
Optimization Forest Thinning Measures for Carbon Budget in a Mixed Pine-Oak Stand of the Qingling Mountains, China: A Case Study

Abstract
:1. Introduction
2. Material and Methods
2.1. Site Description
2.2. Experimental Design
2.3. Thinning, Residual Removal, and Dynamics of Tree Growth and Litterfall
2.4. Soil CO2 efflux
2.5. Chemical Analysis
2.6. Data Processing and Analysis
3. Results
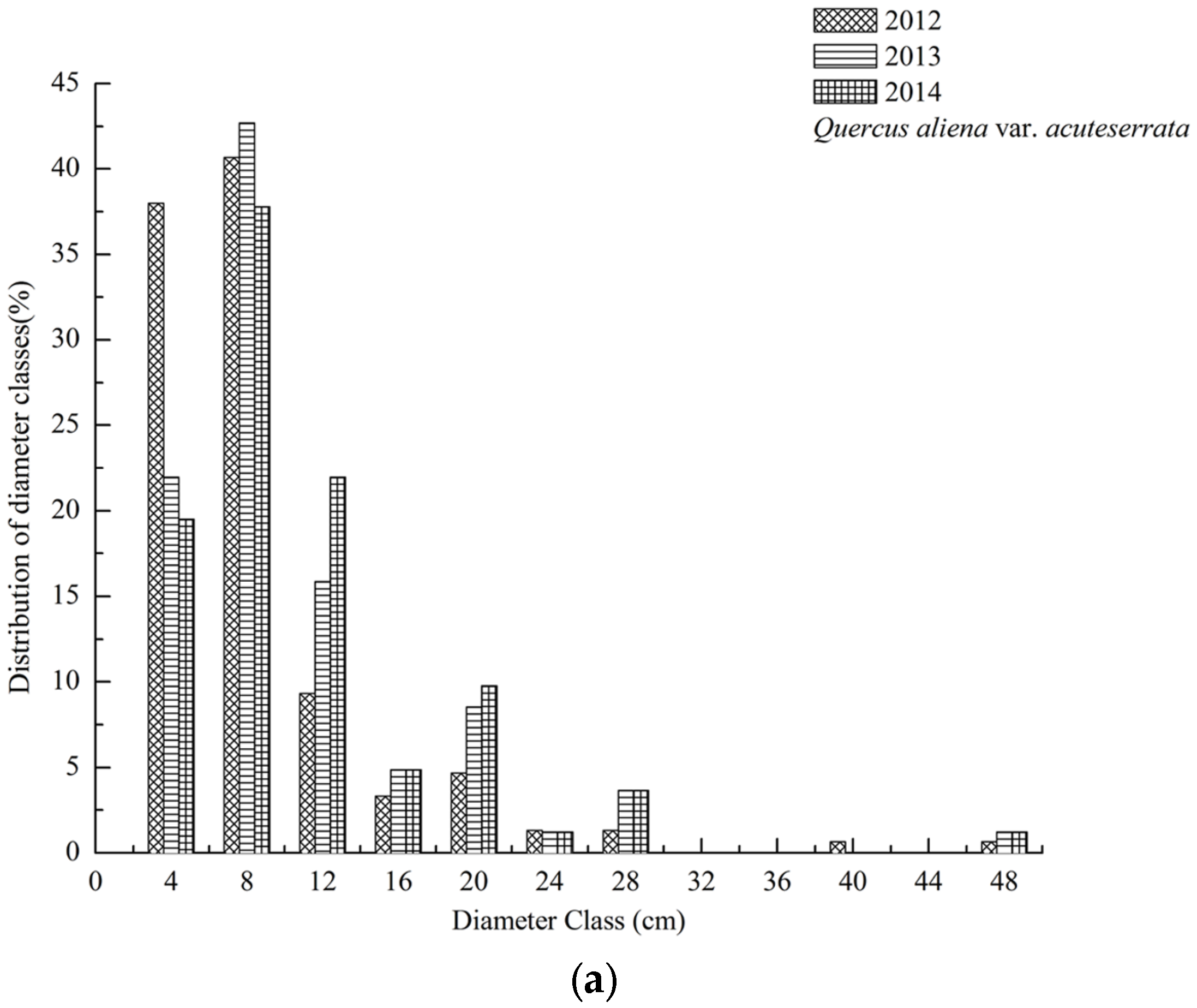
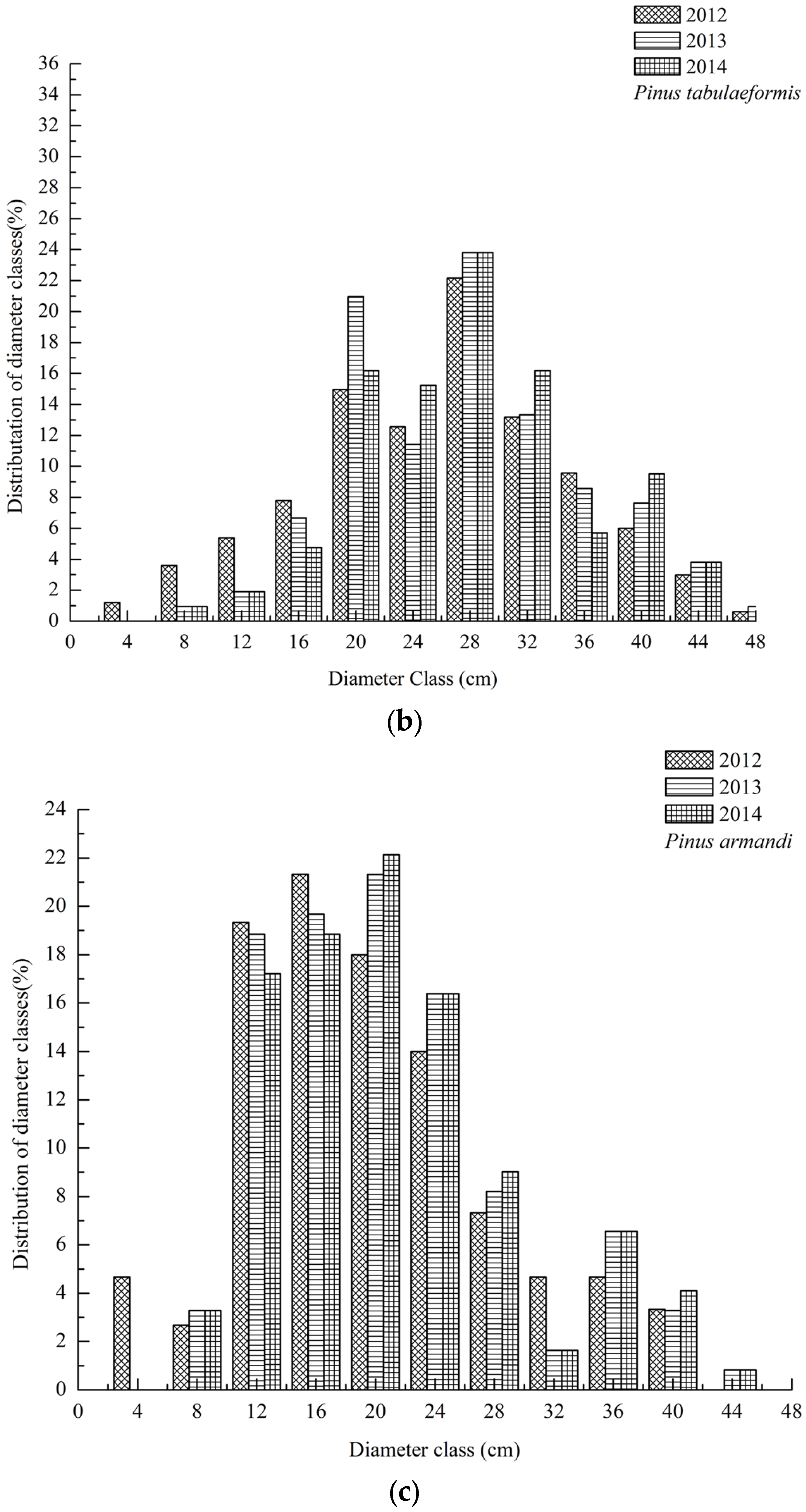
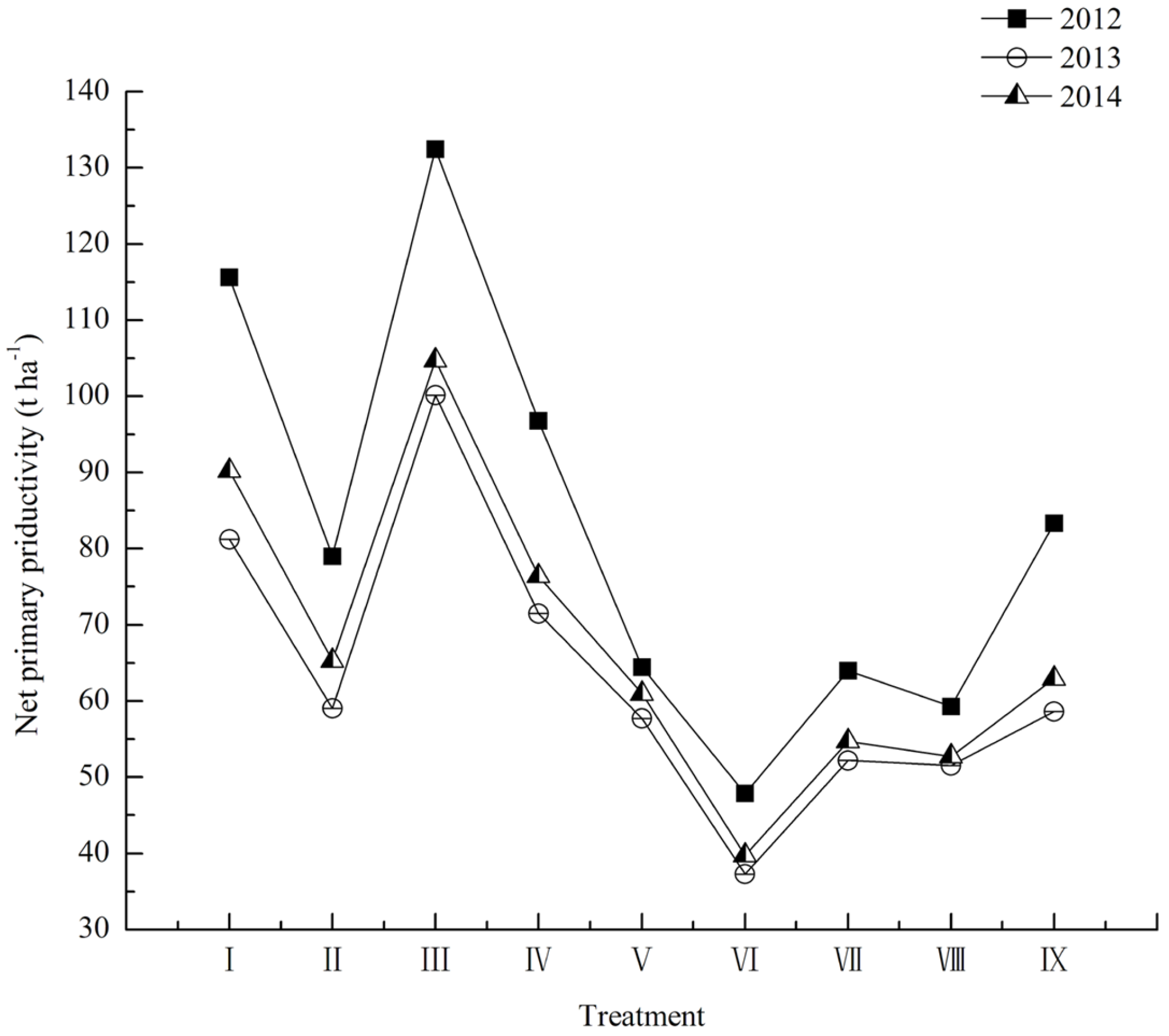
3.1. Dynamics of Tree Species Composition
3.2. DBH Dynamics of Tree Species
3.3. Dynamics of Net Primary Productivity of Living Trees
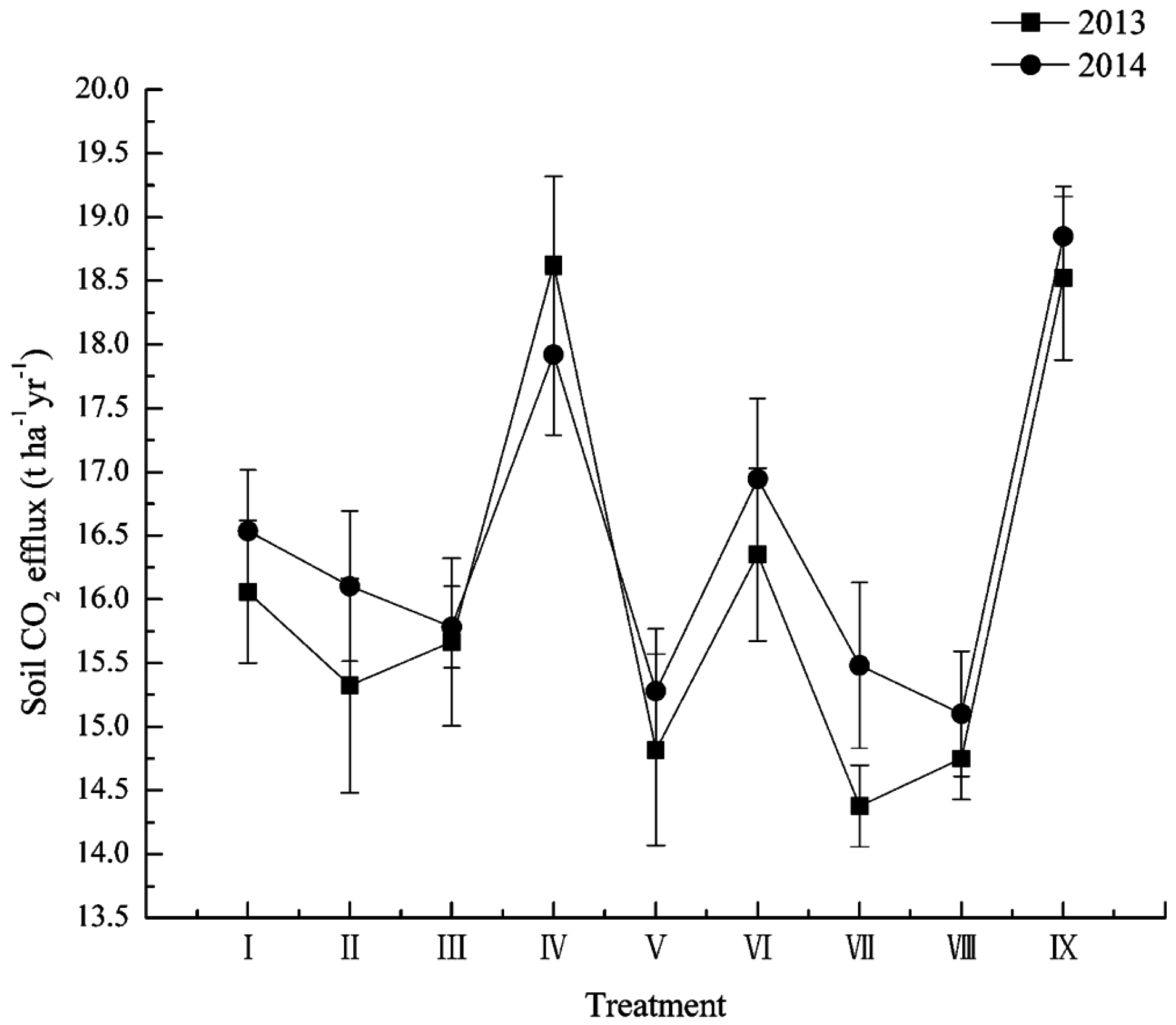
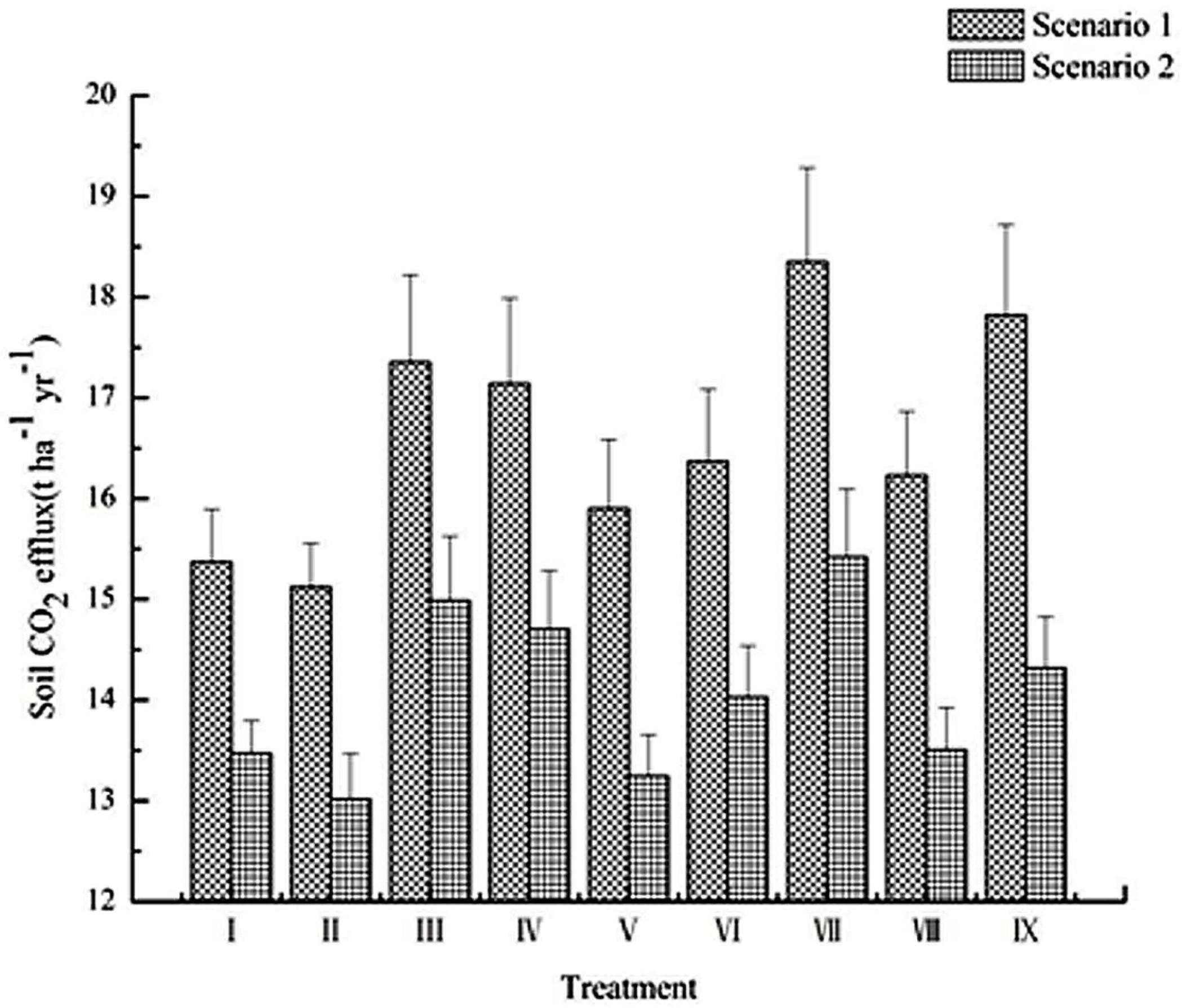
3.4. Soil CO2 Efflux Stimulation
3.5. Model Fitting
