
Elevated Atmospheric CO2 Affects Ectomycorrhizal Species Abundance and Increases Sporocarp Production under Field Conditions


 ,
, 
Abstract
:1. Introduction
2. Material and Methods
2.1. The Bílý Kříž Site
2.2. The BangorFACE Facility
2.3. Root Sampling and Identification of Ectomycorrhizas
2.3.1. Bílý Kříž
2.3.2. BangorFACE
2.4. Collection of Sporocarps
2.5. Statistical Analysis
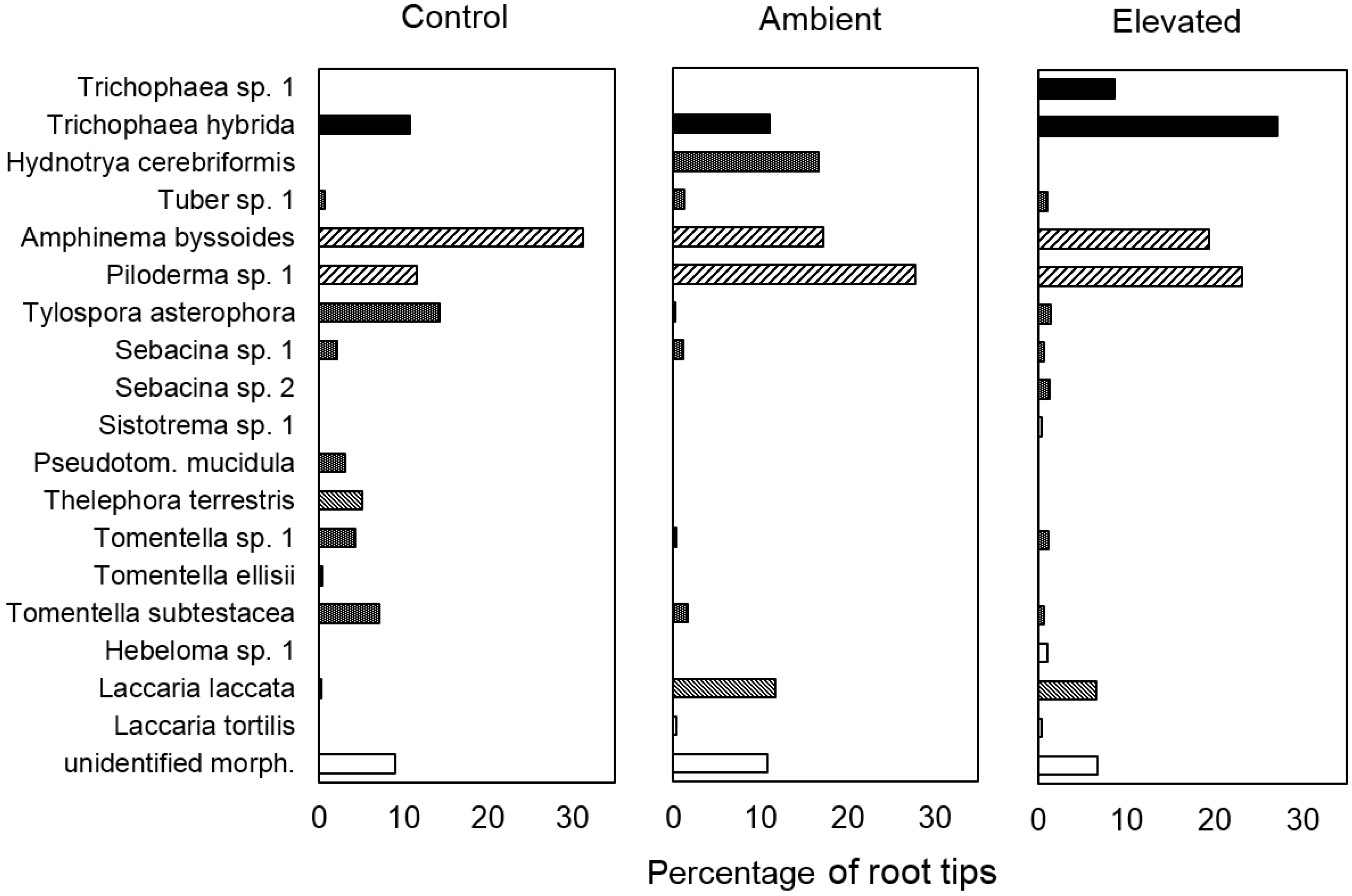
