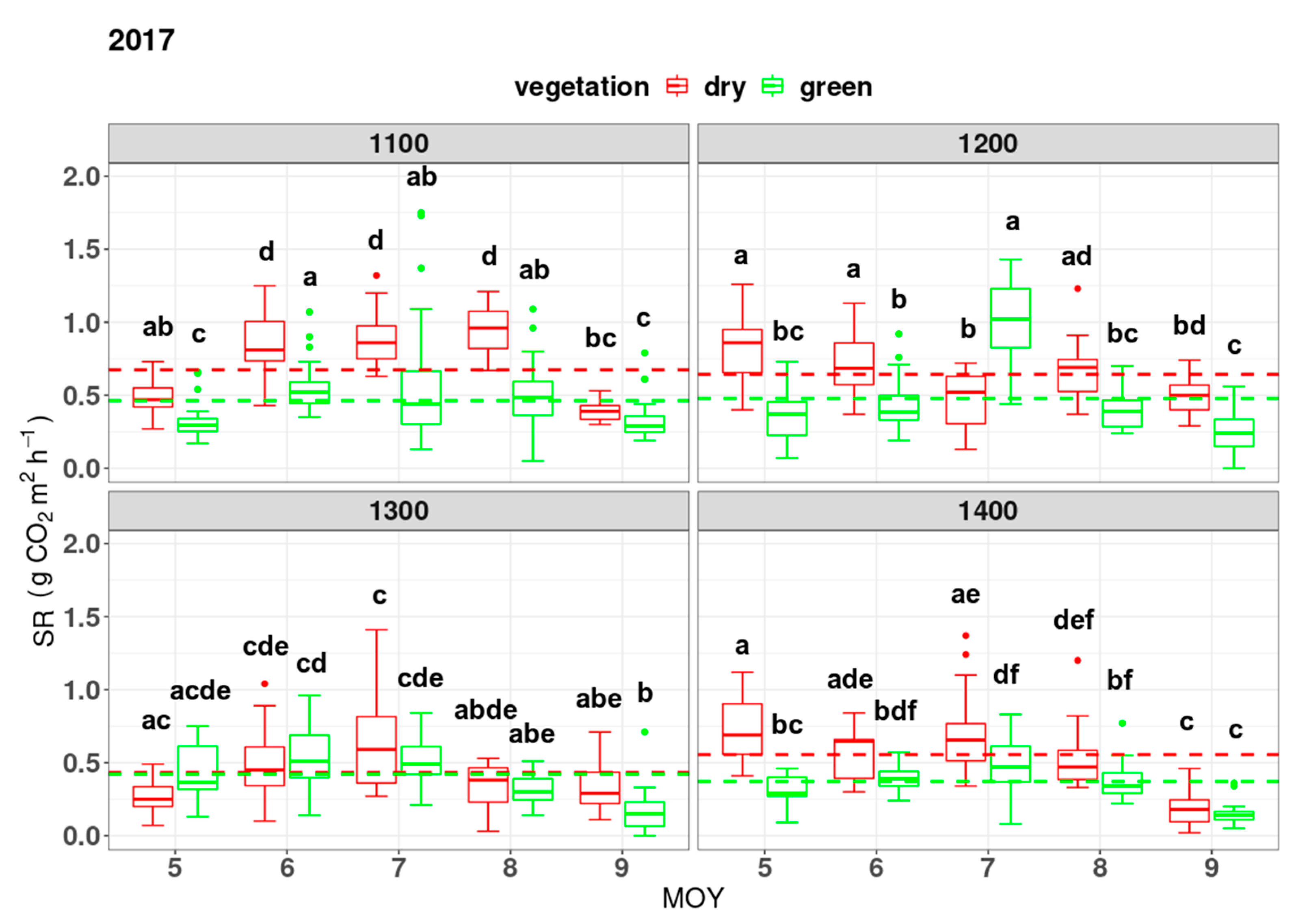
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Canadell, J.G.; Monteiro, P.M.S.; Costa, M.H.; Cotrim da Cunha, L.; Cox, P.M.; Eliseev, A.V.; Henson, S.; Ishii, M.; Jaccard, S.; Koven, C.; et al. 2021: Global Carbon and other Biogeochemical Cycles and Feedbacks. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2023; pp. 673–816. [Google Scholar] [CrossRef]
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A Large and Persistent Carbon Sink in the World’s Forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef] [PubMed]
- Kirschbaum, M.U.F. Will Changes in Soil Organic Carbon Act as a Positive or Negative Feedback on Global Warming? Biogeochemistry 2000, 48, 21–51. [Google Scholar] [CrossRef]
- Hashimoto, S. A New Estimation of Global Soil Greenhouse Gas Fluxes Using a Simple Data-Oriented Model. PLoS ONE 2012, 7, e41962. [Google Scholar] [CrossRef]
- Bond-Lamberty, B.; Thomson, A. Temperature-Associated Increases in the Global Soil Respiration Record. Nature 2010, 464, 579–582. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, H.; Spooner, F.; Roser, M. Forests and Deforestation. Our World Data 2023. Available online: https://ourworldindata.org/forests-and-deforestation (accessed on 28 October 2023).
