
Localized Effects of Coarse Woody Material on Soil Oribatid Communities Diminish over 700 Years of Stand Development in Black-Spruce-Feathermoss Forests

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site

{kind=link}
{kind=link}
{kind=link}
| Stand | Time since fire | Chronosequence stage | Basal area (m2/ha) | CWM volume (m3/ha) | Bryophyte thickness (cm) |
|---|---|---|---|---|---|
| N4 | 60 | 1 | 39.59 | 7.44 | 26.65 |
| N23 | 94 | 1 | 46.00 | 19.73 | 15.15 |
| S1 | 95 | 1 | 45.03 | 5.66 | 22.40 |
| Ch75 | 133 | 2 | 25.93 | 8.67 | 44.70 |
| N18 | 134 | 2 | 35.55 | 8.53 | 46.40 |
| N8 | 183 | 2 | 34.51 | 24.92 | 43.15 |
| POP | 183 | 2 | 13.86 | 53.61 | 48.05 |
| L22 | 283 | 2 | 21.25 | 57.18 | 49.45 |
| N50 | 373 | 2 | 19.65 | 47.80 | 48.30 |
| N6 | 718 | 3 | 21.70 | 33.84 | 58.75 |
2.2. Sampling Design
2.3. Statistical Analyses
3. Results
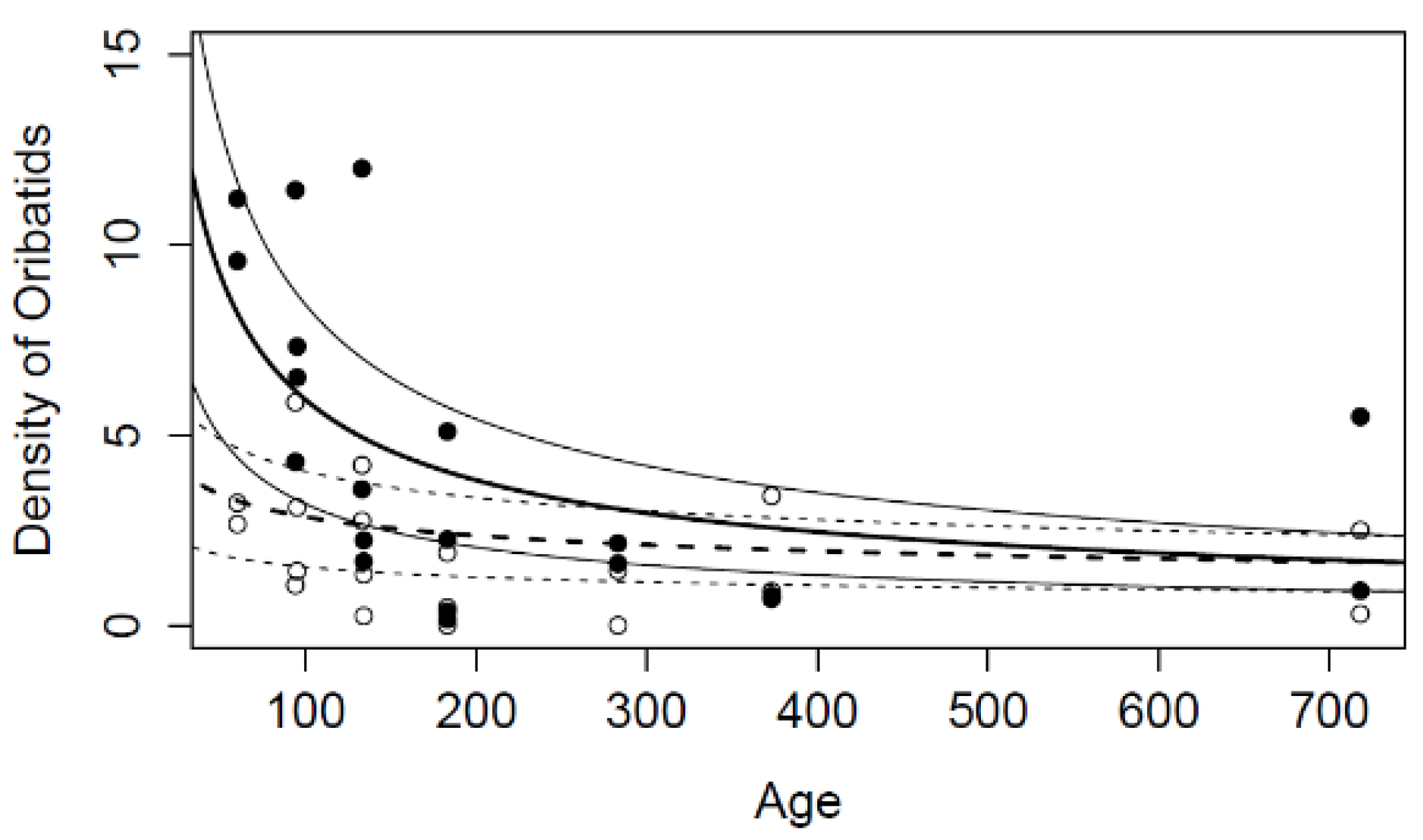
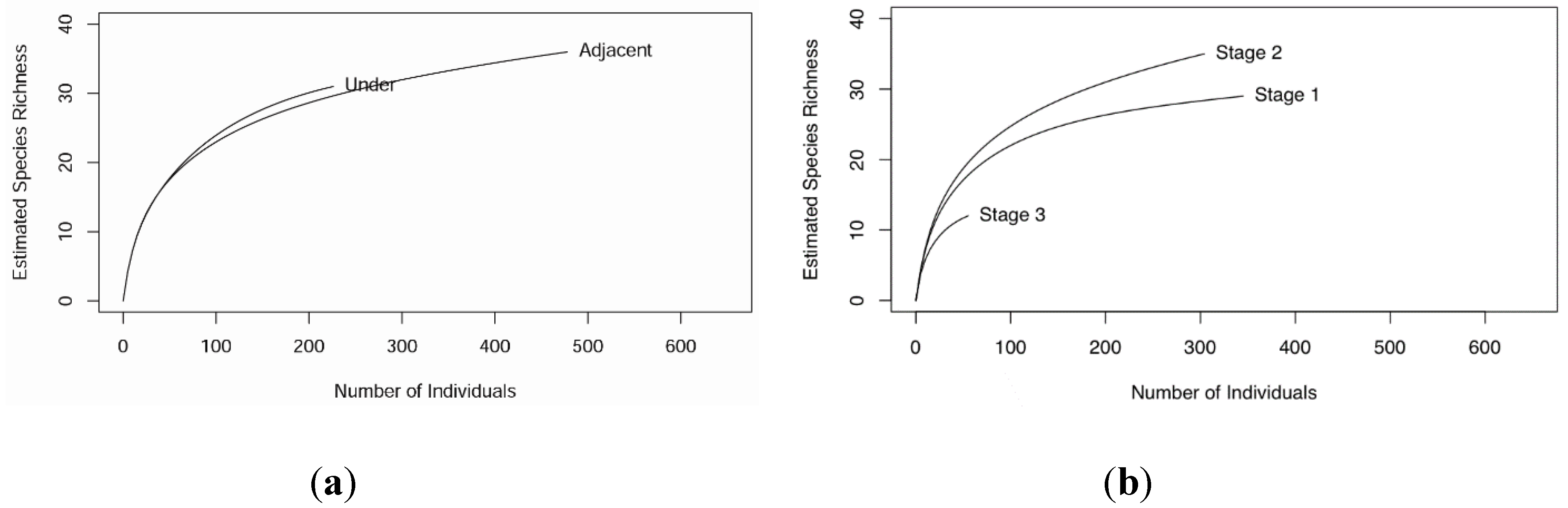
3.1. Soil Microarthropod Community
3.2. Oribatid Distribution

| Parameter | Value | Std. Error | DF ** | t-value | p-value |
|---|---|---|---|---|---|
| Total Oribatids | |||||
| Intercept (Position Adjacent *) | 4.71 | 1.22 | 27 | 3.85 | 0.0007 |
| Age | −0.64 | 0.23 | 8 | −2.71 | 0.0268 |
| Position Under | −2.41 | 1.15 | 27 | −2.10 | 0.0451 |
| Age:Position Under | 0.36 | 0.22 | 27 | 1.65 | 0.1103 |
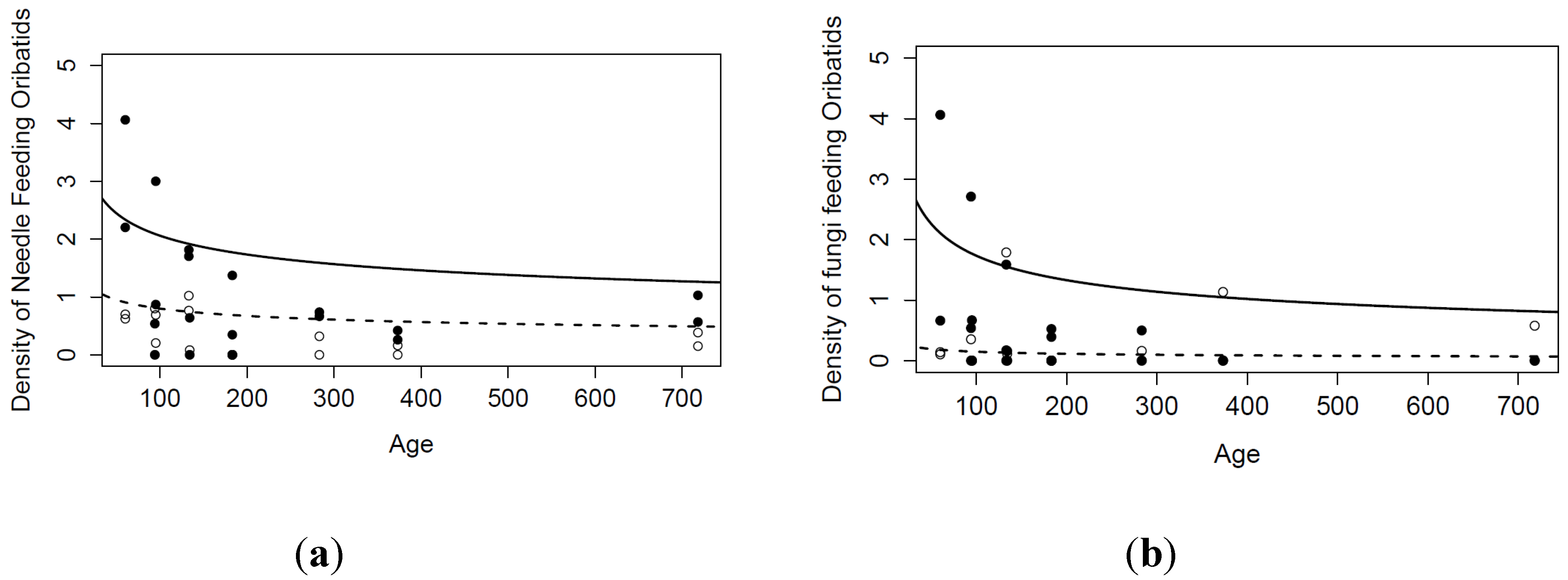
| Litter feeders | |||||
| Intercept (Position Adjacent) | 1.86 | 0.77 | 27 | 2.41 | 0.0232 |
| Age | −0.25 | 0.15 | 8 | −1.66 | 0.1344 |
| Position Under | −0.95 | 0.61 | 27 | −1.55 | 0.1320 |
| Age:Position Under | 0.11 | 0.12 | 27 | 0.96 | 0.3439 |
| Fungi feeders | |||||
| Intercept (Position Adjacent) | 2.23 | 0.57 | 27 | 3.88 | 0.0006 |
| Age | −0.37 | 0.11 | 8 | −3.36 | 0.0100 |