3.6. Effects of Forest Management on Carbon Budget
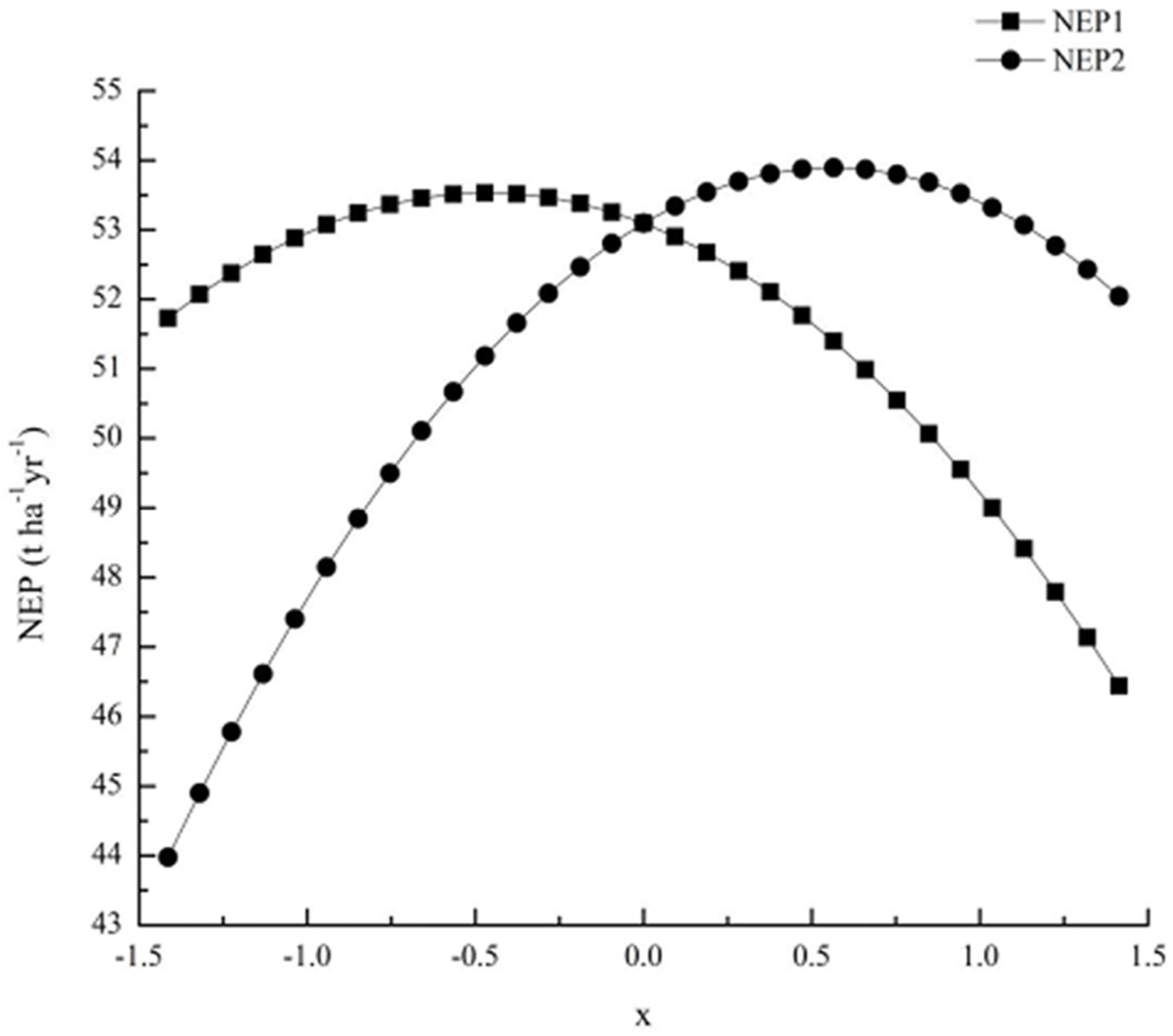
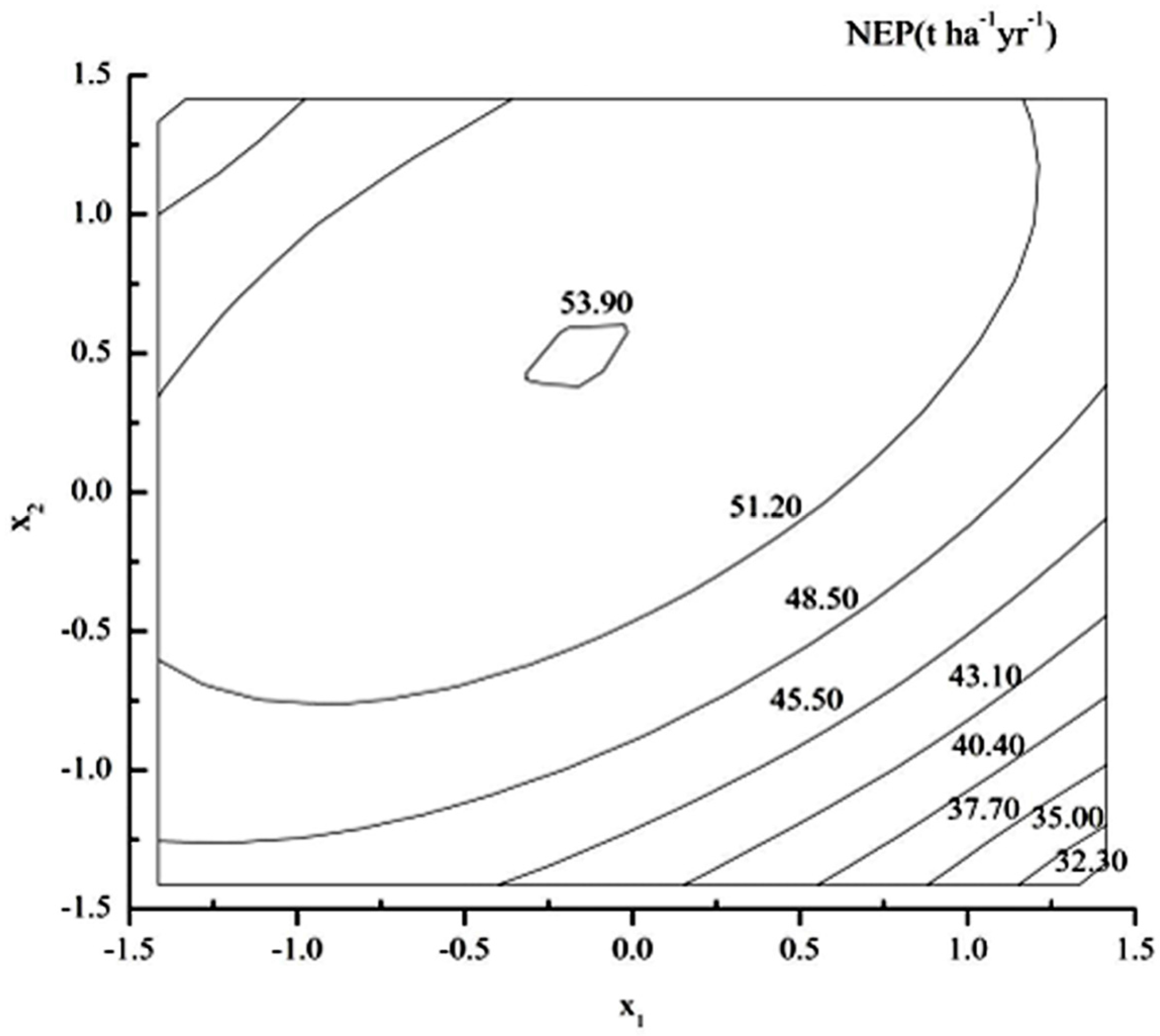
3.7. Optimization Forest Management Measures for NEP
4. Discussion
4.1. Effect of Chemical Compound Groups of Litterfall on Soil CO2 Efflux
4.2. The Response of Soil CO2 Efflux to Management Measures
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Magruder, M.; Chhin, S.; Palik, B.; Bradford, J.B. Thinning increases climatic resilience of red pine. Can. J. For. Res. 2013, 43, 878–889. [Google Scholar] [CrossRef]
- Verschuyl, J.; Riffell, S.; Miller, D.; Wigley, T.B. Biodiversity response to intensive biomass production from forest thinning in north American forests—A meta-analysis. For. Ecol. Manag. 2011, 261, 221–232. [Google Scholar] [CrossRef]