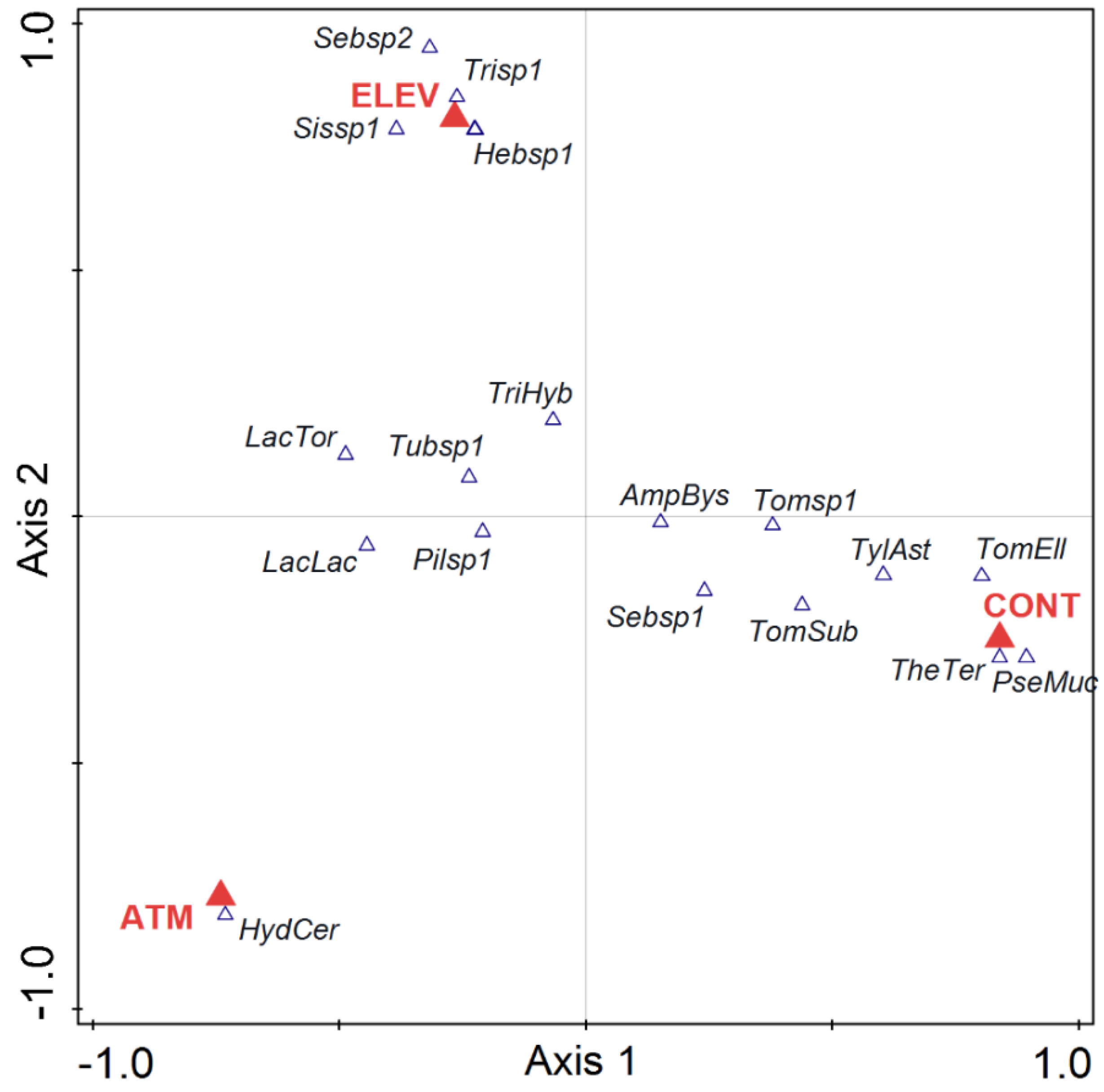
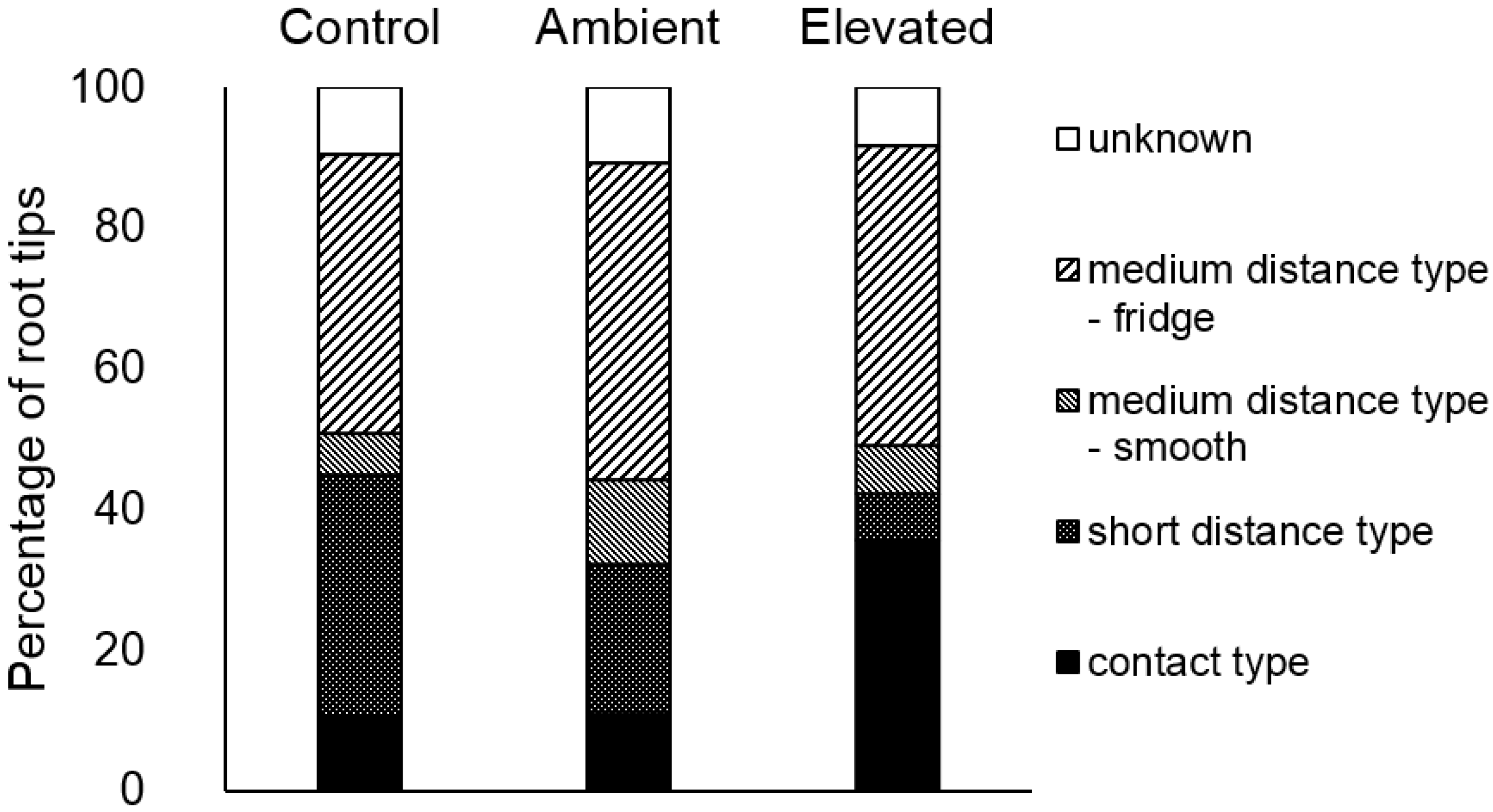
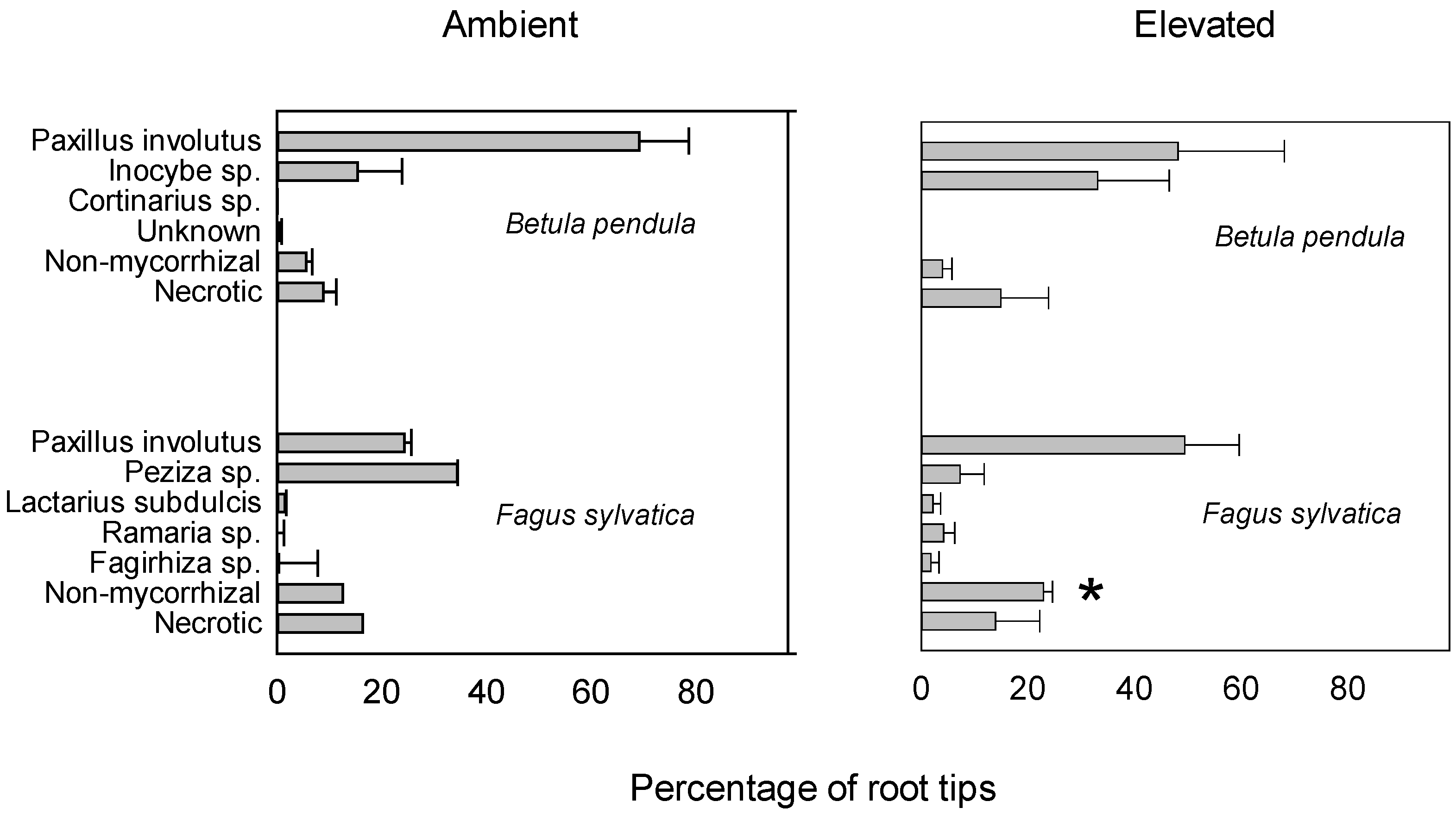
3. Results
 contact,
contact,  short distance,
short distance,  medium distance smooth,
medium distance smooth,  medium distance fridge,
medium distance fridge,  not known.
contact, short distance, medium distance smooth, medium distance fridge, not known.
not known.
