1. Introduction
A plethora of
Fusarium species are acknowledged as causative agents of severe forest diseases, posing significant challenges to forest nurseries and plantations worldwide [
1,
2]. Among these,
Fusarium circinatum Nirenberg & O’Donnell stands out as a prominent instigator of damping-off and root rot in pine seedling nurseries, leading to substantial damage to conifers [
3] and posing a significant threat. This fungal pathogen is particularly notorious for its capacity to inflict widespread and destructive losses on numerous conifer species [
4], notably through the induction of pitch canker (PPC) of susceptible tree species [
5].
Originally believed to have originated in North America,
F. circinatum has spread to Europe, with reported occurrences in Portugal, Spain, France, and Italy across various susceptible
Pinus species [
5]. The fungus has only been reported to infect one known host outside the genus
Pinus, which is
Pseudotsuga menziesii (Douglas fir), observed to be infected under field conditions in California [
6]. Currently, the disease in Europe has only been reported from the Iberian Peninsula, with a high likelihood of further entry and spread within the EU [
4]. Anticipated alterations in pine pitch canker distribution, influenced by rapid global climate change and diverse pathways for pathogen entry, underscore the need for proactive measures. The extent and severity of PPC are also expected to depend on host factors and other forest and climatic conditions. Furthermore, the latent phase of infection complicates early identification, posing a significant challenge for timely intervention and containment measures [
1]. This latent phase increases the risk of PPC spreading from nurseries to forests, emphasizing the importance of identifying vulnerable and tolerant species or provenances to mitigate its rapid spread into disease-free European countries. While natural infections primarily occur through wounds, the pathogen also has the ability to colonize the roots of mature trees without causing damage to surrounding tissues, thereby enabling plants to avoid exhibiting symptoms [
7].
F. circinatum exhibits a considerable capacity to establish itself in various parts of Europe. According to the European Food Safety Authority (EFSA), the impact of PPC in regions conducive to its proliferation could be substantial [
8]. PPC affects over 60 species of
Pinus, including
P. sylvestris L., across all stages of tree development [
7]. In adult trees, the disease manifests through symptoms such as pitch flow, the formation of resinous cankers, and branch and crown dieback. In nurseries, symptoms include tip dieback and damping-off of seedlings [
7].
The fungus causes resin-soaked cankers on roots and root collars of mature trees, while also being present internally in pine seeds or as a surface contaminant [
7]. Although ascospores are presumed to have a minimal role in infection, the pathogen primarily spreads through macroconidia and microconidia carried by rain, wind, or insects [
4,
7]. While bark-feeding insects may contribute to infection through wounds, recent studies suggest
F. circinatum can colonize seedlings and tree branches without wounds [
7]. Predicting the environmental suitability for PPC infection is complex due to the influence of various factors on both
F. circinatum and its hosts and vectors. High sporulation occurs under conditions of high relative humidity, either through precipitation or fog, coupled with cooler temperatures (15–25 °C), while sporulation diminishes as temperatures rise, and ceases when minimum temperatures approach zero [
8]. These patterns suggest that the disease may proliferate more readily in coastal, mild, wet climates rather than in mountainous or continental climates with extreme temperature fluctuations.
Numerous studies have delved into the infection mechanisms and virulence of
F. circinatum. Following conidial germination, the pathogen radially colonizes the host stem, progressing towards the pith, potentially facilitated by the release of cell wall-degrading enzymes to extract nutrients from the host tissues [
9]. Previous studies have also explored variation in susceptibility within certain
Pinus families, clones, and hybrids [
5], revealing significant differences in tolerance to
F. circinatum among various pine species and provenances. Various phenotypic and molecular characteristics have been linked to
F. circinatum pathogenicity or virulence [
7]. Our previous study also presented the first investigation into the susceptibility of Polish provenances of Scots pine (
Pinus sylvestris) to
F. circinatum infection, revealing high susceptibility among all tested pines and highlighting the potential risk of establishment posed by this invasive pathogen in Poland [
10]. These findings are of particular interest for forestry in Europe, North America, and South Africa [
1,
7].
