
Foundation Species Loss and Biodiversity of the Herbaceous Layer in New England Forests


Abstract
:1. Introduction
2. Materials and Methods
2.1. Eastern Hemlock
2.2. The Harvard Forest Hemlock Removal Experiment
2.3. The Herbaceous Layer
3. Results and Discussion
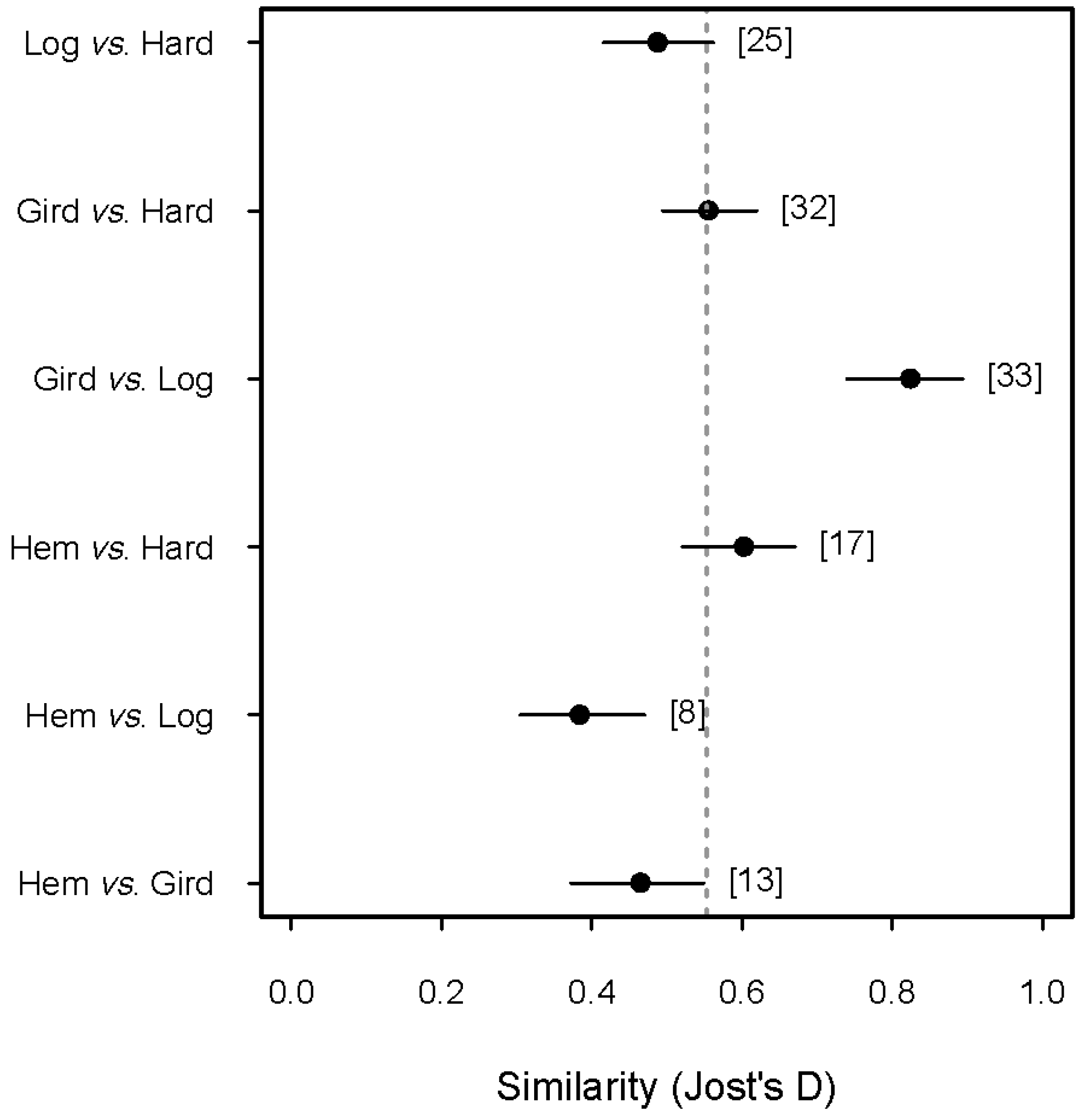
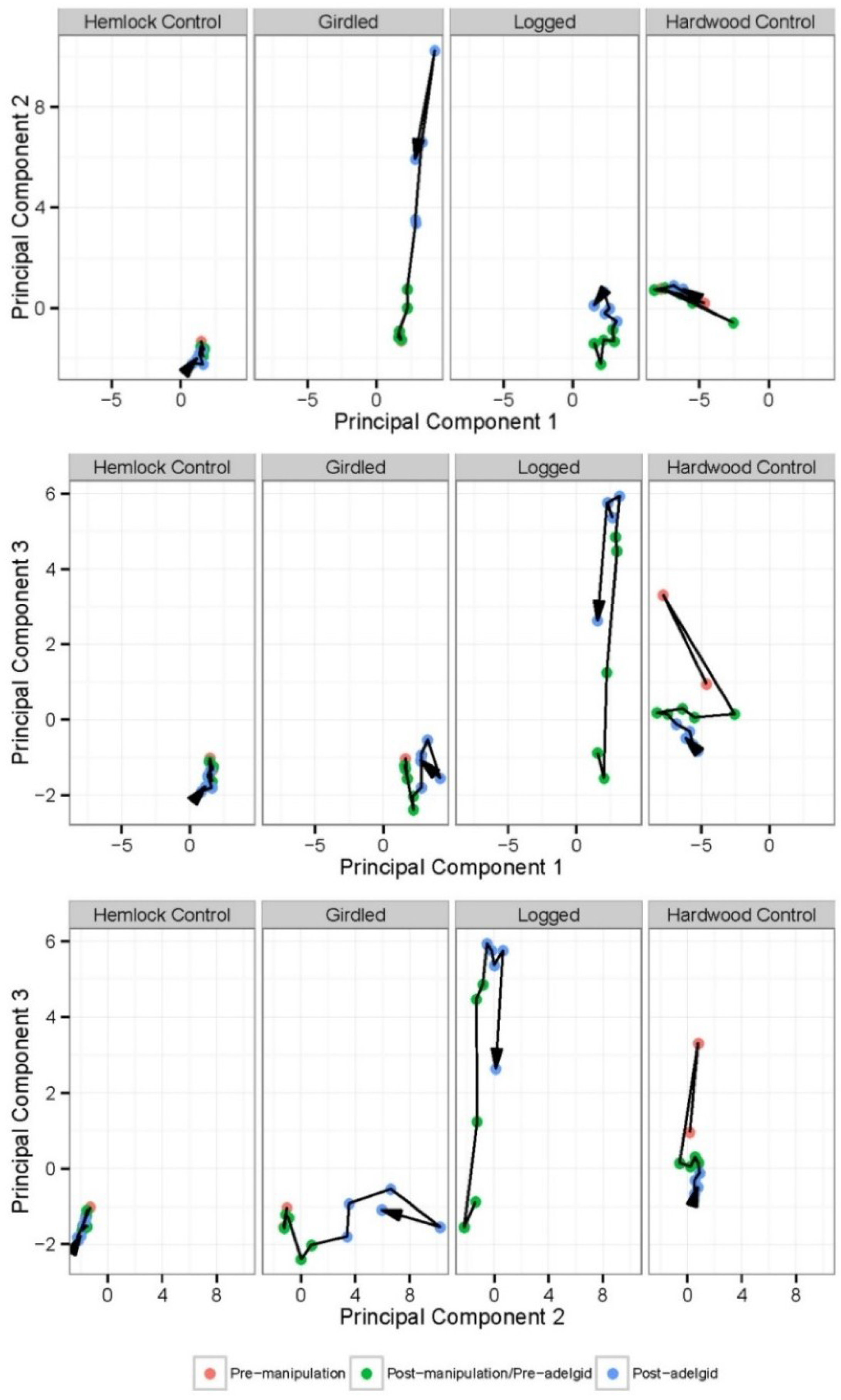
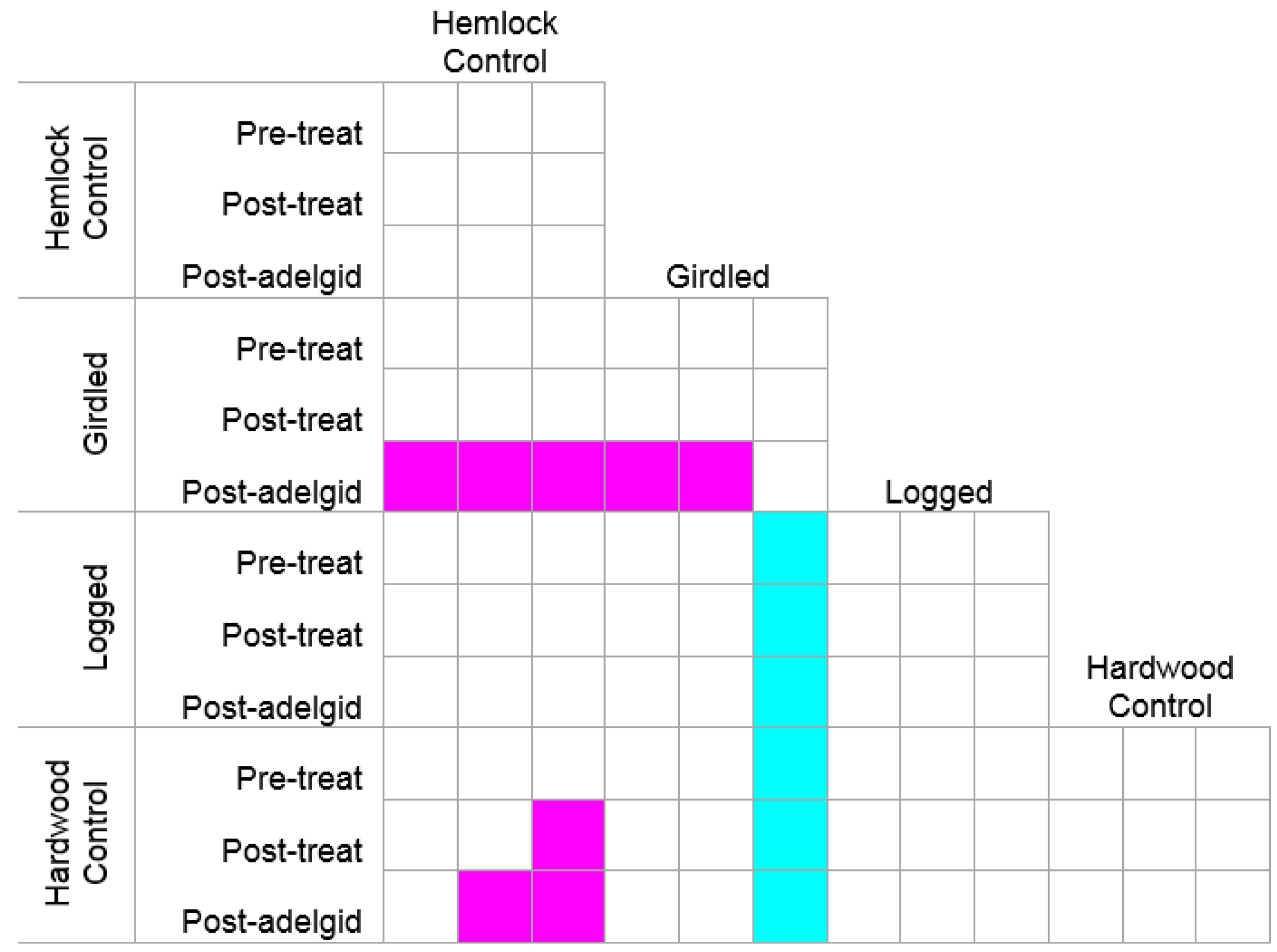
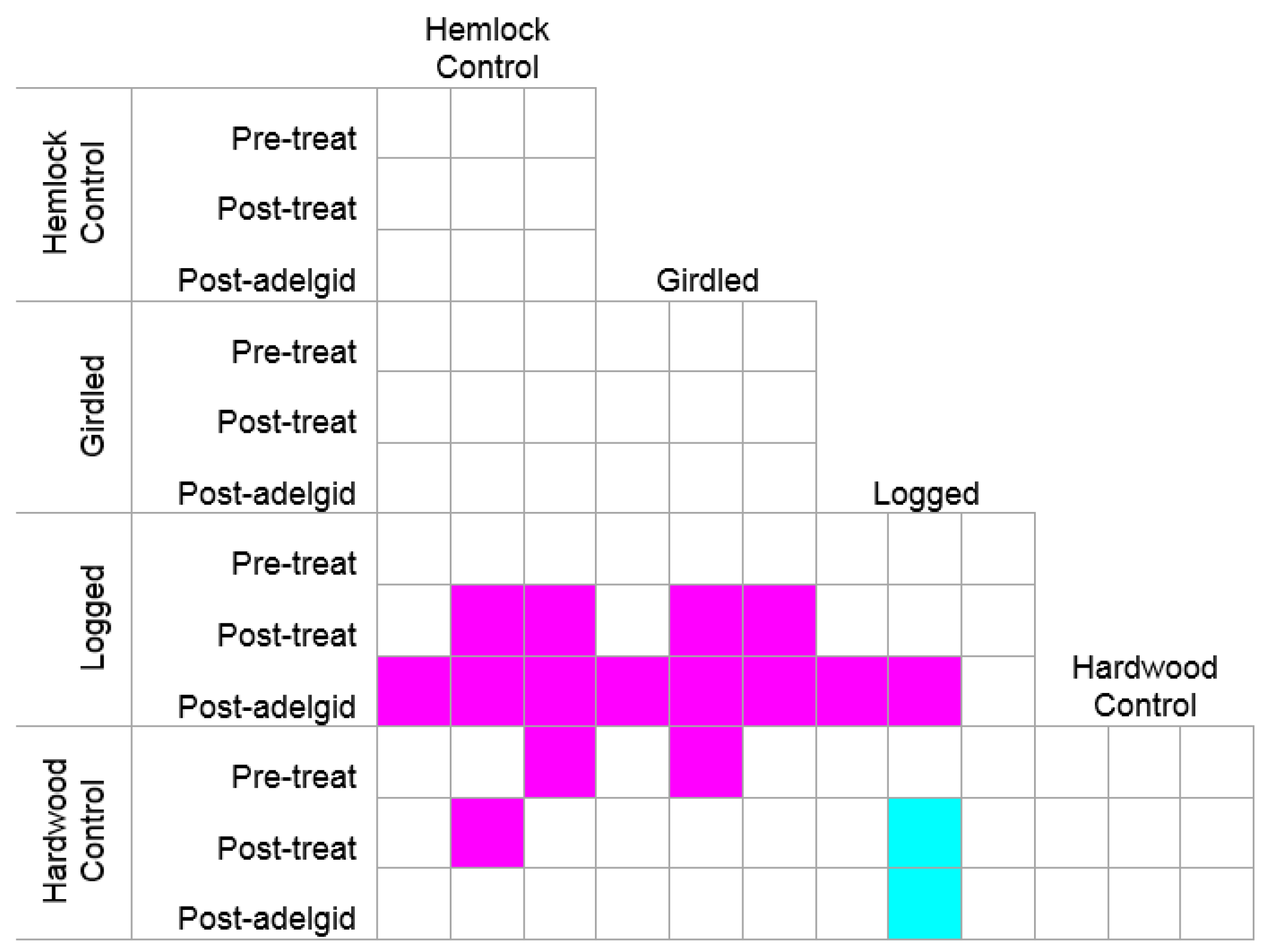
3.1. The Response of the Herbaceous Layer to Experimental Removal of Eastern Hemlock

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Incidences | Sobs | 0q | 1q | 2q | |
|---|---|---|---|---|---|
| Hemlock control | 188 | 18 | 19.9 (16.9, 23.0) | 15.9 (14.8, 17.1) | 14.9 (13.4, 16.8) |
| Girdled | 388 | 53 | 54.2 (45.0, 63.5) | 42.8 (39.5, 46.0) | 36.7 (33.3, 40.0) |
| Logged | 305 | 38 | 50.2 (17.7, 82.8) | 30.4 (27.2, 33.5) | 25.9 (23.1, 28.7) |
| Hardwood control | 616 | 51 | 52.6 (40.0, 65.2) | 41.1 (39.1, 43.1) | 37.2 (35.0, 39.4) |
| All treatments pooled | 1497 | 73 | 73.5 (73.0, 81.4) | 53.8 (50.2, 54.9) | 43.2 (39.3, 44.6) |


| Species | PC-1 | PC-2 | PC-3 |
|---|---|---|---|
| Aralia nudicaulis L. | −0.26 | ||
| Dendrolycopodium obscurum (L). A. Haines | −0.26 | ||
| Lysimachia borealis (Raf.) U. Manns & A. Anderb. | −0.25 | ||
| Berberis thunbergii DC. | 0.36 | ||
| Lonicera canadensis Bartr. Ex Marsh. | 0.36 | ||
| Dryopteris carthusiana (Vill.) H.P. Fuchs | 0.32 | ||
| Ilex verticillata (L.) Gray | 0.32 | ||
| Osmundastrum cinnamomeum (L.) C. Presl | 0.31 | ||
| Epigaea repens L. | 0.31 | ||
| Viburnum acerifolium L. | 0.31 | ||
| Rhododendron periclymenoides (Michx.) Shinners | 0.26 | ||
| Lysimachia quadrifolia L. | 0.38 | ||
| Carex sp. | 0.34 | ||
| Rubusallegheniensis Porter | 0.31 | ||
| Araliahispida Veng. | 0.30 | ||
| Rubusidaeus L. | 0.28 | ||
| Carex cf. pennsylvanica Lam. | 0.28 | ||
| Panicum sp. | 0.27 |
| Response | Factor | df | MS | F | P |