- Randerson, J.T.; Chapin, F.S.; Harden, J.W.; Neff, J.C.; Harmon, M.E. Net Ecosystem Production: A Comprehensive Measure of Net Carbon Accumulation by Ecosystems. Ecol. Appl. 2002, 12, 937–947. [Google Scholar] [CrossRef]
- Lindauer, M.; Schmid, H.P.; Grote, R.; Mauder, M.; Steinbrecher, R.; Wolpert, B. Net Ecosystem Exchange over a Non-Cleared Wind-Throw-Disturbed Upland Spruce Forest—Measurements and Simulations. Agric. For. Meteorol. 2014, 197, 219–234. [Google Scholar] [CrossRef]
- Yamanoi, K.; Mizoguchi, Y.; Utsugi, H. Effects of a Windthrow Disturbance on the Carbon Balance of a Broadleaf Deciduous Forest in Hokkaido, Japan. Biogeosciences 2015, 12, 6837–6851. [Google Scholar] [CrossRef]
- Hermle, S.; Lavigne, M.B.; Bernier, P.Y.; Bergeron, O.; Pare, D. Component Respiration, Ecosystem Respiration and Net Primary Production of a Mature Black Spruce Forest in Northern Quebec. Tree Physiol. 2010, 30, 527–540. [Google Scholar] [CrossRef]
- IPCC. Summary for Policymakers. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2021; pp. 3–32. [Google Scholar] [CrossRef]
- Pörtner, H.-O.; Roberts, D.C.; Adams, H.; Adelekan, I.; Adler, C.; Adrian, R.; Aldunce, P.; Ali, E.; Begum, R.A.; Bednar-Friedl, B.; et al. Technical Summary. In Climate Change 2022: Impacts, Adaptation and Vulnerability. Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Pörtner, H.-O., Roberts, D.C., Tignor, M.M.B., Poloczanska, E.S., Mintenbeck, K., Alegría, A., Craig, M., Langsdorf, S., Löschke, S., Möller, V., et al., Eds.; Cambridge University Press: Cambridge, UK, 2022. [Google Scholar]
- Jakoby, O.; Lischke, H.; Wermelinger, B. Climate Change Alters Elevational Phenology Patterns of the European Spruce Bark Beetle (Ips Typographus). Glob. Chang. Biol. 2019, 25, 4048–4063. [Google Scholar] [CrossRef]
- Romashkin, I.; Neuvonen, S.; Tikkanen, O. Northward Shift in Temperature Sum Isoclines May Favour Ips typographus Outbreaks in European Russia. Agric. For. Entomol. 2020, 22, 238–249. [Google Scholar] [CrossRef]
- Marini, L.; Ayres, M.P.; Battisti, A.; Faccoli, M. Climate Affects Severity and Altitudinal Distribution of Outbreaks in an Eruptive Bark Beetle. Clim. Chang. 2012, 115, 327–341. [Google Scholar] [CrossRef]
- Seidl, R.; Schelhaas, M.-J.; Rammer, W.; Verkerk, P.J. Increasing Forest Disturbances in Europe and Their Impact on Carbon Storage. Nat. Clim. Chang. 2014, 4, 806–810. [Google Scholar] [CrossRef]
- Hlásny, T.; König, L.; Krokene, P.; Lindner, M.; Montagné-Huck, C.; Müller, J.; Qin, H.; Raffa, K.F.; Schelhaas, M.-J.; Svoboda, M.; et al. Bark Beetle Outbreaks in Europe: State of Knowledge and Ways Forward for Management. Curr. For. Rep. 2021, 7, 138–165. [Google Scholar] [CrossRef]
- Trubin, A.; Mezei, P.; Zabihi, K.; Surový, P.; Jakuš, R. Northernmost European Spruce Bark Beetle Ips typographus Outbreak: Modelling Tree Mortality Using Remote Sensing and Climate Data. For. Ecol. Manag. 2022, 505, 119829. [Google Scholar] [CrossRef]