Against this backdrop, our study aims to investigate differences in the pathogenicity of F. circinatum in seeds and seedlings of Pinus sylvestris provenances from Poland, Lithuania, and Ukraine. This research seeks to facilitate the rapid selection of PPC-tolerant or less susceptible individuals, offering valuable insights for subsequent tree breeding trials.
2. Materials and Methods
2.1. Fungal Isolate and Plant Material for Testing
The
F. circinatum isolate (FcCa6) utilized in this study belongs to mating type 2 (MAT-2) and was isolated from an infected tree of
Pinus radiata D. Don. located in Comillas (Cantabria, Northern Spain; GPS: 4°17′17.706″ W; 43°20′05.033″ N) [
11]. The isolate was obtained through the quarantine service, and all testing procedures were conducted at the Research Centre of Quarantine, Invasive, and Genetically Modified Organisms, Institute of Plant Protection (Poland). The
F. circinatum inoculum was produced on potato dextrose agar (PDA) at 25 °C.
To evaluate the susceptibility of
P. sylvestris within its native range, trees were germinated from seeds representing populations in various provenances in Poland, Ukraine, and Lithuania. These seeds were sourced from national gene bank collections (
Table 1).
The selected forest stands in Poland originated from six commercial forests spanning four major regions: the central region of the country (Łochów), the eastern border near Ukraine (Józefów), the northern region (Woziwoda) within Bory Tucholskie, and the northwest (Smolarz). These provenances were chosen for their representation of typical pine forests that thrive on post-glacial, poor sandy soils. Despite these conditions, they exhibit robust growth parameters and are emblematic of Polish forests. The choice of Ukrainian (Makariv and Manevychy) and Lithuanian (Šlienava) provenances was primarily justified by their geographic proximity to Poland, facilitating a relevant comparison in terms of environmental conditions and potential pathogen spread. Additionally, these forests in Lithuania and Ukraine represent typical forested areas in their respective countries, ensuring the inclusion of diverse and representative ecosystems in the study.
Seeds from selected mother trees with confirmed optimal phenotypic characteristics were used for the study, sourced from the Forest Gene Bank in Kostrzyca, Nature Research Centre in Lithuania, and Ukrainian Research Institute of Forestry and Forest Melioration. These seeds, of consistent age and quality, were obtained from a separate batch than those used in a previous study.
This assessment aimed to establish a direct correlation with existing data for the trees, including high viability, tolerance to stress, and resistance to pine diseases. Each seed lot included three hundred seeds that were divided, with one portion (two hundred seeds) used in assays conducted at the seedling emergence stage (Experiment 1), as described below, and another portion (one hundred seeds) grown for use in an inoculation assay conducted at four months of age (Experiment 2).
Cultures of FcCa6 were grown on potato dextrose agar (PDA) at 24 °C for 5 days. Spores were then harvested by flooding plates with 25 mL of sterile water and scraping the colonies with sterile glass beads. The resulting suspension was filtered through double layers of sterile cheesecloth. The spore density in the suspension was estimated using a hemacytometer, and subsequently diluted with sterile water to obtain spore densities of 5000 spores per mL.
2.2. In Planta Pathogenicity Test
Each trial encompassed distinct sets of treatments. Stratified seeds of P. sylvestris from diverse provenances in Poland, Ukraine, and Lithuania were enclosed in new mesh bags and immersed in tap water to reduce seed contaminants. The absence of F. circinatum in Poland, Lithuania, and Ukraine meant that this pathogen should be absent. In preparation for planting, all seeds underwent surface sterilization in 30% H2O2 for 30 min, followed by ten rinses with sterile distilled water.