- Boncina, A.; Kadunc, A.; Robic, D. Effects of selective thinning on growth and development of beech (Fagus sylvatica L.) forest stands in south-eastern Slovenia. Ann. For. Sci. 2007, 64, 47–57. [Google Scholar] [CrossRef]
- Rodríguez-Calcerrada, J.; Pérez-Ramos, I.M.; Ourcival, J.-M.; Limousin, J.-M.; Joffre, R.; Rambal, S. Is selective thinning an adequate practice for adapting Quercus ilex coppices to climate change? Ann. For. Sci. 2011, 68, 575–582. [Google Scholar] [CrossRef]
- Fernandez, I.; Älvarez-Gonzalez, J.G.; Carrasco, B.; Ruíz-González, A.D.; Cabaneiro, A. Post-thinning soil organic matter evolution and soil CO2 effluces in temperate radiata pine plantations: Impacts of moderate thinning regimes on the forest C cycle. Can. J. For. Res. 2012, 42, 1953–1964. [Google Scholar] [CrossRef]
- Dobarco, M.R.; Miegroet, H.V. Soil organic carbon storage and stability in the aspen-conifer ecotone in montane forests in Utah State, USA. Forests 2014, 5, 666–688. [Google Scholar] [CrossRef]
- Houghton, R.A. Revised estimates of the annual net flux of carbon to the atmosphere from changes in land use and land management 1850–2000. Tellus B 2003, 55, 378–390. [Google Scholar] [CrossRef]
- Chatterjee, A.; Vance, G.F.; Pendall, E.; Stahl, P.D. Timber harvesting alters soil carbon mineralization and microbial community structure in coniferous forests. Soil Biol. Biochem. 2008, 40, 1901–1907. [Google Scholar] [CrossRef]
- Diochon, A.; Kellman, L.; Beltrami, H. Looking deeper: An investigation of soil carbon losses following harvesting from a managed northeastern red spruce (Picea rubens Sarg.) forest chronosequence. For. Ecol. Manag. 2009, 257, 413–420. [Google Scholar] [CrossRef]
- Jassal, R.S.; Black, T.A.; Cai, T.; Morgenstern, K.; Li, Z.; Gaumont-Guay, D.; Nesic, Z. Components of ecosystem respiration and an estimate of net primary productivity of an intermediate-aged Douglas-fir stand. Agricu. For. Meteorol. 2007, 144, 44–57. [Google Scholar] [CrossRef]
- Mäkipää, R.; Linkosalo, T.; Komarov, A.; Mäkelä, A. Mitigation of climate change with biomass harvesting in Norway spruce stands: Are harvesting practices carbon neutral? Can. J. For. Res. 2014, 45, 1–9. [Google Scholar] [CrossRef]
- User-Interface Software of Yasso07. Available online: http://www.syke.fi/en-US/Research__Development/Research_and_development_projects/Projects/Soil_carbon_model_Yasso/Download (accessed on 5 November 2016).