- Bright, B.C.; Hicke, J.A.; Meddens, A.J.H. Effects of Bark Beetle-caused Tree Mortality on Biogeochemical and Biogeophysical MODIS Products. J. Geophys. Res. Biogeosciences 2013, 118, 974–982. [Google Scholar] [CrossRef]
- Edburg, S.L.; Hicke, J.A.; Brooks, P.D.; Pendall, E.G.; Ewers, B.E.; Norton, U.; Gochis, D.; Gutmann, E.D.; Meddens, A.J. Cascading Impacts of Bark Beetle-caused Tree Mortality on Coupled Biogeophysical and Biogeochemical Processes. Front. Ecol. Environ. 2012, 10, 416–424. [Google Scholar] [CrossRef]
- Kurz, W.A.; Dymond, C.C.; Stinson, G.; Rampley, G.J.; Neilson, E.T.; Carroll, A.L.; Ebata, T.; Safranyik, L. Mountain Pine Beetle and Forest Carbon Feedback to Climate Change. Nature 2008, 452, 987–990. [Google Scholar] [CrossRef]
- Moore, D.J.P.; Trahan, N.A.; Wilkes, P.; Quaife, T.; Stephens, B.B.; Elder, K.; Desai, A.R.; Negron, J.; Monson, R.K. Persistent Reduced Ecosystem Respiration after Insect Disturbance in High Elevation Forests. Ecol. Lett. 2013, 16, 731–737. [Google Scholar] [CrossRef]
- Xiong, Y.; D’Atri, J.J.; Fu, S.; Xia, H.; Seastedt, T.R. Rapid Soil Organic Matter Loss from Forest Dieback in a Subalpine Coniferous Ecosystem. Soil Biol. Biochem. 2011, 43, 2450–2456. [Google Scholar] [CrossRef]
- Hu, Y.-H.; Zhang, X.-Y.; Zhang, K.; Song, M.-H.; Gao, J.-Q.; Dorodnikov, M.; Soromotin, A.; Kuzyakov, Y. Tussock Microhabitats Increase Nitrogen Uptake by Plants in an Alpine Wetland. Plant Soil 2021, 466, 569–580. [Google Scholar] [CrossRef]
- Powlson, D.S.; Addiscott, T.M. Nitrogen in Soils|Nitrates. In Encyclopedia of Soils in the Environment; Elsevier: Amsterdam, The Netherlands, 2005; pp. 21–31. [Google Scholar] [CrossRef]
- Cigan, P.W.; Karst, J.; Cahill, J.F.; Sywenky, A.N.; Pec, G.J.; Erbilgin, N. Influence of Bark Beetle Outbreaks on Nutrient Cycling in Native Pine Stands in Western Canada. Plant Soil 2015, 390, 29–47. [Google Scholar] [CrossRef]
- Högberg, M.N.; Högberg, P. Extramatrical Ectomycorrhizal Mycelium Contributes One-third of Microbial Biomass and Produces, Together with Associated Roots, Half the Dissolved Organic Carbon in a Forest Soil. New Phytol. 2002, 154, 791–795. [Google Scholar] [CrossRef]
- Luo, Y.; Zhou, X. Soil Respiration and the Environment; Elsevier: Amsterdam, The Netherlands, 2006. [Google Scholar]
- Kobler, J.; Jandl, R.; Dirnböck, T.; Mirtl, M.; Schindlbacher, A. Effects of Stand Patchiness Due to Windthrow and Bark Beetle Abatement Measures on Soil CO2 Efflux and Net Ecosystem Productivity of a Managed Temperate Mountain Forest. Eur. J. For. Res. 2015, 134, 683–692. [Google Scholar] [CrossRef]
- Trahan, N.A.; Dynes, E.L.; Pugh, E.; Moore, D.J.P.; Monson, R.K. Changes in Soil Biogeochemistry Following Disturbance by Girdling and Mountain Pine Beetles in Subalpine Forests. Oecologia 2015, 177, 981–995. [Google Scholar] [CrossRef]