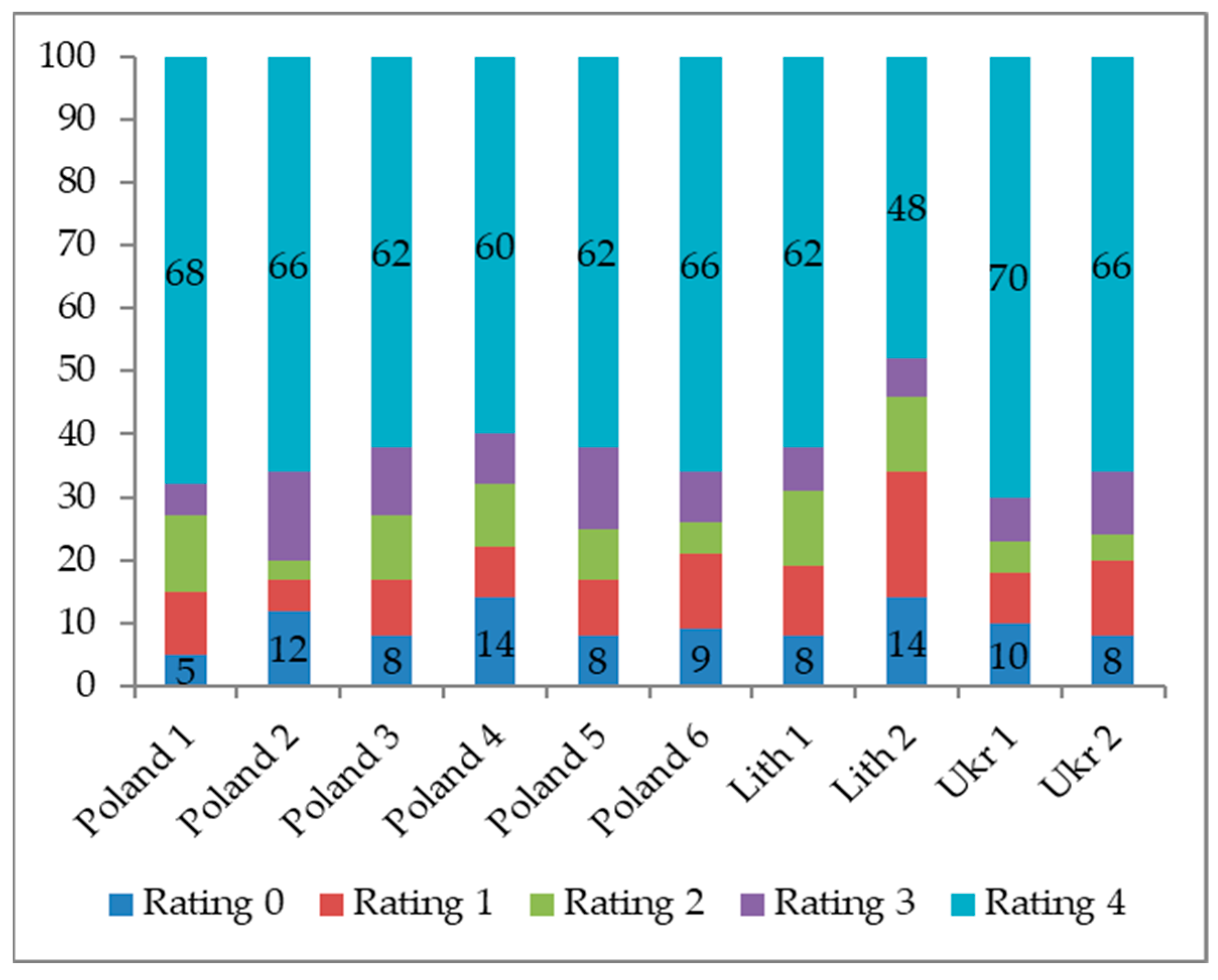
Experiment 1: Two hundred seeds from each trial were germinated at approximately 22 °C on moistened, sterile Whatman No. 3 filter paper within sterile Petri dishes (150 × 20 mm) without any artificial lighting. All tested seeds were inoculated by pipetting 200 µL of a spore solution into Petri dishes and subsequently incubating them at 23–24 °C. Conidial suspensions were applied to each Petri dish without direct contact with the seeds. Control groups for each provenance were inoculated with 200 µL of sterile water. Seed germination progress was monitored daily, with the threshold that seeds were germinated when their primary root reached a length of at least 5 mm. The evaluation period for the
F. circinatum impact on germination lasted 30 days for each trial, during which seedling mortality was recorded. Diseased seedlings were excised, and roots were meticulously washed and placed directly on PDA for re-isolation of the inoculated isolates [
12]. After 30 days, roots of surviving seedlings were scrutinized for length and condition. Root samples from surviving seedlings were also analyzed for evidence of infection by the inoculated isolates, as previously described. Following the examination of seedlings for disease symptoms, re-isolations onto MEA were performed for all inoculated and control seedlings (
Figure 1).