|---|---|---|---|---|---|
| PC-1 | Temporal stratum | 2 | 1.05 | 1.21 | 0.31 |
| Treatment | 3 | 196.21 | 226.23 | <0.001 | |
| Treatment within stratum | 6 | 0.76 | 0.88 | 0.52 | |
| Residual | 36 | 0.87 | |||
| PC-2 | Temporal stratum | 2 | 22.95 | 21.68 | <0.001 |
| Treatment | 3 | 33.31 | 31.47 | <0.001 | |
| Treatment within stratum | 6 | 14.87 | 14.05 | <0.001 | |
| Residual | 36 | 1.06 | |||
| PC-3 | Temporal stratum | 2 | 3.40 | 2.53 | 0.09 |
| Treatment | 3 | 44.97 | 33.44 | <0.001 | |
| Treatment within stratum | 6 | 10.66 | 7.93 | <0.001 | |
| Residual | 36 | 1.34 |


3.2. The Herbaceous Layer in Other Eastern Hemlock Stands
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Dayton, P.K. Toward an understanding of community resilience and the potential effects of enrichments to the benthos at McMurdo Sound, Antarctica. In Proceedings of the Colloquium on Conservation Problems in Antarctica, Blacksburg, VA, USA, 10–12 September 1971; Parker, B.C., Ed.; Allen Press: Lawrence, KS, USA, 1972; pp. 81–95. [Google Scholar]
- Ellison, A.M.; Bank, M.S.; Clinton, B.D.; Colburn, E.A.; Elliott, K.; Ford, C.R.; Foster, D.R.; Kloeppel, B.D.; Knoepp, J.D.; Lovett, G.M.; et al. Loss of foundation species: Consequences for the structure and dynamics of forested ecosystems. Front. Ecol. Environ. 2005, 9, 479–486. [Google Scholar] [CrossRef]
- Baiser, B.; Whitaker, N.; Ellison, A.M. Modeling foundation species in food webs. Ecosphere 2013, 4, 146. [Google Scholar] [CrossRef]
- Sackett, T.E.; Record, S.; Bewick, S.; Baiser, B.; Sanders, N.J.; Ellison, A.M. Response of macroarthropod assemblages to the loss of hemlock (Tsuga canadensis), a foundation species. Ecosphere 2011, 2, 74. [Google Scholar] [CrossRef]
- Hanski, I. Dynamics of regional distribution: The core and satellite species hypothesis. Oikos 1982, 38, 210–221. [Google Scholar] [CrossRef]