contact, short distance, medium distance smooth, medium distance fridge, not known.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Sporocarp Biomass (g) | ||
|---|---|---|---|
| Ambient | Elevated CO2 | E/A | |
| Paxillus involutus | 29 ± 7 | 139 ± 61 | 4.8 |
| Inocybe geophylla | 0.04 ± 0.04 | 1.39 ± 1.05 | 32.0 |
| Peziza badia | 1.5 ± 0.6 | 8.9 ± 2.8 * | 6.1 |
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Taylor, A.F.S.; Martin, F.; Read, D.J. Fungal diversity in ectomycorrhizal communities of Norway spruce (Picea abies L. Karst.) and beech (Fagus sylvatica L.) along north-south transects in Europe. In Carbon and Nitrogen Cycling in European Forest Ecosystems; Schulze, E.-D., Ed.; Springer-Verlag: Berlin, Germany, 2000; Ecological studies 142; pp. 343–365. [Google Scholar]
- Lang, C.; Severn, J.; Polle, A. Host preferences and differential contributions of deciduous tree species shape mycorrhizal species richness in a mixed Central European forest. Mycorrhiza 2011, 21, 297–308. [Google Scholar] [CrossRef] [PubMed]
- Erland, S.; Jonsson, T.; Mahmood, S.; Finlay, R.D. Below-ground ectomycorrhizal community structure in two Picea abies forests in southern Sweden. Scand. J. For. Res. 1999, 14, 209–217. [Google Scholar] [CrossRef]
- Blom, J.M.; Vannini, A.; Vettraino, A.M.; Hale, M.D.; Godbold, D.L. Ectomycorrhizal community structure in a healthy and a Phytophthora infected chestnut (Castanea sativa Mill.) stand in central Italy. Mycorrhiza 2009, 20, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Dahlberg, A. Community ecology of ectomycorrhizal fungi: An advancing interdisciplinary field. New Phytol. 2001, 150, 555–562. [Google Scholar] [CrossRef]
- Agerer, R. Colour Atlas of Ectomycorrhizae; 1st–11th Delivery; Einhorn Verlag: Schwäbisch Gmünd, Germany, 1987–2006. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes-application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W.; Bolchacova, E.; Voigt, K.; Crous, P.W. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [PubMed]
- Fransson, P.M.A.; Taylor, A.F.S.; Finlay, R.D. Elevated atmospheric CO2 alters root symbiont community structure in forest trees. New Phytol. 2001, 152, 431–442. [Google Scholar] [CrossRef]
- Baxter, J.W.; Dighton, J. Ectomycorrhizal diversity alters growth and nutrient acquisition of grey birch (Betula populifolia) seedlings in host-symbiont culture conditions. New Phytol. 2001, 152, 139–149. [Google Scholar] [CrossRef]
- Cairney, J.W.G. Intraspecific physiological variation: Implications for understanding functional diversity in ectomycorrhizal fungi. Mycorrhiza 1999, 9, 125–135. [Google Scholar] [CrossRef]
- Leake, J.R.; Donnelly, D.P.; Saunders, E.M.; Boddy, L.; Read, D.J. Rates and quantities of carbon flux to ectomycorrhizal mycelium following 14C pulse labeling of Pinus sylvestris seedlings: Effects of litter patches and interaction with a wood-decomposer fungus. Tree Physiol. 2001, 21, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Courty, P.-E.; Pritsch, K.; Schloter, M.; Hartmann, A.; Garbaye, J. Activity profiling of ectomycorrhiza communities in two forest soils using multiple enzymatic tests. New Phytol. 2005, 167, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Hobbie, E.A.; Agerer, R. Nitrogen isotopes in ectomycorrhizal sporocarps correspond to belowground exploration types. Plant Soil 2010, 327, 71–83. [Google Scholar] [CrossRef]
- Bardgett, R.D.; van der Putten, W.H. Belowground biodiversity and ecosystem functioning. Nature 2014, 115, 505–511. [Google Scholar] [CrossRef]
- Agerer, R. Exploration types of ectomycorrhizae-proposal to classify ectomycorrhizal mycelial systems according to their patterns of differentiation and putative ecological importance. Mycorrhiza 2001, 11, 107–114. [Google Scholar] [CrossRef]
- Agerer, R.; Rambold, G. DEEMY—An Information System for Characterization and Determination of Ectomycorrhizae. Available online: http://www.deemy.de/Identification/NaviKey/index.html (accessed on 15 May 2013).