| Position Under | −2.36 | 0.76 | 27 | −3.12 | 0.0043 |
| Age:Position Under | 0.42 | 0.14 | 27 | 2.92 | 0.0070 |


3.3. Microclimatic and Environmental Factors Related to Oribatid Density
| Variable | Oribatid density (individuals/g of dry substrate) | ||
|---|---|---|---|
| Rho* | t-value | p-value | |
| CWM Diameter 1 | −0.463 | −2.153 | 0.04599 |
| CWM Density 1 | −0.014 | −0.059 | 0.95360 |
| Moisture 2 | 0.343 | 2.2202 | 0.03261 |
| Variable | Oribatid density (individuals/g of dry substrate) | ||
|---|---|---|---|
| Rho* | t-value | p-value | |
| Basal Area (m2/ha) | 0.573 | 1.975 | 0.0837 |
| CWM Volume (m3/ha) | −0.704 | −2.807 | 0.0230 |
| Organic layer thickness (cm) | −0.452 | −1.434 | 0.1895 |
4. Discussion
5. Conclusions
Acknowledgements
Author Contribution
Conflicts of Interest
Appendix
| Family | Species | N | Relative Abundance % | Trophic guild |
|---|---|---|---|---|
| Hypochthoniidae | Hypochthonius rufulus C.L. Koch, 1835 | 4 | 0.57 | |
| Eniochthoniidae | Eniochthonius minutissimus (Berlese, 1903) | 55 | 7.81 | Fungi |
| Phthiracaridae | Atropacarus striculus (C.L. Koch, 1835) | 111 | 15.77 | Litter |
| Phthiracarus sp. | 17 | 2.41 | ||
| Oribotritiidae | Protoribotritia canadaris Jacot, 1938 | 9 | 1.28 | |
| Euphthiracaridae | Rhysotritia ardua (C.L. Koch, 1841) | 40 | 5.68 | Litter |
| Nothridae | Nothrus sp. | 80 | 11.36 | Unspecif. |
| Camisiidae | Camisia lapponica (Trägårdh, 1910) | 11 | 1.56 | |
| Camisia sp. 2 | 6 | 0.85 | ||
| Trhypochthoniidae | Trhypochthonius tectorum (Berlese, 1896) | 27 | 3.84 | |
| Nanhermanniidae | Nanhermannia elegantula Berlese, 1913 | 5 | 0.71 | |
| Nanhermannia sp. 2 | 30 | 4.26 | Fungi | |
| Hermanniidae | Hermannia sp. | 76 | 10.80 | Unspecif. |
| Plateremaeidae | Allodamaeus sp. | 11 | 1.56 | |