- Grime, J.P. Dominant and subordinate components of plant communities: Implications for succession, stability and diversity. In Colonization, Succession and Stability; Gray, A.J., Crawley, M.J., Eds.; Blackwell Scientific Publishers: Oxford, UK, 1984; pp. 413–428. [Google Scholar]
- Huston, M.A. Biological Diversity: The Coexistence of Species on Changing Landscapes; Cambridge University Press: Cambridge, UK, 1994. [Google Scholar]
- Jones, C.G.; Lawton, J.H.; Shachak, M. Organisms as ecosystem engineers. Oikos 1994, 69, 373–386. [Google Scholar] [CrossRef]
- Wyse, S.V.; Burns, B.R.; Wright, S.D. Distinctive vegetation communities are associated with the long-lived conifer Agathis australis (New Zealand kauri, Araucariaceae) in New Zealand rainforests. Austral Ecol. 2014, 39, 388–400. [Google Scholar] [CrossRef]
- Schöb, C.; Butterfield, B.J.; Pugnaire, F.I. Foundation species influence trait-based community assembly. New Phytol. 2012, 196, 824–834. [Google Scholar] [CrossRef] [PubMed]
- Butterfield, B.J.; Cavieres, L.A.; Callaway, R.M.; Cook, B.J.; Kikvidze, Z.; Lortie, C.J.; Michalet, R.; Pugnaire, F.I.; Schöb, C.; Xiao, S.; et al. Alpine cushion plants inhibit the loss of phylogenetic diversity in severe environments. Ecol. Lett. 2013, 16, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Cavieres, L.A.; Brooker, R.W.; Butterfield, B.J.; Cook, B.J.; Kikvidze, Z.; Lortie, C.J.; Michalet, R.; Pugnaire, F.I.; Schöb, C.; Xiao, S.; et al. Facilitative plant interactions and climate simultaneously drive alpine plant diversity. Ecol. Lett. 2014, 17, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Kikvidze, Z.; Brooker, R.W.; Buttefield, B.J.; Callaway, R.M.; Cavieres, L.A.; Cook, B.J.; Lortie, C.J.; Michalet, R.; Pugnaire, F.I.; Xiao, S.; et al. The effects of foundation species on community assembly: A global study of alpine cushion plant communities. Ecology 2015, 96, 2064–2069. [Google Scholar] [CrossRef] [PubMed]