- Tuomi, M.; Rasinmäki, J.; Repo, A.; Vanhala, P.; Liski, J. Soil carbon model Yasso07 graphical user interface. Environ. Model. Softw. 2011, 26, 1358–1362. [Google Scholar] [CrossRef]
- Ryu, S.R.; Concilio, A.; Chen, J.; North, M.; Ma, S. Prescribed burning and mechanical thinning effects on belowground conditions and soil respiration in a mixed-conifer forest, California. For. Ecol. Manag. 2009, 257, 1324–1332. [Google Scholar] [CrossRef]
- Sullivan, B.W.; Kolb, T.E.; Hart, S.C.; Kaye, J.P.; Dore, S.; Montes-Helu, M. Thinning reduces soil carbon dioxide but not methane flux from southwestern USA Ponderosa pine forests. For. Ecol. Manag. 2008, 255, 4047–4055. [Google Scholar] [CrossRef]
- Campbell, J.; Alberti, G.; Martin, J.; Law, B.E. Carbon dynamics of a ponderosa pine plantation following a thinning treatment in the northern Sierra Nevada. For. Ecol. Manag. 2009, 257, 453–463. [Google Scholar] [CrossRef]
- Kobziar, L.N.; Stephens, S.L. The effects of fuels treatments on soil carbon respiration in a Sierra Nevada pine plantation. Agricu. For. Meteorol. 2006, 141, 161–178. [Google Scholar] [CrossRef]
- Kobziar, L.N. The role of environmental factors and tree injuries in soil carbon respiration response to fire and fuels treatments in pine plantations. Biogeochemistry 2007, 84, 191–206. [Google Scholar] [CrossRef]
- Saunders, M.; Tobin, B.; Black, K.; Gioria, M.; Nieuwenhuis, M.; Osborne, B.A. Thinning effects on the net ecosystem carbon exchange of a Sitka spruce forest are temperature-dependent. Agric. For. Meteorol. 2012, 157, 1–10. [Google Scholar] [CrossRef]
- Hopmans, P.; Elms, S.R. Changes in total carbon and nutrients in soil profiles and accumulation in biomass after a 30-year rotation of Pinus radiata on podzolized sands: Impacts of intensive harvesting on soil resources. For. Ecol. Manag. 2009, 258, 2183–2193. [Google Scholar] [CrossRef]
- Tang, Z.; Fang, J. Temperature variation along the northern and southern slopes of Mt. Taibai, China. Agric. For. Meteorol. 2006, 139, 200–207. [Google Scholar] [CrossRef]
- Oyejola, B.A.; Nwanya, J.C. Selecting the right central composite design. Int. J. Stat. Appl. 2015, 5, 21–30. [Google Scholar]
- Khataee, A.R.; Fathinia, M.; Aber, S.; Zarei, M. Optimization of photocatalytic treatment of dye solution on supported tio2 nanoparticles by central composite design: Intermediates identification. J. Hazard. Mater. 2010, 181, 886–897. [Google Scholar] [CrossRef] [PubMed]
- Kasiri, M.B.; Aleboyeh, H.; Aleboyeh, A. Modeling and optimization of heterogeneous photo-fenton process with response surface methodology and artificial neural networks. Environ. Sci. Technol. 2008, 42, 7970–7975. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.Y. Data Processing System-Experimental Design, Statistical Analysis and Data Mining, 2nd ed.; Science Press: Beijing, China, 2010. [Google Scholar]
- Regulation for tending of forest. GB/T 15781 2009. Available online: http://www.doc88.com/p-94457076273.html (accessed on 5 November 2016).
