Enrichment of Logging Gaps with a High Conservation Value Species (Pericopsis elata) in a Central African Moist Forest

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Studied Species

2.3. Enrichment of Logging Gaps
2.4. Data Analysis
2.4.1. Differences between Sites
2.4.2. Modeling Seedling Performance
logit(πij | bj) = α + βc + θs + bj
with bj ~ Gaussian(0,σ²b)
with bj ~ Gaussian(0,σ²b)
with εij ~ Gaussian(0,σ²ε)
3. Results
3.1. Seedling Performance in Control Gaps

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Site 1 | Site 2 | Site 3 | Total | ||||
|---|---|---|---|---|---|---|---|---|
| Mean | S | Mean | S | Mean | S | Mean | S | |
| Mortality | ||||||||
| Mortality after 5 years (%) | 25.8 | 14.8 | 23.3 | 15.3 | 67.2 | 21.8 | 38.8 | 26.2 |
| Annualized mortality rate (% yearr−1) | 6.0 | 3.9 | 5.4 | 3.9 | 22.9 | 14.2 | 11.4 | 11.5 |
| Growth | ||||||||
| Planted height (cm) | 75 (n = 33) | 23 | 59 (n = 30) | 14 | 54 (n = 32) | 14 | 63 (n = 95) | 20 |
| 5 years height (cm) | 331 (n = 24) | 133 | 296 (n = 23) | 101 | 169 (n = 10) | 76 | 288 (n = 57) | 125 |
| Height increment (cm·year−1) | 50.94 | 26.00 | 47.18 | 19.73 | 24.46 | 13.28 | 44.78 | 23.48 |
| Diameter increment (mm·year−1) | 2.86 | 1.33 | 3.32 | 1.12 | 1.30 | 0.85 | 2.77 | 1.36 |
| RGR in height (cm·cm−1·year−1) | 0.28 | 0.10 | 0.31 | 0.09 | 0.24 | 0.06 | 0.29 | 0.09 |
| RGR in diameter (mm·mm−1·year−1) | 0.23 | 0.10 | 0.24 | 0.06 | 0.12 | 0.06 | 0.21 | 0.09 |
| Gap characteristics | ||||||||
| Age (mth) | 6.8 | 0 | 6.6 | 1.9 | 10.4 | 0.32 | 7.9 | 2.1 |
| Area (m2) | 121 | 25 | 122 | 25 | 191 | 39 | 145 | 44 |
| Number of planted seedlings in a gap | 11 | 2.7 | 10 | 0 | 10.7 | 1.2 | 10.6 | 1.5 |