- Hughes, B.B. Variable effects of a kelp foundation species on rocky intertidal diversity and species interactions in central California. J. Exp. Mar. Biol. Ecol. 2010, 393, 90–99. [Google Scholar] [CrossRef]
- Gilliam, F.S. The herbaceous layer—The forest between the trees. In The Herbaceous Layer in Forests of Eastern North America, 2nd ed.; Gilliam, F.S., Ed.; Oxford University Press: Oxford, UK, 2014; pp. 1–12. [Google Scholar]
- Daubenmire, R.F. The relation of certain ecological factors to the inhibition of forest floor herbs under hemlock. Butl. Univ. Bot. Stud. 1929, 1, 61–76. Available online: http://digitalcommons.butler.edu/botanical/vol1/iss1/7 (accessed on 3 November 2015). [Google Scholar]
- Rogers, R.S. Hemlock stands from Wisconsin to Nova Scotia: Transitions in understory composition along a floristic gradient. Ecology 1980, 61, 178–193. [Google Scholar] [CrossRef]
- Mladenoff, D.J. The relationship of the soil seed bank and understory vegetation in old-growth northern hardwood-hemlock treefall gaps. Can. J. Bot. 1990, 68, 2714–2721. [Google Scholar] [CrossRef]
- D’Amato, A.W.; Orwig, D.A.; Foster, D.R. Understory vegetation in old-growth and second-growth Tsuga canadensis forests in western Massachusetts. For. Ecol. Manag. 2009, 257, 1043–1052. [Google Scholar] [CrossRef]
- Abella, S.R. Imacts and management of hemlock woolly adelgid in national parks of the eastern United States. Southeast. Nat. 2014, 13, 16–45. [Google Scholar]
- McClure, M. Biology and control of hemlock woolly adelgid. Bull. Conn. Agric. Exp. Stn 1987, 851, 1–9. [Google Scholar]
- Orwig, D.A.; Foster, D.R. Forest response to the introduced hemlock woolly adelgid in southern New England, USA. J. Torrey Bot. Soc. 1998, 125, 60–73. [Google Scholar] [CrossRef]
- Smith, W.B.; Miles, P.D.; Perry, C.H.; Pugh, S.A. Forest Resources of the United States, 2007; General Technical Report WO-78; USDA Forest Service: Washington, DC, USA, 2009. [Google Scholar]