- Peay, K.G.; Kennedy, P.G.; Bruns, T.D. Rethinking ectomycorrhizal succession: Are root density and hyphal exploration types drivers of spatial and temporal zonation? Fungal Ecol. 2011, 4, 233–240. [Google Scholar] [CrossRef]
- Koide, R.T.; Christopher Fernandez, C.; Malcolm, G. Determining place and process: Functional traits of ectomycorrhizal fungi that affect both community structure and ecosystem function. New Phytol. 2013, 201, 433–439. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2013: The Physical Science Basis; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Norby, R.J.; deLucia, E.H.; Gielen, B.; Calfapietra, C.; Giardina, C.P.; King, J.S.; Ledford, J.; McCarthy, H.R.; Moore, D.J.P.; Ceulemans, R.; et al. Forest response to elevated CO2 is conserved across a broad range of productivity. Proc. Natl. Acad. Sci. USA 2005, 102, 18052–18056. [Google Scholar] [CrossRef] [PubMed]
- Fransson, P.M.A.; Taylor, A.F.S.; Finlay, R.D. Mycelial production, spread and root colonisation by ectomycorrhizal fungi Hebeloma crustuliniforme and Paxillus involutus under elevated atmospheric CO2. Mycorrhiza 2005, 15, 25–31. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, E.G.; Luxmoore, R.J.; Norby, R.J. Increases in mycorrhizal colonization and seedling growth in Pinus echinata and Quercus alba in an enriched CO2 atmosphere. Can. J. For. Res. 1987, 17, 878–883. [Google Scholar] [CrossRef]
- Treseder, K.K. A meta-analysis of mycorrhizal responses to nitrogen, phosphorus, and atmospheric CO2 in field studies. New Phytol. 2004, 164, 347–355. [Google Scholar] [CrossRef]
- Godbold, D.L.; Berntson, G.M.; Bazzaz, F.A. Growth and mycorrhizal colonization of three North American tree species under elevated atmospheric CO2. New Phytol. 1997, 137, 433–440. [Google Scholar] [CrossRef]
- Fransson, P.M.A.; Anderson, I.C.; Alexander, I.J. Does carbon partitioning in ectomycorrhizal pine seedlings under elevated CO2 vary with fungal species? Plant Soil 2007, 291, 323–333. [Google Scholar] [CrossRef]
- Rygiewicz, P.T.; Martin, K.J.; Tuininga, A.R. Morphotype community structure of ectomycorrhizas on Douglas fir (Pseudotsuga menziesii Mirb. Franco) seedlings grown under elevated atmospheric CO2 and temperature. Oecologia 2000, 124, 299–308. [Google Scholar] [CrossRef]
- Markkola, A.M.; Ohtonen, A.; Ahonen-Jonnarth, U.; Ohtonen, R. Scots pine responses to CO2 enrichment. 1. Ectomycorrhizal fungi and soil fauna. Environ. Pollut. 1996, 94, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Godbold, D.L.; Berntson, G.M. Elevated Atmospheric CO2 concentration leads to changes in ectomycorrhizal morphotype assemblage in Betula papyrifera. Tree Physiol. 1997, 17, 347–350. [Google Scholar] [CrossRef] [PubMed]
- Gorissen, A.; Kuyper, T.H. Fungal species-specific responses of ectomycorrhizal Scots pine (Pinus sylvestris) to elevated (CO2). New Phytol. 2000, 146, 163–168. [Google Scholar] [CrossRef]
- Rey, A.; Jarvis, P.G. Growth response of young birch trees (Betula pendula Roth) after four and a half years of CO2 exposure. Ann. Bot.-Lond. 1997, 80, 809–816. [Google Scholar] [CrossRef]
- Urban, O.; Janouš, D.; Pokorný, R.; Marková, I.; Pavelka, M.; Fojtík, Z.; Šprtová, M.; Kalina, J.; Marek, M.V. Glass domes with adjustable windows: A novel technique for exposing juvenile forest stands to elevated CO2 concentration. Photosynthetica 2001, 39, 395–401. [Google Scholar] [CrossRef]
- Vranova, V.; Skene, K.R.; Pokorny, R.; Rejsek, K.; Formanek, P. Change Research Centre AS CR, Ceske Budejovice, Czech Republic. Unpublished Work. 2014. [Google Scholar]