| Damaeidae | Belba sp. | 8 | 1.14 | |
| Cepheidae | Cepheus corae Jacot, 1928 | 2 | 0.28 | |
| Eremaeidae | Eremaeus sp. | 1 | 0.14 | |
| Tenuialidae | Tenuilaoides sp. 1 | 1 | 0.14 | |
| Tenuilaoides sp. 2 | 19 | 2.70 | ||
| Tenuilaoides sp. 3 | 1 | 0.14 | ||
| Peloppiidae | Ceratoppia quaridentata arctica Hammer, 1955 | 10 | 1.42 | |
| Carabodidae | Carabodes sp. | 6 | 0.85 | |
| Carabodidae sp. 2 | 4 | 0.57 | ||
| Oppiidae | Oppia sp. | 27 | 3.84 | |
| Oppiidae sp. 2 | 1 | 0.14 | ||
| Oppiidae sp. 3 | 5 | 0.71 | ||
| Thyrisomidae | Banksinoma spinifera (Hammer, 1952) | 3 | 0.43 | |
| Cymbaeremaeidae | Scapheremaeus palustris (Sellnick, 1924) | 2 | 0.28 | |
| Achipteriidae | Achipteria sp. | 1 | 0.14 | |
| Parachipteria bella (Sellnick, 1928) | 3 | 0.43 | ||
| Achipteriidae sp. 3 | 1 | 0.14 | ||
| Achipteriidae sp. 4 | 8 | 1.14 | ||
| Phenopelopidae | Peloptulus sp. | 6 | 0.85 | |
| Scheloribatidae | Scheloribates pallidulus (C.L. Koch, 1841) | 59 | 8.38 | Plant tiss. |
| Oribatulidae | Eporibatula modesta (Banks, 1904) | 1 | 0.14 | |
| Phauloppia boletorum (Ewing, 1913) | 17 | 2.41 | ||
| Haplozetidae | Xylobates sp. | 3 | 0.43 | |
| Galumnidae | Pergalumna sp. 1 | 30 | 4.26 | Unspecif. |
| Pergalumna sp. 2 | 3 | 0.43 |
References
- Wardle, D.A.; Walker, L.R.; Bardgett, R.D. Ecosystem properties and forest decline in contrasting long-term chronosequences. Science 2004, 305, 509–513. [Google Scholar] [CrossRef] [PubMed]
- Lagerström, A.; Esberg, C.; Wardle, D.A.; Giesler, R. Soil phosphorus and microbial response to a long-term wildfire chronosequence in northern Sweden. Biogeochemistry 2009, 95, 199–213. [Google Scholar] [CrossRef]