- Post, W.M.; Emanuel, W.R.; Zinke, P.J.; Stangenberger, A.G. Soil carbon pools and world life zones. Nature 1982, 298, 156–159. [Google Scholar] [CrossRef]
- Tuomi, M.; Laiho, R.; Repo, A.; Liski, J. Wood decomposition model for boreal forests. Ecol. Model. 2011, 222, 709–718. [Google Scholar] [CrossRef]
- Parton, W.J.; Schimel, D.S.; Cole, C.V.; Ojima, D.S. Analysis of factors controlling soil organic matter levels in great plains grasslands. Soil Sci. Soc. Am. J. 1987, 51, 1173–1179. [Google Scholar] [CrossRef]
- Rolff, C.; Ågren, G.I. Predicting effects of different harvesting intensities with a model of nitrogen limited forest growth. Ecol. Model. 1999, 118, 193–211. [Google Scholar] [CrossRef]
- Chertov, O.G.; Komarov, A.S.; Nadporozhskaya, M.; Bykhovets, S.S.; Zudin, S.L. Romul—A model of forest soil organic matter dynamics as a substantial tool for forest ecosystem modeling. Ecol. Model. 2001, 138, 289–308. [Google Scholar] [CrossRef]
- Coleman, K.; Jenkinson, D.S. Rothc-26.3—A Model for the Turnover of Carbon in Soil; Springer: Heidelberge, Germany, 1996; pp. 237–246. [Google Scholar]
- Wallman, P.; Belyazid, S.; Svensson, M.G.E.; Sverdrup, H. Decomp—A semi-mechanistic model of litter decomposition. Environ. Model. Softw. 2006, 21, 33–44. [Google Scholar] [CrossRef]
- Tuomi, M.; Thum, T.; Järvinen, H.; Fronzek, S.; Berg, B.; Harmon, M.; Trofymow, J.A.; Sevanto, S.; Liski, J. Leaf litter decomposition—Estimates of global variability based on Yasso07 model. Ecol. Model. 2009, 220, 3362–3371. [Google Scholar] [CrossRef]
- Bai, L.L.; Li, Y.; Hou, L.; Luo, C.L.; Geng, Z.C.; Cheng, H.F. Chemical composition of litters from main forests in qinling mountains. J. Northwest A F Univ. (Nat. Sci. Ed.) 2016, 44, 89–96. [Google Scholar]
- Pan, Y.; Luo, T.; Birdsey, R.; Hom, J.; Melillo, J. New estimates of carbon storage and sequestration in China’s forests: Effects of age-class and method on inventory-based carbon estimation. Clim. Chang. 2004, 67, 211–236. [Google Scholar] [CrossRef]
- Chen, C.; Peng, H. Standing crops and productivity of the major forest-types at the Huoditang forest region of the Qinling Mountains. J. Northwest For. Coll. 1996, 11, 92–102. [Google Scholar]
- Valachovic, Y.S.; Caldwell, B.A.; Cromack, K.; Griffiths, R.P. Leaf litter chemistry controls on decomposition of Pacific northwest trees and woody shrubs. Can. J. For. Res. 2004, 34, 2131–2147. [Google Scholar] [CrossRef]
- Sariyildiz, T.; Anderson, J.M.; Kucuk, M. Effects of tree species and topography on soil chemistry, litter quality, and decomposition in northeast Turkey. Soil Biol. Biochem. 2005, 37, 1695–1706. [Google Scholar] [CrossRef]
- Berg, B. Litter decomposition and organic matter turnover in northern forest soils. For. Ecol. Manag. 2000, 133, 13–22. [Google Scholar] [CrossRef]
- Davidson, E.A.; Elizabeth, B.; Boone, R.D. Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest. Glob. Chang. Biol. 1998, 4, 217–227. [Google Scholar] [CrossRef]
- Epron, D.; Farque, L.; Lucot, É.; Badot, P.M. Soil CO2 efflux in a beech forest: Dependence on soil temperature and soil water content. Ann. For. Sci. 1999, 56, 221–226. [Google Scholar] [CrossRef]
- Burton, A.J.; Pregitzer, K.S. Field measurements of root respiration indicate little to no seasonal temperature acclimation for sugar maple and red pine. Tree Physiol. 2003, 23, 273–280. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot No. | Gradient (°) | Canopy Density | Tree Species Composition | Soil Depth (cm) | |||
|---|---|---|---|---|---|---|---|
| 2012 | 2013 | 2012 | 2013 | 2014 | |||
| 1 | 35 | 0.75 | 0.72 | 7 Pt 2 Pa 1 Q | 9 Pa 1 Pt | 9 Pa 1 Pt | 30 |
| 2 | 30 | 0.8 | 0.73 | 5 Pt 3 Pa 2 Q | 5 Pa 1 Pt 4 Q | 5 Pa 1 Pt 4 Q | 45 |
| 3 | 30 | 0.8 | 0.72 | 7 Pt 2 Pa 1 Q | 7 Pa 2 Pt 1 Q | 7 Pa 2 Pt 1 Q | 40 |
| 4 | 30 | 0.8 | 0.73 | 6 Pt 2 Pa 2 Q | 2 Pa 4 Pt 4 Q | 2 Pa 4 Pt 4 Q | 50 |
| 5 | 25 | 0.85 | 0.81 | 4 Pt 3 Pa 3 Q | 1 Pa 5 Pt 4 Q | 1 Pa 5 Pt 4Q | 46 |
| 6 | 30 | 0.8 | 0.7 | 1 Pt 7 Pa 2 Q | 1 Pa 8 Pt 1 Q | 1 Pa 8 Pt 1 Q | 35 |
| 7 | 30 | 0.75 | 0.73 | 2 Pt 7 Pa 1 Q | 8 Pt 2 Q | 8 Pt 2 Q | 40 |
| 8 | 30 | 0.9 | 0.81 | 2 Pt 5 Pa 3 Q | 2 Pa 5 Pt 3 Q | 2 Pa 5 Pt 3 Q | 44 |
| 9 | 25 | 0.9 | 0.84 | 2 Pt 7 Pa 1 Q | 1 Pa 7 Pt 2 Q | 1 Pa 7 Pt 2 Q | 33 |