- Jaakkola, E.; Gärtner, A.; Jönsson, A.M.; Ljung, K.; Olsson, P.-O.; Holst, T. Spruce Bark Beetle (Ips typographus) Infestation Cause up to 700 Times Higher Bark BVOC Emission Rates from Norway Spruce (Picea Abies); preprint; Biodiversity and Ecosystem Function. Terrestrial 2022, 7, 1–32. [Google Scholar] [CrossRef]
- Swanson, M.E.; Franklin, J.F.; Beschta, R.L.; Crisafulli, C.M.; DellaSala, D.A.; Hutto, R.L.; Lindenmayer, D.B.; Swanson, F.J. The Forgotten Stage of Forest Succession: Early-successional Ecosystems on Forest Sites. Front. Ecol. Environ. 2011, 9, 117–125. [Google Scholar] [CrossRef]
- Morehouse, K.; Johns, T.; Kaye, J.; Kaye, M. Carbon and Nitrogen Cycling Immediately Following Bark Beetle Outbreaks in Southwestern Ponderosa Pine Forests. For. Ecol. Manag. 2008, 255, 2698–2708. [Google Scholar] [CrossRef]
- Reed, D.E.; Ewers, B.E.; Pendall, E. Impact of Mountain Pine Beetle Induced Mortality on Forest Carbon and Water Fluxes. Environ. Res. Lett. 2014, 9, 105004. [Google Scholar] [CrossRef]
- Mayer, M.; Matthews, B.; Rosinger, C.; Sandén, H.; Godbold, D.L.; Katzensteiner, K. Tree Regeneration Retards Decomposition in a Temperate Mountain Soil after Forest Gap Disturbance. Soil Biol. Biochem. 2017, 115, 490–498. [Google Scholar] [CrossRef]
- Hursh, A.; Ballantyne, A.; Cooper, L.; Maneta, M.; Kimball, J.; Watts, J. The Sensitivity of Soil Respiration to Soil Temperature, Moisture, and Carbon Supply at the Global Scale. Glob. Chang. Biol. 2017, 23, 2090–2103. [Google Scholar] [CrossRef]
- Chen, S.; Huang, Y.; Zou, J.; Shen, Q.; Hu, Z.; Qin, Y.; Chen, H.; Pan, G. Modeling Interannual Variability of Global Soil Respiration from Climate and Soil Properties. Agric. For. Meteorol. 2010, 150, 590–605. [Google Scholar] [CrossRef]
- Tang, J.; Bradford, M.A.; Carey, J.; Crowther, T.W.; Machmuller, M.B.; Mohan, J.E.; Todd-Brown, K. Temperature Sensitivity of Soil Carbon. In Ecosystem Consequences of Soil Warming; Elsevier: Amsterdam, The Netherlands, 2019; pp. 175–208. [Google Scholar] [CrossRef]
- Wang, G.; Zhao, M.; Kang, M.; Xing, K.; Wang, Y.; Xue, F.; Chen, C. Diurnal and Seasonal Variation of the Elevation Gradient of Air Temperature in the Northern Flank of the Western Qinling Mountain Range, China. J. Mt. Sci. 2017, 14, 94–105. [Google Scholar] [CrossRef]
- Chen, B.-X.; Sun, Y.-F.; Zhang, H.-B.; Han, Z.-H.; Wang, J.-S.; Li, Y.-K.; Yang, X.-L. Temperature Change along Elevation and Its Effect on the Alpine Timberline Tree Growth in the Southeast of the Tibetan Plateau. Adv. Clim. Change Res. 2018, 9, 185–191. [Google Scholar] [CrossRef]
- Makita, N.; Kosugi, Y.; Sakabe, A.; Kanazawa, A.; Ohkubo, S.; Tani, M. Seasonal and Diurnal Patterns of Soil Respiration in an Evergreen Coniferous Forest: Evidence from Six Years of Observation with Automatic Chambers. PLoS ONE 2018, 13, e0192622. [Google Scholar] [CrossRef] [PubMed]
- Speckman, H.N.; Frank, J.M.; Bradford, J.B.; Miles, B.L.; Massman, W.J.; Parton, W.J.; Ryan, M.G. Forest Ecosystem Respiration Estimated from Eddy Covariance and Chamber Measurements under High Turbulence and Substantial Tree Mortality from Bark Beetles. Glob. Chang. Biol. 2015, 21, 708–721. [Google Scholar] [CrossRef]