- Wardle, D.A.; Bardgett, R.D.; Walker, L.R.; Bonner, K.I. Among- and within-species variation in plant litter decomposition in contrasting long-term chronosequences. Funct. Ecol. 2009, 23, 442–453. [Google Scholar] [CrossRef]
- Peltzer, D.A.; Wardle, D.A.; Allison, V.J.; Baisden, W.T.; Bardgett, R.D.; Chadwick, O.A.; Condron, L.M.; Parfitt, R.L.; Porder, S.; Richardson, S.J.; Turner, B.L.; Vitousek, P.M.; Walker, J.; Walker, L.R. Understanding ecosystem retrogression. Ecol. Monogr. 2010, 80, 509–529. [Google Scholar] [CrossRef]
- Doblas-Miranda, E.; Wardle, D.A.; Peltzer, D.A.; Yeates, G.W. Changes in the community structure and diversity of soil invertebrates across the Franz Josef Glacier chronosequence. Soil Biol. Biochem. 2008, 40, 1069–1081. [Google Scholar] [CrossRef]
- Wardle, D.A.; Hörnberg, G.; Zackrisson, O.; Kalela-Brundin, M.; Coomes, D.A. Long-term effects of wildfire on ecosystem properties across an island area gradient. Science 2003, 300, 972–975. [Google Scholar] [CrossRef] [PubMed]
- Williamson, W.M.; Wardle, D.A.; Yeates, G.W. Changes in soil microbial and nematode communities during ecosystem decline across a long-term chronosequence. Soil Biol. Biochem. 2005, 37, 1289–1301. [Google Scholar] [CrossRef]
- Simard, M.; Lecomte, N.; Bergeron, Y.; Bernier, P.Y.; Paré, D. Forest productivity decline caused by successional paludification of boreal soils. Ecol. Appl. 2007, 17, 1619–1637. [Google Scholar] [CrossRef] [PubMed]
- Lavoie, M.; Paré, D.; Fenton, N.; Groot, A.; Taylor, K. Paludification and management of forested peatlands in Canada: A literature review. Environ. Rev. 2005, 13, 21–50. [Google Scholar] [CrossRef]
- Van Cleve, K.; Dyrness, C.T.; Viereck, L.A.; Fox, J.; Chapin, F.S.; Oechel, W. Taiga ecosystems in interior Alaska. BioScience 1983, 33, 39–44. [Google Scholar] [CrossRef]