Experiment 2: One hundred seeds per treatment were individually sown in 200 mL cylindrical pots containing a sterilized mixture of peat and vermiculite (1:1,
v/
v). Two seeds per pot were sown at a depth of approximately 3 cm. Emerging seedlings were cultivated in the greenhouse at 22–23 °C with a 14/10 h light/dark photoperiod and watering twice a week. If both seeds germinated in the same pot, the second seedlings were removed within 1–2 days of emergence [
13].
Subsequently, five-month-old seedlings for each treatment were inoculated by pipetting 1 mL of a spore solution of
F. circinatum (5000 spores mL
−1) onto the soil surface or irrigated with 1 mL of sterile water (control). Morphological assessments of all pine seedlings were conducted every three days to assess the severity of typical symptoms of pitch canker. Seedlings were rated for symptom development on a 0–4-point scale: 0 = no symptoms, 1 = desiccation of needles, 2 = discoloration of needles, 3 = wilting of needles, and 4 = seedling death [
14,
15]. The final disease evaluation was at 180 days post-inoculation. Seedling heights were measured, and half of the seedlings were individually collected for re-isolation [
12]. Re-isolation from both symptomatic and asymptomatic seedlings involved surface sterilization in a solution of 1.5% sodium hypochlorite for up to 1 min and rinsing with sterile distilled water. These samples were then placed onpotato dextrose agar (PDA) supplemented with 100 mg/mL streptomycin sulphate. For symptomatic and asymptomatic seedlings, isolation was performed from washed sections below the collar and root. Plates were then incubated at room temperature with day/night light alternation, and typical
F. circinatum colonies were transferred to PDA for morphological confirmation and further molecular identification, as described in
Section 2.3 [
11,
16].
The remaining root samples were placed in 2 mL Eppendorf tubes for DNA extraction. The collected samples were homogenized by cutting with surface-sterilized scissors to 10 mm lengths and shaking in 600 mL of extraction buffer, followed by the addition of two glass beads to every Eppendorf tube. Finally, DNA extraction and conventional PCR tests were conducted on all samples to detect
F. circinatum using prescribed tests and species-specific primers (CIRC1A and CIRC4A, as described in
Section 2.3) (
Figure 1).
In this study, the methodology incorporated insights gained from a previous investigation conducted in 2018 [
10]. As a result, adjustments were made regarding the age of the seedlings tested. While two-month-old seedlings proved to be overly susceptible and twelve-month-old seedlings too tolerant, we opted to test five-month-old seedlings in Experiment 2 with ratings at six months. This was a compromise compared to the previous study from 2018, where susceptibility was assessed at 60 and 120 days post-inoculation [
10].
2.3. Confirmation of Fusarium circinatum by PCR
Fresh or frozen samples (roots and fungal mycelium) were freeze-dried at −85 °C for 72 h using a ScanVac freeze drier (Labogene, Lillerød, Denmark). Lyophilized samples were crushed, and samples up to 700 mg dry weight were homogenized in a high-speed homogenizer machine (Bertin instruments, Montigny-leBretonneux, France). The extraction and purification procedure followed the protocols of the NucleoSpin® Plant II Midi kit (Macherey-Nagel, Düren, Germany). DNA quantification and quality control of the DNA samples were analyzed spectrophotometrically with a NanoDrop™ One spectrophotometer (Thermo Scientific, Rochester, NY, USA) and adjusted to 10 ng/μL.
DNA extracts were subjected to PCR using species-specific primers CIRC1A and CIRC4A to detect
F. circinatum in samples. PCR was performed in a 20 μL PCR reaction using 1× PCR buffer supplied with the DNA polymerase, 0.25 mM of each dNTP, 2 mM MgCl
2, 0.5 μM of each primer, 0.05 U/μL DNA polymerase and 6.0 μL of template DNA (approximately 1 ng/μL) [
10,
17].
The intergenic spacer region (IGS) of the rDNA region was amplified using an Applied Biosystems 2720 thermal cycler (Foster City, CA, USA). The PCR cycling program was set as follows: first step at 95 °C for 3 min; followed by 40 cycles of 30 s at 95 °C, annealing at 64 °C for 55 s and 50 s at 72 °C; and final extension at 72 °C for 12 min [
10].