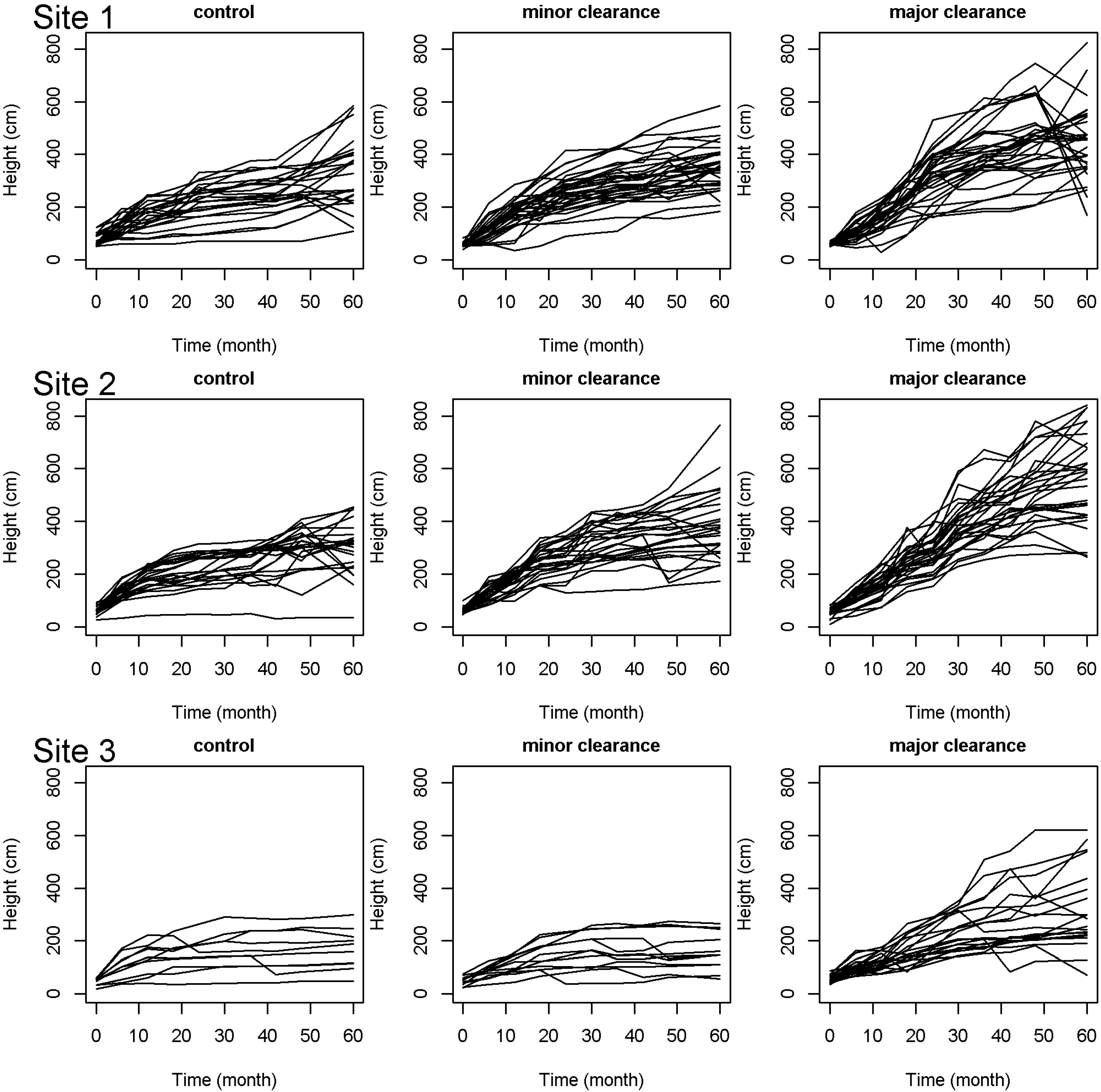
3.2. Effect of Clearance

| Variable | Control | Minor C. | Major C. | |||
|---|---|---|---|---|---|---|
| Mean | S | Mean | S | Mean | S | |
| Mortality | ||||||
| Mortality after 5 years (%) | 38.8 | 26.2 | 26.9 | 25.1 | 14.4 | 20.1 |
| Annualized mortality rate (% year−1) | 11.4 | 11.5 | 7.1 | 7.5 | 3.5 | 5 |
| Growth | ||||||
| Planted height (cm) | 63 (n = 95) | 20 | 55 (n = 96) | 13 | 56 (n = 94) | 13 |
| 5 years height (cm) | 288 (n = 57) | 125 | 331 (n = 70) | 130 | 458 (n = 81) | 179 |
| Height increment (cm year−1) | 44.78 | 23.48 | 54.82 | 25.67 | 80.32 | 36.05 |
| Diameter increment (mm year−1) | 2.77 | 1.36 | 4.23 | 2.24 | 8.04 | 4.05 |
| RGR in height (cm cm−1 year−1) | 0.29 | 0.09 | 0.34 | 0.09 | 0.41 | 0.11 |
| RGR in diameter (mm mm−1 year−1) | 0.21 | 0.09 | 0.29 | 0.10 | 0.40 | 0.12 |
| Gap characteristics | ||||||
| Age (mth) | 7.9 | 2.1 | 8.3 | 1.6 | 7.9 | 2.0 |
| Area (m2) | 145 | 44 | 166 | 59 | 149 | 42 |
| Number of planted seedlings in a gap | 10.6 | 1.5 | 10.7 | 3.0 | 10.0 | 3.3 |
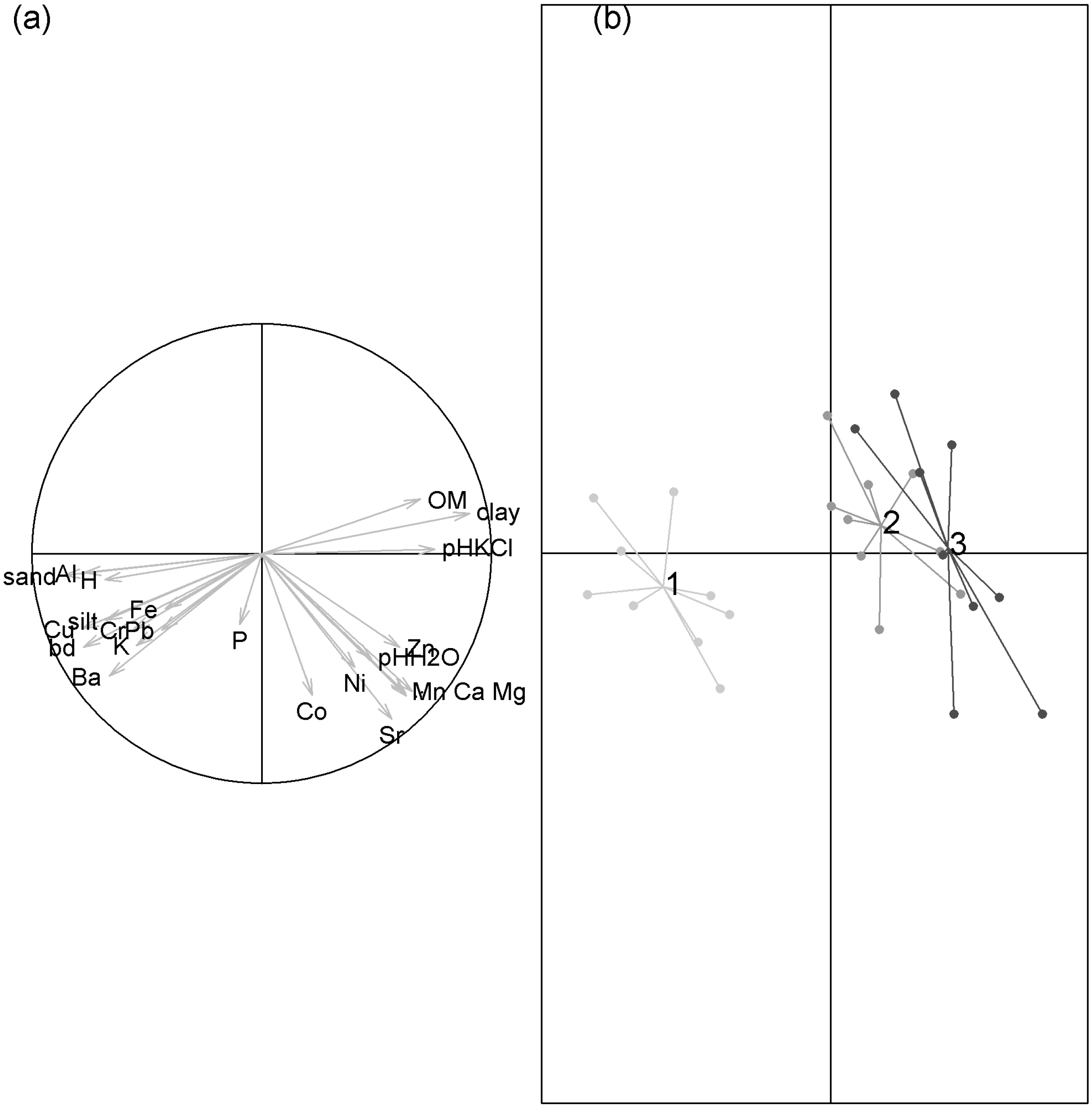
3.3. Between Site Differences

4. Discussion
4.1. The Seedlings of P. elata Performed Well in Logging Gaps
4.2. Clearance Increases the Performance of P. elata Seedlings
4.3. Determinants of between Site Differences
5. Conclusions
Acknowledgments
Author Contributions
Appendix



| Soil variable | Site 1 | Site 2 | Site 3 | |
|---|---|---|---|---|
| % clay | mean | 42.97 a | 61.89 b | 67.11 b |
| sd | 7.15 | 2.93 | 4.98 | |
| % sand | mean | 43.61 a | 33.52 a b | 25.71 b |
| sd | 6.74 | 3.58 | 4.12 | |