- Brock, T.B.C.; Bregman, R. Periodicity in growth, productivity, nutrient content and decomposition of Sphagnum recurvum var. mucronatum in a fen woodland. Oecologia 1989, 80, 44–52. [Google Scholar]
- Laiho, R.; Prescott, C.E. Decay and nutrient dynamics of coarse woody debris in northern coniferous forest: A synthesis. Can. J. For. Res. 2004, 34, 763–777. [Google Scholar] [CrossRef]
- Brais, S.; Paré, D.; Lierman, C. Tree bole mineralization rates of four species of the Canadian eastern boreal forest: Implications for nutrient dynamics following stand-replacing disturbances. Can. J. For. Res. 2006, 36, 2331–2340. [Google Scholar] [CrossRef]
- Moore, J.C.; deRuiter, P.C.; Hunt, H.W.; Coleman, D.C.; Freckman, D.W. Microcosms and soil ecology: Critical linkages between field studies and modelling food webs. Ecology 1996, 77, 694–705. [Google Scholar] [CrossRef]
- Means, J.E.; MacMillan, P.C.; Cromack, K., Jr. Biomass and nutrient content of Douglas-fir logs and otehr detrital pools in an old growth forest. Can. J. For. Res. 1992, 22, 1536–1546. [Google Scholar] [CrossRef]
- Brais, S.; Sadi, F.; Bergeron, Y.; Grenier, Y. Coarse woody debris dynamics in a post-fire jack-pine chronosequence and its relation with site productivity. For. Ecol. Manag. 2005, 220, 216–226. [Google Scholar] [CrossRef]
- Déchêne, A.D.; Buddle, C.M. Decomposing logs increase oribatid mite assemblage diversity in mixedwood boreal forest. Biodivers. Conserv. 2010, 19, 237–256. [Google Scholar] [CrossRef]
- Marra, J.L.; Edmonds, R.L. Effects of coarse woody debris and soil depth on the density and diversity of soil invertebrates on clearcut and forested sites on the Olympic Peninsula, Washington. Environ. Entomol. 1998, 27, 1111–1124. [Google Scholar] [CrossRef]