Final PCR products were analyzed using gel electrophoresis on a 1% agarose gel that included a lane of Nancy-520 (Sigma–Aldrich, Merck Group, Burlington, MA, USA) DNA Ladders and viewed with an UV illuminator. After PCR testing with the CIRC1A/4A primers, samples that generated a 146 bp fragment were classified as positive and with the presence of F. circinatum confirmed. A DNA template containing DNA from an F. circinatum isolate that yielded a 146 bp fragment after amplification with CIRC1A and CIRC4A primers served as a positive control. A negative control (purified water) and a false control (a DNA template containing DNA from isolates of F. poae and F. oxysporum from a previous study) were also used to confirm the specificity of the F. circinatum primers and to avoid bias from other Fusarium species.
2.4. Statistical Analyses
Average decline and mortality for seedlings were compared using one-way analysis of variance (ANOVA); comparisons were made between provenances; and significant means (
p-value = 0.05) were separated using post hoc test Fisher’s Least Significant Difference (LSD) test in Statistica software (STATISTICA
® 7.0 (StatSoft, Inc., Tulsa, OK, USA)). Welch’s F-test or the unequal variances test was used to test the hypothesis that all three provenances have equal means when the samples have unequal variances and/or unequal sample sizes. The survival analysis was performed based on the nonparametric estimator Kaplan–Meier with the “Survival” package to test final mortality. These analyses were performed using Survival 3.5-8 in R 4.1.1 (
https://www.r-project.org, accessed on 20 December 2023).
4. Discussion
Numerous studies have reported significant variability in the susceptibility of conifers to pitch canker, including
Pinus sylvestris, emphasizing the high or moderate virulence of the pathogen for both seedlings and mature plants [
5,
18,
19]. Given the absence of cost-effective and efficient treatments for mitigating pitch canker, genetic resistance emerges as the primary strategy to address potential PPC damage. The advancements in genetic resistance integrated into breeding programs can mitigate the risk of infection by different fungal pathogens, based on field observations of tested resistant phenotypes [
20,
21,
22]. While efforts to screen for tolerance to
F. circinatum have been undertaken in many locations, such as the USA, Africa, and Europe, there are currently no Scots pine breeding initiatives producing seeds from populations with confirmed pitch canker tolerance [
15,
23,
24]. This is mainly because field data have been found consistent for screening for pitch canker resistance.
Several studies have also highlighted variations in tolerance among
Pinus species in different taxonomic sections, as well as differences among species and families within these sections [
9,
15,
20,
25]. These studies, along with others involving inoculations, have demonstrated that varieties of
P. sylvestris are susceptible to damage but there exists a broad spectrum of the resistance level. Variations in susceptibility to pitch canker have also been observed among pine provenances in research conducted across various countries [
14,
26]. Preliminary screens of Polish provenances of Scots pine [
10] revealed the absence of PPC symptoms in some seedlings from two provenances after inoculation with
F. circinatum, similar to findings observed in certain Romanian and Austrian provenances [
5,
26]. The initial investigation of Polish pine provenances spanned the four-month post-inoculation period [
10]; therefore, in present study, the incubation period was extended to six months to explore potential resistance against
F. circinatum, considering the variability in susceptibility observed among the Polish provenances [
10] and new data from neighboring Ukrainian and Lithuanian provenances. While only two provenances were utilized for Lithuania and Ukraine, six sites were selected for Poland to account for risks of potential pathogen spread. The methodology incorporated lessons from a previous study [
10], with adjustments made regarding seedling age and the timing of susceptibility data collection. Despite promising viability post-inoculation, challenges in the survival analysis and the risk of latent infections emphasize the need for continued observation to accurately assess susceptibility. Certain provenances from Poland and Lithuania have not been deemed completely resistant to pitch canker caused by
F. circinatum. The inability of
F. circinatum to induce disease in certain seedlings may be attributable to various factors, such as individual antifungal activity or the upregulation of proteins associated with carbohydrate and amino acid metabolism, as well as secondary metabolism. These molecular responses could be interpreted as a strategy to compensate for the energy deficit incurred during infection and to withstand the pathogen’s impact. This study highlights the effectiveness of such an approach in preparing foresters for emerging diseases. Despite promising viability post-inoculation, challenges in the survival analysis and the risk of latent infection underscore the importance of ongoing observation to accurately assess susceptibility. The high susceptibility of