- Badraghi, A.; Ventura, M.; Polo, A.; Borruso, L.; Giammarchi, F.; Montagnani, L. Soil Respiration Variation along an Altitudinal Gradient in the Italian Alps: Disentangling Forest Structure and Temperature Effects. PLoS ONE 2021, 16, e0247893. [Google Scholar] [CrossRef]
- DeForest, J.L.; Noormets, A.; McNulty, S.G.; Sun, G.; Tenney, G.; Chen, J. Phenophases Alter the Soil Respiration–Temperature Relationship in an Oak-Dominated Forest. Int. J. Biometeorol. 2006, 51, 135–144. [Google Scholar] [CrossRef]
- Mathes, K.; Schriefer, T. Soil Respiration during Secondary Succession: Influence of Temperature and Moisture. Soil Biol. Biochem. 1985, 17, 205–211. [Google Scholar] [CrossRef]
- Davidson, E.A.; Holbrook, N.M. Is Temporal Variation of Soil Respiration Linked to the Phenology of Photosynthesis? In Phenology of Ecosystem Processes; Noormets, A., Ed.; Springer New York: New York, NY, USA, 2009; pp. 187–199. [Google Scholar] [CrossRef]
- Prach, K.; Pyšek, P. How Do Species Dominating in Succession Differ from Others? J. Veg. Sci. 1999, 10, 383–392. [Google Scholar] [CrossRef]
- Sedláková, I.; Fiala, K. Ecological Problems of Degradation of Alluvial Meadows Due to Expanding Calamagrostis Epigejos. Ekol. Bratisl. 2001, 20, 226–233. [Google Scholar]
- Mayer, M.; Matthews, B.; Schindlbacher, A.; Katzensteiner, K. Soil CO2 Efflux from Mountainous Windthrow Areas: Dynamics over 12 Years Post-Disturbance. Biogeosciences 2014, 11, 6081–6093. [Google Scholar] [CrossRef]
- Budzáková, M.; Galvánek, D.; Littera, P.; Šibík, J. The Wind and Fire Disturbance in Central European Mountain Spruce Forests: The Regeneration after Four Years. Acta Soc. Bot. Pol. 2013, 81, 13–24. [Google Scholar] [CrossRef]
- Pyšek, P. What Do We Know about Calamagrostis Villosa? A Review of the Species Behaviour in Secondary Habitats. Preslia 1993, 65, 1–20. [Google Scholar]
- Chang, S.X.; Shi, Z.; Thomas, B.R. Soil Respiration and Net Ecosystem Productivity in a Chronosequence of Hybrid Poplar Plantations1. Can. J. Soil Sci. 2020, 100, 488–502. [Google Scholar] [CrossRef]
- Macdonald, C.A.; Anderson, I.C.; Khachane, A.; Singh, B.P.; Barton, C.V.M.; Duursma, R.A.; Ellsworth, D.S.; Singh, B.K. Plant Productivity Is a Key Driver of Soil Respiration Response to Climate Change in a Nutrient-Limited Soil. Basic Appl. Ecol. 2021, 50, 155–168. [Google Scholar] [CrossRef]
- Caprez, R.; Niklaus, P.A.; Körner, C. Forest Soil Respiration Reflects Plant Productivity across a Temperature Gradient in the Alps. Oecologia 2012, 170, 1143–1154. [Google Scholar] [CrossRef]
- Rodeghiero, M.; Cescatti, A. Main Determinants of Forest Soil Respiration along an Elevation/Temperature Gradient in the Italian Alps. Glob. Chang. Biol. 2005, 11, 1024–1041. [Google Scholar] [CrossRef]