| % silt | mean | 13.46 a | 4.59 b | 7.16 b |
| sd | 2.62 | 1.88 | 1.90 | |
| % organic matter | mean | 5.18 a | 6.72b | 7.33 b |
| sd | 0.76 | 1.38 | 0.84 | |
| pH H2O | mean | 3.99 a | 3.98 a | 4.11 a |
| sd | 0.09 | 0.12 | 0.13 | |
| pH KCl | mean | 3.87 a | 4.09 b | 4.13 b |
| sd | 0.11 | 0.21 | 0.10 | |
| Bulk density | mean | 1.21a | 1.02 b | 1.02b |
| (g/cm3) | sd | 0.05 | 0.03 | 0.09 |
| P | mean | 2.48 a | 2.90 a | 2.08 a |
| (μg/g) | sd | 0.52 | 1.06 | 0.52 |
| K | mean | 0.14 a | 0.08 b | 0.10 a b |
| (cmolc/kg) | sd | 0.04 | 0.02 | 0.02 |
| Ca | mean | 0.32 a | 0.41 a | 0.64a |
| (cmolc/kg) | sd | 0.21 | 0.17 | 0.34 |
| Mg | mean | 0.18 a | 0.23 a | 0.26 a |
| (cmolc/kg) | sd | 0.06 | 0.08 | 0.09 |
| Al | mean | 3.03 a | 1.60 b | 1.26 b |
| (cmolc/kg) | sd | 1.07 | 0.47 | 0.61 |
| H | mean | 0.86 a | 0.40 b | 0.35 b |
| (cmolc/kg) | sd | 0.30 | 0.22 | 0.27 |
Conflicts of Interest
References
- Sist, P.; Gourlet-Fleury, S.; Putz, F.E. The impacts of selective logging: Questionable conclusions. BioScience 2012, 62, 786–786. [Google Scholar]
- Fargeot, C.; Forni, E.; Nasi, R. Réflexion sur l’aménagement des forêts de production dans le bassin du Congo. Bois For. Trop. 2004, 281, 19–34. [Google Scholar]
- Karsenty, A.; Gourlet-Fleury, S. Assessing sustainability of logging practices in the Congo Basin’s managed forests: The issue of commercial species recovery. Ecol. Soc. 2006, 11, 26. [Google Scholar]
- Gourlet-Fleury, S.; Mortier, F.; Fayolle, A.; Baya, F.; Ouédraogo, D.; Bénédet, F.; Picard, N. Tropical forest recovery from logging: A 24 year silvicultural experiment from Central Africa. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368. [Google Scholar] [CrossRef] [Green Version]