- Smith, A.R.; Lukac, M.; Hood, R.; Healey, J.R.; Miglietta, F.; Godbold, D. Elevated CO2 enrichment induces a differential biomass response in a mixed species temperate forest plantation. New Phytol. 2013, 198, 156–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, A.R.; Lukac, M.; Bambrick, M.; Miglietta, F.; Godbold, D.L. Tree species diversity interacts with elevated CO2 to induce a greater root system response. Glob. Change Biol. 2013, 19, 217–228. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Nikolcheva, L.G.; Bärlocher, F. Taxon-specific fungal primers reveal unexpectedly high diversity during leaf decomposition in a stream. Mycol. Prog. 2004, 3, 41–49. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef]
- Abarenkov, K.; Nilsson, R.H.; Larsson, K.H.; Alexander, I.J.; Eberhardt, U.; Erland, S.; Høiland, K.; Kjøller, R.; Larsson, E.; Pennanen, T.; et al. The UNITE database for molecular identification of fungi-recent updates and future perspectives. New Phytol. 2010, 186, 281–285. [Google Scholar] [CrossRef] [PubMed]
- Benson, D.A.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. “GenBank”. Nucleic Acids Res. 2011, 39, D32–D37. [Google Scholar] [CrossRef] [PubMed]
- Agerer, R. Fungal relationships and structural identity of their ectomycorhizae. Mycol. Progr. 2006, 5, 67–107. [Google Scholar] [CrossRef]
- Phillips, R. Mushrooms; Macmillan: New York, NY, USA, 2006; p. 384. [Google Scholar]
- Ter Braak, C.J.F.; Šmilauer, P. Canoco Reference Manual and User’s Guide: Software for Ordination (Version 5.0); Microcomputer Power: Ithaca, NY, USA, 2012; p. 496. [Google Scholar]
- Colwell, R.K. EstimateS: Statistical estimation of species richness and shared species from samples. Version 9. Available online: http://purl.oclc.org/estimates (accessed on 15 May 2013).
- Colwell, R.K.; Chao, A.; Gotelli, N.J.; Lin, S.-Y.; Mao, C.X.; Chazdon, R.L.; Longino, J.T. Models and estimators linking individual-based and sample-based rarefaction, extrapolation, and comparison of assemblages. J. Plant Ecol. 2012, 5, 3–21. [Google Scholar] [CrossRef]
- Nie, M.; Lu, M.; Bell, J.; Raut, S.; Pendall, E. Altered root traits due to elevated CO2: A meta-analysis. Global Ecol. Biogeogr. 2013, 22, 1095–1105. [Google Scholar] [CrossRef]
- Parrent, J.L.; Morris, W.F.; Vilgalys, R. CO2-enrichment and nutrient availability alter ectomycorrhizal fungal communities. Ecology 2006, 87, 2278–2287. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, S.G.; Taylor, B.N.; Cooper, E.R.; Beidler, K.V.; Strand, A.E.; McCormack, M.L.; Zhang, S. Long-term dynamics of mycorrhizal root tips in a loblolly pine forest grown with free-air CO2 enrichment and soil N fertilization for 6 years. Glob. Change Biol. 2014, 20, 1313–1326. [Google Scholar] [CrossRef]
- Norby, R.J.; Warren, J.M.; Iversen, C.M.; Medlyn, B.E.; McMurtrie, R.E. CO2 enhancement of forest productivity constrained by limited nitrogen availability. Proc. Natl. Acad. Sci. USA 2010, 107, 19368–19373. [Google Scholar] [CrossRef] [PubMed]
- Lukac, M.; Calfapietra, C.; Godbold, D.L. Production, turnover and mycorrhizal colonisation of root systems of three Populus species grown under elevated CO2 (POPFACE). Glob. Change Biol. 2003, 9, 838–848. [Google Scholar] [CrossRef]
- Parrent, J.L.; Vilgalys, R. Biomass and compositional responses of ectomycorrhizal fungal hyphae to elevated CO2 and nitrogen fertilization. New Phytol. 2007, 176, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Saikkonen, K.; Ahonen-Jonnarth, U.; Markkola, A.M.; Helander, M.; Tuomi, J.; Roitto, M.; Ranta, H. Defoliation and mycorrhizal symbiosis: A functional balance between carbon sources and below-ground sinks. Ecol. Lett. 1999, 2, 19–26. [Google Scholar] [CrossRef]