- Zheng, X.; Zhao, C.; Peng, S.; Jian, S.; Liang, B.; Wang, X.; Yang, S.; Wang, C.; Peng, H.; Wang, Y. Soil CO2 Efflux along an Elevation Gradient in Qinghai Spruce Forests in the Upper Reaches of the Heihe River, Northwest China. Environ. Earth Sci. 2014, 71, 2065–2076. [Google Scholar] [CrossRef]
- Borkhuu, B.; Peckham, S.D.; Ewers, B.E.; Norton, U.; Pendall, E. Does Soil Respiration Decline Following Bark Beetle Induced Forest Mortality? Evidence from a Lodgepole Pine Forest. Agric. For. Meteorol. 2015, 214, 201–207. [Google Scholar] [CrossRef]
- Jamnická, G.; Konôpková, A.; Fleischer, P.; Kurjak, D.; Petrík, P.; Petek, A.; Húdoková, H.; Fleischer, P.; Homolová, Z.; Ježík, M.; et al. Physiological Vitality of Norway Spruce (Picea Abies L.) Stands along an Altitudinal Gradient in Tatra National Park. Cent. Eur. For. J. 2020, 66, 227–242. [Google Scholar] [CrossRef]
- Fleischer, P.; Homolová, Z. Tatry Ako Objekt Dlhodobého Ekologického Výskumu Prírodných Disturbancií. Životné Prostr. 2016, 50, 1–4. [Google Scholar]
- Fleischer, P.; Homolová, Z. Long-Term Research on Ecological Condition in the Larch-Spruce Forest in High Tatras after Natural Disturbances. Lesn. Čas. For. J. 2011, 57, 237–250. [Google Scholar]
- Elkin, L.A.; Kay, M.; Higgins, J.J.; Wobbrock, J.O. An Aligned Rank Transform Procedure for Multifactor Contrast Tests. In Proceedings of the 34th Annual ACM Symposium on User Interface Software and Technology, Virtual, 10–14 October 2021; pp. 754–768. [Google Scholar] [CrossRef]
- Lei, N.; Wang, H.; Zhang, Y.; Chen, T. Components of Respiration and Their Temperature Sensitivity in Four Reconstructed Soils. Sci. Rep. 2022, 12, 6107. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, Y.; Sha, L.; Wu, C.; Tan, Z.; Song, Q.; Liu, Y.; Dong, L. Effects of Continuous Drought Stress on Soil Respiration in a Tropical Rainforest in Southwest China. Plant Soil 2015, 394, 343–353. [Google Scholar] [CrossRef]
- Tang, X.; Zhou, G.; Liu, S.; Zhang, D.; Liu, S.; Li, J.; Zhou, C. Dependence of Soil Respiration on Soil Temperature and Soil Moisture in Successional Forests in Southern China. J. Integr. Plant Biol. 2006, 48, 654–663. [Google Scholar] [CrossRef]
- Högberg, P.; Nordgren, A.; Buchmann, N.; Taylor, A.F.; Ekblad, A.; Högberg, M.N.; Nyberg, G.; Ottosson-Löfvenius, M.; Read, D.J. Large-Scale Forest Girdling Shows That Current Photosynthesis Drives Soil Respiration. Nature 2001, 411, 789–792. [Google Scholar] [CrossRef]
- Levy-Varon, J.H.; Schuster, W.S.F.; Griffin, K.L. Rapid Rebound of Soil Respiration Following Partial Stand Disturbance by Tree Girdling in a Temperate Deciduous Forest. Oecologia 2014, 174, 1415–1424. [Google Scholar] [CrossRef] [PubMed]
- Brouillard, B.M.; Mikkelson, K.M.; Bokman, C.M.; Berryman, E.M.; Sharp, J.O. Extent of Localized Tree Mortality Influences Soil Biogeochemical Response in a Beetle-Infested Coniferous Forest. Soil Biol. Biochem. 2017, 114, 309–318. [Google Scholar] [CrossRef]
- Han, M.; Feng, J.; Chen, Y.; Sun, L.; Fu, L.; Zhu, B. Mycorrhizal Mycelial Respiration: A Substantial Component of Soil Respired CO2. Soil Biol. Biochem. 2021, 163, 108454. [Google Scholar] [CrossRef]