- Hawthorne, W.D. Ecological Profiles of Ghanaian Forest Trees; Oxford Forestry Institute, Department of Plant Sciences, University of Oxford: Oxford, UK, 1995. [Google Scholar]
- Doucet, J.L. L’alliance délicate de la gestion forestière et de la biodiversité dans les forêts du centre du Gabon. Ph.D. Thesis, Faculté Universitaire des Sciences agronomiques de Gembloux, Gembloux, Belgium, 2003. [Google Scholar]
- Makana, J.R.; Thomas, S.C. Effects of light gaps and litter removal on the seedling performance of six African timber species. Biotropica 2005, 37, 227–237. [Google Scholar] [CrossRef]
- Makana, J.R.; Thomas, S.C. Impacts of selective logging and agricultural clearing on forest structure, floristic composition and diversity, and timber tree regeneration in the Ituri Forest, Democratic Republic of Congo. Biodivers. Conserv. 2006, 15, 1375–1397. [Google Scholar] [CrossRef]
- Doucet, J.-L.; Kouadio, Y.L.; Monticelli, D.; Lejeune, P. Enrichment of logging gaps with moabi (Baillonella toxisperma Pierre) in a Central African rain forest. For. Ecol. Manag. 2009, 258, 2407–2415. [Google Scholar] [CrossRef]
- Brokaw, N.V.L. Gap-phase regeneration in a tropical forest. Ecology 1985, 66, 682–687. [Google Scholar] [CrossRef]
- Kyereh, B.; Swaine, M.D.; Thompson, J. Effect of light on the germination of forest trees in Ghana. J. Ecol. 1999, 87, 772–783. [Google Scholar] [CrossRef]
- Babaasa, D.; Eilu, G.; Kasangaki, A.; Bitariho, R.; McNeilage, A. Gap characteristics and regeneration in Bwindi Impenetrable National Park, Uganda. Afr. J. Ecol. 2004, 42, 217–224. [Google Scholar] [CrossRef]