P. sylvestris underscores the significant threat posed by the introduction of
F. circinatum, especially in nursery facilities conducive to its global spread.
In the present study, F. circinatum was re-isolated from all inoculated pine seedlings, despite the apparent resistance of seedlings to F. circinatum infection. The frequency of re-isolation varied among and within provenances, with no variation observed in re-isolation between certain sites. Although certain seedlings appeared resistant to F. circinatum in this study, the fungus was still re-isolated from them, suggesting that some degree of growth into host tissues may occur without necessarily resulting in decline and mortality. Therefore, while certain provenances may exhibit resistance to F. circinatum infection, the presence of the fungus within host tissues indicates the potential for colonization without visible disease symptoms. Further research is needed to elucidate the mechanisms underlying the resistance of some provenances to pitch canker and the dynamics of F. circinatum colonization in host species, especially genotypes that are infected but apparently asymptomatic.
Contrary to the traditional view of pitch canker inducing damping-off symptoms, our study underscores the severe negative impact of F. circinatum on the decline, root inhibition, and post-emergent mortality of P. sylvestris seedlings. Surviving seedlings, however, did not exhibit a subsequent increase in root and shoot biomass or growth even six months post-inoculation compared to the control group. Weakened seedlings showed reduced root development without an enhancement of resistance to stem infections. In contrast, asymptomatic seedlings (score 0, showing no symptoms of wilt or damping-off) displayed comparable root and height parameters to the control group. This suggests that while F. circinatum may not cause the death of certain seedlings, it might even induce resistance, potentially influencing their field survival and subsequent forest dynamics.
Climate is also considered a limiting factor for the spread of
F. circinatum in Europe [
7,
26]. Colder climates are considered restrictive, although the climatic change scenarios of the potential distribution of PPC in Europe includes the Netherlands and Denmark [
26,
27,
28]. Climatic constraints are less restrictive for
F. circinatum inducing massive damping-off in forest nurseries, particularly under protected cultivation. Moreover, any international trade in live plant material further increases the risk of pathogen dissemination to disease-free regions of Europe where Scots pine plantations and native forests are present. Our results align with the emerging concept that high-pathogenicity agents such as
F. circinatum may serve varied ecological functions extending beyond the immediate impacts of the disease [
29]. Nevertheless, our findings unequivocally indicate the susceptibility of the majority of
Pinus sylvestris provenances from Poland, Lithuania, and Ukraine to
F. circinatum, with the pathogen capable of adversely affecting and killing pine seedlings. However, some provenances and seedlings may exhibit potential resistance.
The dearth of data regarding resistance to
F. circinatum constrains our capacity to strategically deploy resources and enhance tolerance, which has not been a primary focus for breeding populations of
P. sylvestris. As certain groups have explored the potential benefits of employing genomic tools to characterize pitch canker tolerance in other pine species, these analyses could be effectively integrated into
P. sylvestris breeding programs to enhance tolerance [
25,
30,
31].
Despite these observations, it is crucial to recognize that most tests and experiments involving invasive quarantine pathogens like
F. circinatum, which rely on greenhouse inoculation data, require further validation. In both nursery and stand environments, multiple pathogens affecting roots, collars, foliage, and vasculature may coexist with
F. circinatum [
8]. While our data are preliminary, it is conceivable that the results could be influenced by volatiles or other compounds in plants or soil that enhance seedling or tree growth [
29,
30]. Therefore, considering the unique characteristics of Lithuanian provenance, along with the inherent resistance observed in certain seedlings from two Polish provenances and the variation in susceptibility observed in previous studies, our findings imply potential considerations for future genetic breeding programs of
P. sylvestris. In future investigations, expanding the scope to include more varieties from other Eastern European countries would be beneficial. The high susceptibility of
P. sylvestris underscores the potential threat of
F. circinatum establishment in Eastern and Northern Europe, underscoring the serious risks associated with its introduction to Poland, Ukraine, and Lithuania.


 ,
, 






{kind=link}
{kind=link}
{kind=link}