- Klutsch, J.G.; Negrón, J.F.; Costello, S.L.; Rhoades, C.C.; West, D.R.; Popp, J.; Caissie, R. Stand Characteristics and Downed Woody Debris Accumulations Associated with a Mountain Pine Beetle (Dendroctonus Ponderosae Hopkins) Outbreak in Colorado. For. Ecol. Manag. 2009, 258, 641–649. [Google Scholar] [CrossRef]
- Whitney, G.G. The Productivity and Carbohydrate Economy of a Developing Stand of Rubus Idaeus. Can. J. Bot. 1982, 60, 2697–2703. [Google Scholar] [CrossRef]
- Tang, X.; Pei, X.; Lei, N.; Luo, X.; Liu, L.; Shi, L.; Chen, G.; Liang, J. Global Patterns of Soil Autotrophic Respiration and Its Relation to Climate, Soil and Vegetation Characteristics. Geoderma 2020, 369, 114339. [Google Scholar] [CrossRef]
- Zeeshan, M.; Wenjun, Z.; Chuansheng, W.; Yan, L.; Azeez, P.A.; Qinghai, S.; Yuntong, L.; Yiping, Z.; Zhiyun, L.; Liqing, S. Soil Heterotrophic Respiration in Response to Rising Temperature and Moisture along an Altitudinal Gradient in a Subtropical Forest Ecosystem, Southwest China. Sci. Total Environ. 2022, 816, 151643. [Google Scholar] [CrossRef] [PubMed]
- Yazaki, T.; Hirano, T.; Sano, T. Biomass Accumulation and Net Primary Production during the Early Stage of Secondary Succession after a Severe Forest Disturbance in Northern Japan. Forests 2016, 7, 287. [Google Scholar] [CrossRef]
- Norton, U.; Ewers, B.E.; Borkhuu, B.; Brown, N.R.; Pendall, E. Soil Nitrogen Five Years after Bark Beetle Infestation in Lodgepole Pine Forests. Soil Sci. Soc. Am. J. 2015, 79, 282–293. [Google Scholar] [CrossRef]
- Kubov, M.; Janík, R.; Tomes, J.; Schieber, B. Nutrient Content in Biomass of Bilberry (Vaccinium Myrtillus L.) in Different Habitats of Protected Areas of Inner Western Carpathians. J. For. Sci. 2024; 70, in press. [Google Scholar]
- Mayer, M.; Sandén, H.; Rewald, B.; Godbold, D.; Katzensteiner, K. Increase in Heterotrophic Soil Respiration by Temperature Drives Decline in Soil Organic Carbon Stocks after Forest Windthrow in a Mountainous Ecosystem. Funct. Ecol. 2017, 31, 1163–1172. [Google Scholar] [CrossRef]
- Maurer, G.E.; Chan, A.M.; Trahan, N.A.; Moore, D.J.P.; Bowling, D.R. Carbon Isotopic Composition of Forest Soil Respiration in the Decade Following Bark Beetle and Stem Girdling Disturbances in the Rocky Mountains. Plant Cell Environ. 2016, 39, 1513–1523. [Google Scholar] [CrossRef] [PubMed]
- Watts, J.D.; Natali, S.M.; Minions, C.; Risk, D.; Arndt, K.; Zona, D.; Euskirchen, E.S.; Rocha, A.V.; Sonnentag, O.; Helbig, M.; et al. Soil Respiration Strongly Offsets Carbon Uptake in Alaska and Northwest Canada. Environ. Res. Lett. 2021, 16, 084051. [Google Scholar] [CrossRef]
- Ananyeva, N.D.; Sushko, S.V.; Ivashchenko, K.V.; Vasenev, V.I. Soil Microbial Respiration in Subtaiga and Forest-Steppe Ecosystems of European Russia: Field and Laboratory Approaches. Eurasian Soil Sci. 2020, 53, 1492–1501. [Google Scholar] [CrossRef]