- Keefe, K.; Alavalapati, J.A.A.; Pinheiro, C. Is enrichment planting worth its costs? A financial cost-benefit analysis. For. Policy Econ. 2012, 23, 10–16. [Google Scholar] [CrossRef]
- Schwartz, G.; Lopes, J.C.A.; Mohren, G.M.J.; Peña-Claros, M. Post-harvesting silvicultural treatments in logging gaps: A comparison between enrichment planting and tending of natural regeneration. For. Ecol. Manag. 2013, 293, 57–64. [Google Scholar] [CrossRef]
- Owona Ndongo, P.-A.; Peltier, R.; Linjouom, I.; Louppe, D.; Smektala, G.; Beligne, V.; Njoukam, R.; Tieche, B.; Temgoua, L. Plantations de bois d’oeuvre en zone équatoriale africaine : Cas de l’arboretum de l’Enef de Mbalmayo au sud du Cameroun. Bois For. Trop. 2009, 299, 37–48. [Google Scholar]
- Doucet, J.-L.; Ntchandi Otimbo, P.-A.; Boubady, A.-G. Assisting natural regeneration of Okoume populations in logging concessions? Bois For. Trop. 2004, 279, 59–71. [Google Scholar]
- Bosu, P.P.; Cobbinah, J.R.; Nichols, J.D.; Nkrumah, E.E.; Wagner, M.R. Survival and growth of mixed plantations of Milicia excelsa and Terminalia superba 9 years after planting in Ghana. For. Ecol. Manag. 2006, 233, 352–357. [Google Scholar] [CrossRef]
- Opuni-Frimpong, E.; Karnosky, D.F.; Storer, A.J.; Cobbinah, J.R. Silvicultural systems for plantation mahogany in Africa: Influences of canopy shade on tree growth and pest damage. For. Ecol. Manag. 2008, 255, 328–333. [Google Scholar] [CrossRef]
- African Regional Workshop. IUCN red List threat species version. Proceedings of the Conservation & Sustainable Management of Trees, Pericopsis elata, Harare, Zimbabwe, July 1996; Available online: http://www.iucnredlist.org (accessed on 1 October 2014).