- Ellison, A.M.; Barker Plotkin, A.A.; Foster, D.R.; Orwig, D.A. Experimentally testing the role of foundation species in forests: The Harvard Forest Hemlock Removal Experiment. Methods Ecol. Evol. 2010, 1, 168–179. [Google Scholar] [CrossRef]
- Orwig, D.A.; Barker Plotkin, A.A.; Davidson, E.A.; Lux, H.; Savage, K.E.; Ellison, A.M. Foundation species loss affects vegetation structure more than ecosystem function in a northeastern USA forest. PeerJ 2013, 1, e41. [Google Scholar] [CrossRef] [PubMed]
- Ellison, A.M. Reprise: Eastern hemlock as a foundation species. In Hemlock: A Forest Giant on the Edge; Foster, D.R., Ed.; Yale University Press: New Haven, CT, USA, 2014; pp. 165–171. [Google Scholar]
- Lustenhouwer, M.N.; Nicoll, L.; Ellison, A.M. Microclimatic effects of the loss of a foundation species from New England forests. Ecosphere 2012, 3, 26. [Google Scholar] [CrossRef]
- Cobb, R.C.; Orwig, D.A.; Currie, S.J. Decomposition of green foliage in eastern hemlock forests of southern New England impacted by hemlock woolly adelgid infestations. Can. J. For. Res. 2006, 36, 1–11. [Google Scholar] [CrossRef]
- Tingley, M.W.; Orwig, D.A.; Field, R.; Motzkin, G. Avian response to removal of a forest dominant: Consequences of hemlock woolly adelgid infestations. J. Biogeogr. 2002, 29, 1505–1516. [Google Scholar] [CrossRef]
- Ellison, A.M.; Chen, J.; Díaz, D.; Kammerer-Burnham, C.; Lau, M. Changes in ant community structure and composition associated with hemlock decline in New England. In Proceedings of the 3rd Symposium on Hemlock Woolly Adelgid in the Eastern United States, Asheville, NC, USA, 1–3 February 2005; Onken, B., Reardon, R., Eds.; US Department of Agriculture, US Forest Service Forest Health Technology Enterprise Team: Morgantown, WV, USA, 2005; pp. 280–289. [Google Scholar]