- Markkola, A.; Kuikka, K.; Rautio, P.; Härmä, E.; Roitto, M.; Tuomi, J. Defoliation increases carbon limitation in ectomycorrhizal symbiosis of Betula pubescens. Oecologia 2004, 140, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Saravesi, K.; Markkola, A.M.; Rautio, P.; Roitto, M.; Tuomi, J. Defoliation causes parallel temporal responses in a host tree and its fungal symbionts. Oecologia 2008, 156, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Fransson, P.M.A. Elevated CO2 impacts ectomycorrhiza-mediated forest soil carbon flow: Fungal biomass production, respiration and exudation. Fungal Ecol. 2012, 5, 85–98. [Google Scholar] [CrossRef]
- Pritchard, S.G.; Strand, A.E.; McCormack, M.L.; Davis, M.A.; Oren, R. Mycorrhizal and rhizomorph dynamics in a loblolly pine forest during 5 years of freeair-CO2-enrichment. Glob. Change Biol. 2008, 14, 1252–1264. [Google Scholar] [CrossRef]
- Agerer, R. Exploration and exploitation strategies of ectomycorrhizal fungi. Nova Act. Lc. 2013, 114, 201–219. [Google Scholar]
- Agerer, R.; Hartmann, A.; Pritsch, K.; Raidl, S.; Schloter, M.; Verma, R.; Weigt, R. Plants and Their Ectomycorrhizosphere: Cost and Benefit of Symbiotic Soil Organisms. In Growth and Defence in Plants, Ecological Studies 220; Matyssek, R., Schnyder, H., Oßwald, W., Ernst, D., Munch, J.C., Pretzsch, H., Eds.; Springer-Verlag: Berlin, Germany, 2012; pp. 213–242. [Google Scholar]
- Vasutova, M. Global Change Research Centre AS CR, Ceske Budejovice, Czech Republic. Unpublished Work. 2014. [Google Scholar]
- Rudawska, M.; Leski, T.; Trocha, L.K.; Gornowicz, R. Ectomycorrhizal status of Norway spruce seedlings from bare-root forest nurseries. For. Ecol. Manag. 2006, 236, 375–384. [Google Scholar] [CrossRef]
- Andrew, C.; Lilleskov, E.A. Elevated CO2 and O3 effects on ectomycorrhizal fungal root tip communities in consideration of a post-agricultural soil nutrient gradient legacy. Mycorrhiza 2014, 24, 581–593. [Google Scholar] [CrossRef] [PubMed]
- Andrew, C.; Lilleskov, E.A. Productivity and community structure of ectomycorrhizal fungal sporocarps under increased atmospheric CO2 and O3. Ecol. Lett. 2009, 12, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Last, F.T.; Dighton, J.; Mason, P.A. Successions of sheathing mycorrhizal fungi. Trends Ecol. Evol. 1987, 2, 157–161. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Godbold, D.L.; Vašutová, M.; Wilkinson, A.; Edwards-Jonášová, M.; Bambrick, M.; Smith, A.R.; Pavelka, M.; Cudlin, P. Elevated Atmospheric CO2 Affects Ectomycorrhizal Species Abundance and Increases Sporocarp Production under Field Conditions. Forests 2015, 6, 1256-1273. https://doi.org/10.3390/f6041256
Godbold DL, Vašutová M, Wilkinson A, Edwards-Jonášová M, Bambrick M, Smith AR, Pavelka M, Cudlin P. Elevated Atmospheric CO2 Affects Ectomycorrhizal Species Abundance and Increases Sporocarp Production under Field Conditions. Forests. 2015; 6(4):1256-1273. https://doi.org/10.3390/f6041256
Chicago/Turabian StyleGodbold, Douglas L., Martina Vašutová, Anna Wilkinson, Magda Edwards-Jonášová, Michael Bambrick, Andrew R. Smith, Marian Pavelka, and Pavel Cudlin. 2015. "Elevated Atmospheric CO2 Affects Ectomycorrhizal Species Abundance and Increases Sporocarp Production under Field Conditions" Forests 6, no. 4: 1256-1273. https://doi.org/10.3390/f6041256
APA StyleGodbold, D. L., Vašutová, M., Wilkinson, A., Edwards-Jonášová, M., Bambrick, M., Smith, A. R., Pavelka, M., & Cudlin, P. (2015). Elevated Atmospheric CO2 Affects Ectomycorrhizal Species Abundance and Increases Sporocarp Production under Field Conditions. Forests, 6(4), 1256-1273. https://doi.org/10.3390/f6041256