- Bourland, N.; Kouadio, Y.L.; Lejeune, P.; Sonké, B.; Phillipart, J.; Daïnou, K.; Fétéké, F.; Doucet, J.-L. Ecology of Pericopsis elata (Fabaceae), an Endangered Timber Species in Southeastern Cameroon. Biotropica 2012, 44, 840–847. [Google Scholar] [CrossRef]
- Bourland, N.; Kouadio, Y.L.; Fétéké, F.; Lejeune, P.; Doucet, J.-L. Ecology and management of Pericopsis elata (Harms) Meeuwen (Fabaceae) populations: A review. Biotechnol. Agron. Soc. Environ. 2012, 16, 486–498. [Google Scholar]
- Schulze, M. Technical and financial analysis of enrichment planting in logging gaps as a potential component of forest management in the eastern Amazon. For. Ecol. Manag. 2008, 255, 866–879. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Letouzey, R. Carte Phytogéographique du Cameroun; Institut de la Recherche Agronomique: Paris, France, 1985. [Google Scholar]
- Fayolle, A.; Picard, N.; Doucet, J.-L.; Swaine, M.; Bayol, N.; Bénédet, F.; Gourlet-Fleury, S. A new insight in the structure, composition and functioning of central African moist forests. For. Ecol. Manag. 2014, 329, 195–205. [Google Scholar] [CrossRef]
- ATIBT. Association Technique Internationale des Bois Tropicaux Statistiques 2010. La Lettre de L’ATIBT 2012, 34, 1–45. [Google Scholar]
- Veenendaal, E.M.; Swaine, M.D.; Lecha, R.T.; Walsh, M.F.; Abebrese, I.K.; Owusu-Afriyie, K. Responses of West African forest tree seedlings to irradiance and soil fertility. Funct. Ecol. 1996, 10, 501–511. [Google Scholar] [CrossRef]
- Swaine, M.D.; Hall, J.B. The mosaic theory of forest regeneration and the determination of forest composition in Ghana. J. Trop. Ecol. 1988, 4, 253–269. [Google Scholar] [CrossRef]
- Boyemba, F. Écologie de Pericopsis elata (Harms) Van Meeuwen (Fabaceae), arbre de forêt tropicale africaine à répartition agrégée. Ph.D. Thesis, Université libre de Bruxelles, Brussels, Belgium, 2011. [Google Scholar]
- Hardy, O.J.; Born, C.; Budde, K.; Daïnou, K.; Dauby, G.; Duminil, J.; Ewédjé, E.-E.B.; Gomez, C.; Heuertz, M.; Koffi, G.K. Comparative phylogeography of African rain forest trees: A review of genetic signatures of vegetation history in the Guineo-Congolian region. Comptes Rendus Geosci. 2013, 345, 284–296. [Google Scholar] [CrossRef]
- Pansu, M.; Gautheyrou, J. Handbook of Soil Analysis: Mineralogical, Organic and Inorganic Methods; Springer: Berlin, Germany, 2007. [Google Scholar]
- Hollander, M.; Wolfe, D.A. Nonparametric Statistical Methods; John Wiley & Sons: New York, NY, USA, 1999. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Sheil, D.; Burslem, D.F.; Alder, D. The interpretation and misinterpretation of mortality rate measures. J. Ecol. 1995, 83, 331–333. [Google Scholar] [CrossRef]
- Hunt, R.; Thomas, B.; Murphy, D.J.; Murray, D. Growth analysis, individual plants. Encycl. Appl. Plant Sci. 2003, 2, 579–588. [Google Scholar]
- Nakagawa, S.; Schielzeth, H. A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods Ecol. Evol. 2013, 4, 133–142. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ouédraogo, D.-Y.; Fayolle, A.; Daïnou, K.; Demaret, C.; Bourland, N.; Lagoute, P.; Doucet, J.-L. Enrichment of Logging Gaps with a High Conservation Value Species (Pericopsis elata) in a Central African Moist Forest. Forests 2014, 5, 3031-3047. https://doi.org/10.3390/f5123031
Ouédraogo D-Y, Fayolle A, Daïnou K, Demaret C, Bourland N, Lagoute P, Doucet J-L. Enrichment of Logging Gaps with a High Conservation Value Species (Pericopsis elata) in a Central African Moist Forest. Forests. 2014; 5(12):3031-3047. https://doi.org/10.3390/f5123031
Chicago/Turabian StyleOuédraogo, Dakis-Yaoba, Adeline Fayolle, Kasso Daïnou, Charles Demaret, Nils Bourland, Paul Lagoute, and Jean-Louis Doucet. 2014. "Enrichment of Logging Gaps with a High Conservation Value Species (Pericopsis elata) in a Central African Moist Forest" Forests 5, no. 12: 3031-3047. https://doi.org/10.3390/f5123031
APA StyleOuédraogo, D. -Y., Fayolle, A., Daïnou, K., Demaret, C., Bourland, N., Lagoute, P., & Doucet, J. -L. (2014). Enrichment of Logging Gaps with a High Conservation Value Species (Pericopsis elata) in a Central African Moist Forest. Forests, 5(12), 3031-3047. https://doi.org/10.3390/f5123031