- Dilling, C.; Lambdin, P.; Grant, J.; Buck, L. Insect guild structure associated with eastern hemlock in the southern Appalachians. Environ. Entomol. 2007, 36, 1408–1414. [Google Scholar] [CrossRef]
- Rohr, J.R.; Mahan, C.G.; Kim, K. Response of arthropod biodiversity to foundation species declines: The case of the eastern hemlock. For. Ecol. Manag. 2009, 258, 1503–1510. [Google Scholar] [CrossRef]
- Mathewson, B.G. Eastern red-backed salamander relative abundance in eastern hemlock-dominated and mixed deciduous forests at the Harvard Forest. Northeast. Nat. 2009, 16, 1–12. [Google Scholar] [CrossRef]
- Porter, T.M.; Skillman, J.E.; Moncalvo, J.M. Fruiting body and soil rDNA sampling detects complementary assemblage of Agaricomycotina (Basidiomycota, Fungi) in a hemlock-dominated forest plot in southern Ontario. Mol. Ecol. 2008, 17, 3037–3050. [Google Scholar] [CrossRef] [PubMed]
- Baird, R.E.; Watson, C.E.; Woolfok, S. Microfungi from bark of healthy and damaged American beech, Fraser fir, and eastern hemlock trees during an all taxa biodiversity inventory in forests of the Great Smoky Mountains National Park. Southeast. Nat. 2007, 6, 67–82. [Google Scholar] [CrossRef]
- Baird, R.E.; Woolfok, S.; Watson, C.E. Microfungi of forest litter from healthy American beech, Fraser fir, and eastern hemlock stands in Great Smoky Mountains National Park. Southeast. Nat. 2009, 8, 609–630. [Google Scholar] [CrossRef]
- Catovsky, S.; Bazzaz, F. The role of resource interactions and seedling regeneration in maintaining a positive feedback in hemlock stands. J. Ecol. 2000, 88, 100–112. [Google Scholar] [CrossRef]
- Sullivan, K.A.; Ellison, A.M. The seed bank of hemlock forests: Implications for forest regeneration following hemlock decline. J. Torrey Bot. Soc. 2006, 133, 393–402. [Google Scholar] [CrossRef]
- Farnsworth, E.J.; Barker Plotkin, A.A.; Ellison, A.M. The relative contributions of seed bank, seed rain, and understory vegetation dynamics to the reorganization of Tsuga canadensis forests after loss due to logging or simulated attack by Adelges tsugae. Can. J. For. Res. 2012, 42, 2090–2105. [Google Scholar] [CrossRef]