- Evans, A.M.; Clinton, P.W.; Allen, R.B.; Frampton, C.M. The influence of logs on the spatial distribution of litter-dwelling invertebrates and forest floor processes in New Zealand forests. For. Ecol. Manag. 2003, 184, 251–262. [Google Scholar] [CrossRef]
- Harper, K.A.; Bergeron, Y.; Drapeau, P.; Gauthier, S.; De Grandpré, L. Structural development following fire in black spruce boreal forest. For. Ecol. Manag. 2005, 206, 293–306. [Google Scholar] [CrossRef]
- Lecomte, N.; Simard, M.; Fenton, N.; Bergeron, Y. Fire severity and long-term ecosystem biomass dynamics in coniferous boreal forests of eastern Canada. Ecosystems 2006, 9, 1215–1230. [Google Scholar] [CrossRef]
- Crawford, R.M.M.; Jeffree, C.E.; Rees, W.G. Paludification and forest retreat in northern oceanic environments. Ann. Bot. 2003, 91, 213–226. [Google Scholar] [CrossRef] [PubMed]
- Fenton, N.J.; Bergeron, Y. Facilitative succession in a boreal bryophyte community driven by changes in available moisture and light. J. Veg. Sci. 2006, 17, 65–76. [Google Scholar] [CrossRef]
- Hagemann, U.; Moroni, M.T.; Makeschin, F. Deadwood abundance in Labrador high-boreal black spruce forests. Can. J. For. Res. 2009, 39, 131–142. [Google Scholar] [CrossRef]
- Dynesius, M.; Gibb, H.; Hjältén, J. Surface covering of downed logs: Drivers of a neglected process in dead wood ecology. PLoS ONE 2010, 5, e13237. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, S.; De Grandpré, L.; Bergeron, Y. Differences in forest composition in two boreal forest ecoregions of Quebec. J. Veg. Sci. 2000, 11, 781–790. [Google Scholar] [CrossRef]
- Environment Canada. Canadian Climate Normals or Averages 1971–2000. National Climate Data and Information Archive. 2000. Available online: http://www.climate.weatheroffice.ec.gc.ca/climate_normals/index_e.html (accessed on 21 June 2010).