| 10 | 30 | 0.8 | 0.72 | 1 Pa 8 Pt 1 Q | 5 Pa 1 Pt 4 Q | 6 Pa 4 Q | 38 |
| 11 | 30 | 0.9 | 0.83 | 6 Pt 2 Pa 2 Q | 1 Pa 3 Pt 6 Q | 1 Pa 3 Pt 6 Q | 41 |
| 12 | 25 | 0.8 | 0.72 | 6 Pt 3 Pa 1 Q | 7 Pa 2 Pt 1 Q | 8 Pa 2 Q | 45 |
| 13 | 28 | 0.8 | 0.73 | 8 Pt 1 Pa 1 Q | 9 Pa 1 Q | 8 Pa 1 Pt 1 Q | 45 |
| Variables | Levels | ||||
|---|---|---|---|---|---|
| (−α) − 1.414 | −1 | 0 | 1 | (+α) + 1.414 | |
| X1 (%) | 5.00 | 7.93 | 15.00 | 22.07 | 25.00 |
| X2 (%) | 0.00 | 14.64 | 50.00 | 85.36 | 100.00 |
| Plot No. | Treatment | Design Code | Thinning Factors | Block | ||
|---|---|---|---|---|---|---|
| x1 | x2 | X1 (%) | X2 (%) | |||
| 1 | I | 1 | 1 | 22.07 | 85.36 | Factorial points |
| 2 | II | 1 | −1 | 22.07 | 14.64 | |
| 3 | III | −1 | 1 | 7.93 | 85.36 | |
| 4 | IV | −1 | −1 | 7.93 | 14.64 | |
| 5 | V | −α | 0 | 5 | 50 | Axial points |
| 6 | VI | +α | 0 | 25 | 50 | |
| 7 | VII | 0 | −α | 15 | 0 | |
| 8 | VIII | 0 | +α | 15 | 100 | |
| 9 | IX | 0 | 0 | 15 | 50 | Center points |
| 10 | IX | 0 | 0 | 15 | 50 | |
| 11 | IX | 0 | 0 | 15 | 50 | |
| 12 | IX | 0 | 0 | 15 | 50 | |
| 13 | IX | 0 | 0 | 15 | 50 | |
| Diameter Class (cm) | DBH (cm) |
|---|---|
| 4 | ≤6 |
| 8 | 6.1 ≤ DBH ≤ 10 |
| 12 | 10.1 ≤ DBH ≤ 14 |
| 16 | 14.1 ≤ DBH ≤ 18 |
| 20 | 18.1 ≤ DBH ≤ 22 |
| 24 | 22.1 ≤ DBH ≤ 26 |
| 28 | 26.1 ≤ DBH ≤ 30 |
| 32 | 30.1 ≤ DBH ≤ 34 |
| 36 | 34.1 ≤ DBH ≤ 38 |
| 40 | 38.1 ≤ DBH ≤ 42 |
| 44 | 42.1 ≤ DBH ≤ 46 |
| 48 | 46.1 ≤ DBH ≤ 50 |
| Parameter | Degrees of Freedom | Sum of Squares | Mean Square | F-Value | Pr > F |
|---|---|---|---|---|---|
| x1 | 1 | 27.9913 | 27.9913 | 5.5179 | 0.0512 |
| x2 | 1 | 65.1148 | 65.1148 | 12.8360 | 0.0089 |
| x12 | 1 | 27.9793 | 27.9793 | 5.5155 | 0.0512 |
| x22 | 1 | 44.9868 | 44.9868 | 8.8682 | 0.0206 |
| x1x2 | 1 | 24.5520 | 24.5520 | 4.8399 | 0.0637 |
| Model | 5 | 182.4717 | 36.4943 | 7.1941 | 0.0226 |
| Lack of fit | 3 | 13.3448 | 4.4483 | 0.8028 | 0.5308 |
| Residual | 7 | 35.5097 | 5.0728 | ||
| Error | 4 | 22.1649 | 5.5412 | ||
| Total | 12 | 217.9815 | |||
| R2 = 0.837 | |||||
| Adjusted R2 = 0.849 | |||||
| Tree Species | Ethanol | Ethanol Std | Water | Water Std | Acid | Acid Std | Non Soluble | Non Soluble Std | Litter Types |
|---|---|---|---|---|---|---|---|---|---|
| Pt | 0.125a | 0.004 | 0.153b | 0.006 | 0.433w | 0.003 | 0.289c | 0.006 | Leaf/needle |
| Pa | 0.122a | 0.005 | 0.151b | 0.006 | 0.418w | 0.005 | 0.309c | 0.011 | Leaf/needle |
| Q | 0.199b | 0.004 | 0.252c | 0.004 | 0.445w | 0.005 | 0.104d | 0.002 | Leaf/needle |
| Pt | 0.120a | 0.005 | 0.142d | 0.002 | 0.477w | 0.003 | 0.261e | 0.007 | Fine root |
| Pa | 0.119a | 0.002 | 0.130ad | 0.002 | 0.476w | 0.002 | 0.276e | 0.003 | Fine root |
| Q | 0.154c | 0.003 | 0.187e | 0.009 | 0.429w | 0.009 | 0.229f | 0.015 | Fine root |
| Pt | 0.086k | 0.004 | 0.088kf | 0.004 | 0.525m | 0.027 | 0.301c | 0.005 | Twig |
| Pa | 0.061e | 0.006 | 0.097f | 0.005 | 0.523m | 0.005 | 0.321c | 0.005 | Twig |
| Q | 0.038u | 0.002 | 0.081f | 0.008 | 0.562m | 0.005 | 0.319c | 0.32 | Twig |
| Pt | 0.073g | 0.002 | 0.091f | 0.003 | 0.530m | 0.002 | 0.306c | 0.004 | Coarse root |
| Pa | 0.072g | 0.004 | 0.085f | 0.004 | 0.524m | 0.004 | 0.319c | 0.006 | Coarse root |
| Q | 0.111h | 0.003 | 0.139g | 0.013 | 0.473w | 0.007 | 0.278f | 0.007 | Coarse root |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hou, L.; Li, Z.; Luo, C.; Bai, L.; Dong, N. Optimization Forest Thinning Measures for Carbon Budget in a Mixed Pine-Oak Stand of the Qingling Mountains, China: A Case Study. Forests 2016, 7, 272. https://doi.org/10.3390/f7110272
Hou L, Li Z, Luo C, Bai L, Dong N. Optimization Forest Thinning Measures for Carbon Budget in a Mixed Pine-Oak Stand of the Qingling Mountains, China: A Case Study. Forests. 2016; 7(11):272. https://doi.org/10.3390/f7110272
Chicago/Turabian StyleHou, Lin, Zhe Li, Chunlin Luo, Longlong Bai, and Ningning Dong. 2016. "Optimization Forest Thinning Measures for Carbon Budget in a Mixed Pine-Oak Stand of the Qingling Mountains, China: A Case Study" Forests 7, no. 11: 272. https://doi.org/10.3390/f7110272
APA StyleHou, L., Li, Z., Luo, C., Bai, L., & Dong, N. (2016). Optimization Forest Thinning Measures for Carbon Budget in a Mixed Pine-Oak Stand of the Qingling Mountains, China: A Case Study. Forests, 7(11), 272. https://doi.org/10.3390/f7110272