- Ellison, A.M.; Lavine, M.; Kerson, P.B.; Barker Plotkin, A.A.; Orwig, D.A. Building a foundation: Land-use history and dendrochronology reveal temporal dynamics of a Tsuga canadensis (Pinaceae) forest. Rhodora 2014, 116, 377–427. [Google Scholar] [CrossRef]
- Albani, M.; Moorcroft, P.R.; Ellison, A.M.; Orwig, D.A.; Foster, D.R. Predicting the impact of hemlock woolly adelgid on carbon dynamics of eastern US forests. Can. J. For. Res. 2010, 40, 119–133. [Google Scholar] [CrossRef]
- Stadler, B.; Müller, T.; Orwig, D. The ecology of energy and nutrient fluxes in hemlock forests invaded by hemlock woolly adelgid. Ecology 2006, 87, 1792–1804. [Google Scholar] [CrossRef]
- Stadler, B.; Müller, T.; Orwig, D.; Cobb, R. Hemlock woolly adelgid in New England forests: Transforming canopy impacts transforming ecosystem processes and landscapes. Ecosystems 2005, 8, 233–247. [Google Scholar] [CrossRef]
- Haines, A. Flora Novae Angliae: A Manual for the Identification of Native and Naturalized Vascular Plants of New England; Yale University Press: New Haven, CT, USA, 2011. [Google Scholar]
- Chao, A.; Jost, L. Estimating diversity and entropy profiles via discovery rates of new species. Methods Ecol. Evol. 2015, 6, 873–882. [Google Scholar] [CrossRef]
- Chao, A.; Jost, L.; Chiang, S.C.; Jiang, Y.H.; Chazdon, R. A two-stage probabilistic approach to multiple-community similarity indices. Biometrics 2008, 64, 1178–1186. [Google Scholar] [CrossRef] [PubMed]
- Jost, L. GST and its relatives do not measure differentiation. Mol. Ecol. 2008, 17, 4015–4026. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; Version 3.2.2; R Foundation for Statistical Computing: Vienna, Austria, 2015; Available online: http://r-project.org/ (accessed on 1 July 2015).