- Clayton, M.; Humble, L. Microarthropod Voucher Specimens; Canadian Forest Service: Victoria, BC, Canada, 1999. [Google Scholar]
- Krivolutsky, D.A. The Oribatid Mites; Naukaî: Moscow, Russia, 1995. [Google Scholar]
- Balogh, J. The Oribatid Genera of the World; Académiai Kiadó: Budapest, Hungary, 1972. [Google Scholar]
- Niedbała, W. Ptyctimous Mites (Acari, Oribatida) of the Nearctic Region; Monographs of the Upper Silesian Museum 4: Bytom, Poland, 2002. [Google Scholar]
- Norton, R.A.; Behan-Pelletier, V.M. Eniochthonius Mahunkai sp. n. (Acari: Oribatida: Enionchthoniidae), from north american peatlands, with a redescription of Eniochthonius and key to north american species. Acta Zool. Acad. Sci. Hung. 2007, 53, 295–333. [Google Scholar]
- Karg, W. The armoured mite Scheloribates pallidulus C.L. Koch as a pest of cucumber crops under glass and plastic covers. Arch. Phytopathol. Plant Prot. 1984, 20, 181–183. [Google Scholar] [CrossRef]
- Siepel, H. Niche relationships between two panphytophagous soil mites, Nothrus silvestris Nicolet (Acari, Oribatida, Nothridae) and Platynothrus peltifer (Koch) (Acari, Oribatida, Camisiidae). Biol. Fert. Soils 1990, 9, 139–144. [Google Scholar] [CrossRef]
- Schneider, K.; Migge, S.; Norton, R.A.; Scheu, S.; Langel, R.; Reineking, A.; Maraun, M. Trophic niche differentiation in soil microartropods (Oribatida, Acari): Evidence from stable isotope ratios (15N/14N). Soil Biol. Biochem. 2004, 36, 1769–1774. [Google Scholar] [CrossRef]
- Norton, R.A.; Behan-Pelletier, V.M. Calcium carbonate and calcium oxalate as cuticular hardening agents in oribatid mites (Acari: Oribatida). Can. J. Zool. 1991, 69, 1504–1511. [Google Scholar] [CrossRef]
- Oliveira, A.R.; de Moraes, G.J.; Ferraz, L.C. Consumption rate of phytonematodes by Pergalumna sp. (Acari: Oribatida: Galumnidae) under laboratory conditions determined by a new method. Exp. Appl. Acarol. 2007, 41, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Robert, E.; Brais, S.; Harvey, B.D.; Greene, D. Seedling establishment and survival on decaying logs in boreal mixedwood stands following a mast year. Can. J. For. Res. 2012, 42, 1446–1455. [Google Scholar] [CrossRef]
- Zuur, A.; Ieno, E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer: New York, NY, USA, 2009. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2010. [Google Scholar]
- Colwell, R.K.; Chao, A.; Gotelli, N.J.; Lin, S.-Y.; Mao, C.X.; Chazdon, R.L.; Longino, J.T. Models and estimators linking individual-based and sample-based rarefaction, extrapolation and comparison of assemblages. J. Plant. Ecol. 2012, 5, 3–21. [Google Scholar] [CrossRef]
- Arroyo, J.; Kenny, J.; Bolger, T. Variation between mite communities in Irish forest types-Importance of bark and moss cover in canopy. Pedobiologia 2013, 56, 241–250. [Google Scholar] [CrossRef]
- Farska, J.; Prejzkova, K.; Rusek, J. Management intensity affects traits of soil microarthropod community in montane spruce forest. Appl. Soil Eco. 2014, 75, 71–79. [Google Scholar] [CrossRef]
- Zaitsev, A.S.; Chauvat, M.; Wolters, V. Spruce forest conversion to a mixed beech-coniferous stand modifies oribatid community structure. Appl. Soil Eco. 2014, 76, 60–67. [Google Scholar] [CrossRef]
- Paquin, P. Carabid beetle (Coleoptera: Carabidae) diversity in the black spruce succession of eastern Canada. Biol. Conserv. 2008, 141, 261–275. [Google Scholar] [CrossRef]
- Paradis, S. Évaluation de l'effet des Coupes Partielles sur les Arthropodes Terricoles dans des Peuplements D'épinette noire Sujets à la Paludification. Ph. D. Thesis, Université du Québec à Montréal, Montreal, Canada, 2010. [Google Scholar]
- Migge, S.; Maraun, M.; Scheu, S.; Schaefer, M. The oribatid mite community (Acarina) of pure and mixed stands of beech (Fagus sylvatica) and spruce (Picea abies) of different age. Appl. Soil Ecol. 1998, 9, 115–121. [Google Scholar] [CrossRef]