- Luo, S.; Liu, G.; Li, Z.; Hu, C.; Gong, L.; Wang, M.; Hu, H. Soil Respiration along an Altitudinal Gradient in a Subalpine Secondary Forest in China. IForest Biogeosci. For. 2015, 8, 526–532. [Google Scholar] [CrossRef]
- Zheng, X.; Zhang, Y.; Zhang, Y.; Cui, Y.; Wu, J.; Zhang, W.; Wang, D.; Zou, J. Interactions between Nitrogen and Phosphorus in Modulating Soil Respiration: A Meta-Analysis. Sci. Total Environ. 2023, 905, 167346. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Shang, H. Contribution of Soil Respiration to the Global Carbon Equation. J. Plant Physiol. 2016, 203, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Kuzyakov, Y. Sources of CO2 Efflux from Soil and Review of Partitioning Methods. Soil Biol. Biochem. 2006, 38, 425–448. [Google Scholar] [CrossRef]
- Zhang, J.; Li, Y.; Wang, J.; Chen, W.; Tian, D.; Niu, S. Different Responses of Soil Respiration and Its Components to Nitrogen and Phosphorus Addition in a Subtropical Secondary Forest. For. Ecosyst. 2021, 8, 37. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Contrast | p Value | Sig. | ||||
|---|---|---|---|---|---|---|
| Soil temperature | vegetation/year | dry, 2016 | - | dry, 2017 | 2.765609 × 10−2 | * |
| mean/sd | 14.3/3.42 | 13.2/4.58 | ||||
| vegetation/year | dry, 2016 | - | green, 2016 | 2.267665 × 10−9 | *** | |
| mean/sd | 14.3/3.42 | 12.1/3.08 | ||||
| vegetation/year | dry, 2017 | - | green, 2017 | 1.307363 × 10−2 | * | |
| mean/sd | 13.2/4.58 | 12.1/5.04 | ||||
| vegetation/year | green, 2016 | - | green, 2017 | 6.340607 × 10−1 | ns | |
| mean/sd | 12.1/3.08 | 12.1/5.04 | ||||
| Soil moisture | vegetation/year | dry, 2016 | - | dry, 2017 | 4.890919 × 10−2 | * |
| mean/sd | 35.2/13.1 | 40.6/17.2 | ||||
| vegetation/year | dry, 2016 | - | green, 2016 | 4.949577 × 10−7 | *** | |
| mean/sd | 35.2/13.1 | 27.0/9.05 | ||||
| vegetation/year | dry, 2017 | - | green, 2017 | 2.894861 × 10−6 | *** | |
| mean/sd | 40.6/17.2 | 29.7/10.7 | ||||
| vegetation/year | green, 2016 | - | green, 2017 | 4.890919 × 10−2 | * | |
| mean/sd | 27.0/9.05 | 29.7/10.7 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tomes, J.; Fleischer, P., Jr.; Kubov, M.; Fleischer, P. Soil Respiration after Bark Beetle Infestation along a Vertical Transect in Mountain Spruce Forest. Forests 2024, 15, 611. https://doi.org/10.3390/f15040611
Tomes J, Fleischer P Jr., Kubov M, Fleischer P. Soil Respiration after Bark Beetle Infestation along a Vertical Transect in Mountain Spruce Forest. Forests. 2024; 15(4):611. https://doi.org/10.3390/f15040611
Chicago/Turabian StyleTomes, Jakub, Peter Fleischer, Jr., Martin Kubov, and Peter Fleischer. 2024. "Soil Respiration after Bark Beetle Infestation along a Vertical Transect in Mountain Spruce Forest" Forests 15, no. 4: 611. https://doi.org/10.3390/f15040611
APA StyleTomes, J., Fleischer, P., Jr., Kubov, M., & Fleischer, P. (2024). Soil Respiration after Bark Beetle Infestation along a Vertical Transect in Mountain Spruce Forest. Forests, 15(4), 611. https://doi.org/10.3390/f15040611