- Chao, A.; Ma, K.H.; Hsieh, T.C. SpadeR: Species Prediction and Diversity Estimation with R, Version 0.1.0. Available online: http://chao.stat.nthu.edu.tw/blog/software-download/ (accessed on 18 October 2015).
- Ellison, A.; Plotkin, B.A. Understory Vegetation in Hemlock Removal Experiment at Harvard Forest Since 2003. Harvard Forest Data Archive: HF106. 2005. Available online: http://harvardforest.fas. harvard.edu:8080/exist/apps/datasets/showData.html?id=hf106 (accessed on 3 November 2015).
- Kendrick, J.A.; Ribbons, R.R.; Classen, A.T.; Ellison, A.M. Changes in canopy structure and ant assemblages affect soil ecosystem variables as a foundation species declines. Ecosphere 2015, 6, 77. [Google Scholar] [CrossRef]
- Oosting, H.J.; Bordeau, P.F. Virgin hemlock forest segregates in the Joyce Kilmer Memorial Forest of western North Carolina. Bot. Gaz. 1955, 116, 340–359. [Google Scholar] [CrossRef]
- Bieri, R.; Anliot, S.F. The structure and floristic composition of a virgin hemlock forest in West Virginia. Castanea 1965, 30, 205–226. [Google Scholar]
- Martin, K.L.; Goebel, P.C. The foundation species influence of eastern hemlock (Tsuga canadensis) on biodiversity and ecosystem function on the unglaciated Allegheny Plateau. For. Ecol. Manag. 2013, 289, 143–152. [Google Scholar] [CrossRef]
- Abella, S.R.; Shelburne, V.B. Ecological species groups of South Carolina’s Jocassee Gorges, Southern Applachian Mountains. J. Torrey Bot. Soc. 2004, 131, 200–222. [Google Scholar] [CrossRef]
- Cleavitt, N.L.; Eschtruth, A.K.; Battles, J.J.; Fahey, T.J. Bryophyte response to eastern hemlock decline casued by hemlock woolly adelgid infestation. J. Torrey Bot. Soc. 2008, 135, 12–25. [Google Scholar] [CrossRef]
- Eschtruth, A.K.; Cleavitt, N.L.; Battles, J.J.; Evans, R.A.; Fahey, T.J. Vegetation dynamics in declining eastern hemlock stands: 9 years of forest response to hemlock woolly adelgid infestation. Can. J. For. Res. 2006, 36, 1435–1450. [Google Scholar] [CrossRef]
- Eschtruth, A.K.; Battles, J.J. Acceleration of exotic plant invasion in a forested ecosystem by a generalist herbivore. Conserv. Biol. 2009, 23, 388–399. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ellison, A.M.; Barker Plotkin, A.A.; Khalid, S. Foundation Species Loss and Biodiversity of the Herbaceous Layer in New England Forests. Forests 2016, 7, 9. https://doi.org/10.3390/f7010009
Ellison AM, Barker Plotkin AA, Khalid S. Foundation Species Loss and Biodiversity of the Herbaceous Layer in New England Forests. Forests. 2016; 7(1):9. https://doi.org/10.3390/f7010009
Chicago/Turabian StyleEllison, Aaron M., Audrey A. Barker Plotkin, and Shah Khalid. 2016. "Foundation Species Loss and Biodiversity of the Herbaceous Layer in New England Forests" Forests 7, no. 1: 9. https://doi.org/10.3390/f7010009
APA StyleEllison, A. M., Barker Plotkin, A. A., & Khalid, S. (2016). Foundation Species Loss and Biodiversity of the Herbaceous Layer in New England Forests. Forests, 7(1), 9. https://doi.org/10.3390/f7010009