- Zaitsev, A.S.; Chauvat, M.; Pflug, A.; Wolters, V. Oribatid mite diversity and community dynamics in a spruce chronosequence. Soil Biol. Biochem. 2002, 34, 1919–1927. [Google Scholar] [CrossRef]
- Salmon, S.; Artuso, N.; Frizzera, L.; Zampedri, R. Relationships between soil fauna communities and humus forms: Response to forest dynamics and solar radiation. Soil Biol. Biochem. 2008, 40, 1707–1715. [Google Scholar] [CrossRef]
- Wang, C.; Bond-Lamberty, B.; Gower, S.T. Environmental controls on carbon dioxide flux from black spruce coarse woody debris. Oecologia 2002, 132, 374–381. [Google Scholar] [CrossRef]
- Seastedt, T.R.; Reddy, M.V.; Cline, S.P. Microarthropods in decaying wood from temperate coniferous and deciduous forests. Pedobiologia 1989, 33, 69–77. [Google Scholar]
- Zak, D.R.; Groffman, P.M.; Pregitzer, K.S.; Christensen, S.; Tiedje, J.M. The vernal dam: Plant-microbe competition for nitrogen in Northern hardwood forests. Ecology 1990, 71, 651–656. [Google Scholar] [CrossRef]
- Fenton, N.; Lecomte, N.; Légaré, S.; Bergeron, Y. Paludification in black spruce (Picea mariana) forests of eastern Canada: Potential factors and management implications. For. Ecol. Manag. 2005, 213, 151–159. [Google Scholar] [CrossRef]
- Skubała, P.; Sokołowska, M. Oribatid fauna (Acari, Oribatida) in fallen spruce trees in the Babia Góra National Park. Biol. Lett. 2006, 43, 243–248. [Google Scholar]
- Siira-Pietikainen, A.; Penttinen, R.; Huhta, V. Oribatid mites (Acari:Oribatida) in boreal forest floor and decaying wood. Pedobiologia 2008, 52, 111–118. [Google Scholar] [CrossRef]
- Nielsen, U.N.; Osler, G.H.R.; van der Wal, R.; Campbell, C.D.; Burslem, D.F.R.P. Soil pore volume and the abundance of soil mites in two contrasting habitats. Soil Biol. Biochem. 2008, 40, 1538–1541. [Google Scholar] [CrossRef]
- Lindberg, N.; Bengtsson, J.B.; Persson, T. Effects of experimental irrigation and drought on the composition and diversity of soil fauna in a coniferous stand. J. Appl. Ecol. 2002, 39, 924–936. [Google Scholar] [CrossRef]
- Tsiafouli, M.A.; Kallimanis, A.S.; Katana, E.; Stamou, G.P.; Sgardelis, S.P. Responses of soil microarthropods to experimental short-term manipulations of soil moisture. Appl. Soil Ecol. 2005, 29, 17–26. [Google Scholar] [CrossRef]
- Siepel, H. The importance of unpredictable and short-term environmental extremes for biodiversity in oribatid mites. Biodiv. Let. 1996, 3, 26–34. [Google Scholar] [CrossRef]
- Read, D.J.; Leake, J.R.; Perez-Moreno, J. Mycorrhizal fungi as drivers of ecosystem processes in heathland and boreal forest biomes. Can. J. Bot. 2004, 82, 1243–1263. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Doblas-Miranda, E.; Work, T.T. Localized Effects of Coarse Woody Material on Soil Oribatid Communities Diminish over 700 Years of Stand Development in Black-Spruce-Feathermoss Forests. Forests 2015, 6, 914-928. https://doi.org/10.3390/f6040914
Doblas-Miranda E, Work TT. Localized Effects of Coarse Woody Material on Soil Oribatid Communities Diminish over 700 Years of Stand Development in Black-Spruce-Feathermoss Forests. Forests. 2015; 6(4):914-928. https://doi.org/10.3390/f6040914
Chicago/Turabian StyleDoblas-Miranda, Enrique, and Timothy T. Work. 2015. "Localized Effects of Coarse Woody Material on Soil Oribatid Communities Diminish over 700 Years of Stand Development in Black-Spruce-Feathermoss Forests" Forests 6, no. 4: 914-928. https://doi.org/10.3390/f6040914
APA StyleDoblas-Miranda, E., & Work, T. T. (2015). Localized Effects of Coarse Woody Material on Soil Oribatid Communities Diminish over 700 Years of Stand Development in Black-Spruce-Feathermoss Forests. Forests, 6(4), 914-928. https://doi.org/10.3390/f6040914