
Heterotrophic Soil Respiration Affected by Compound Fertilizer Types in Red Pine (Pinus densiflora S. et Z.) Stands of Korea



Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Stand and Soil Characteristics
2.3. Soil Respiration Rates
2.4. Data Analysis
3. Results
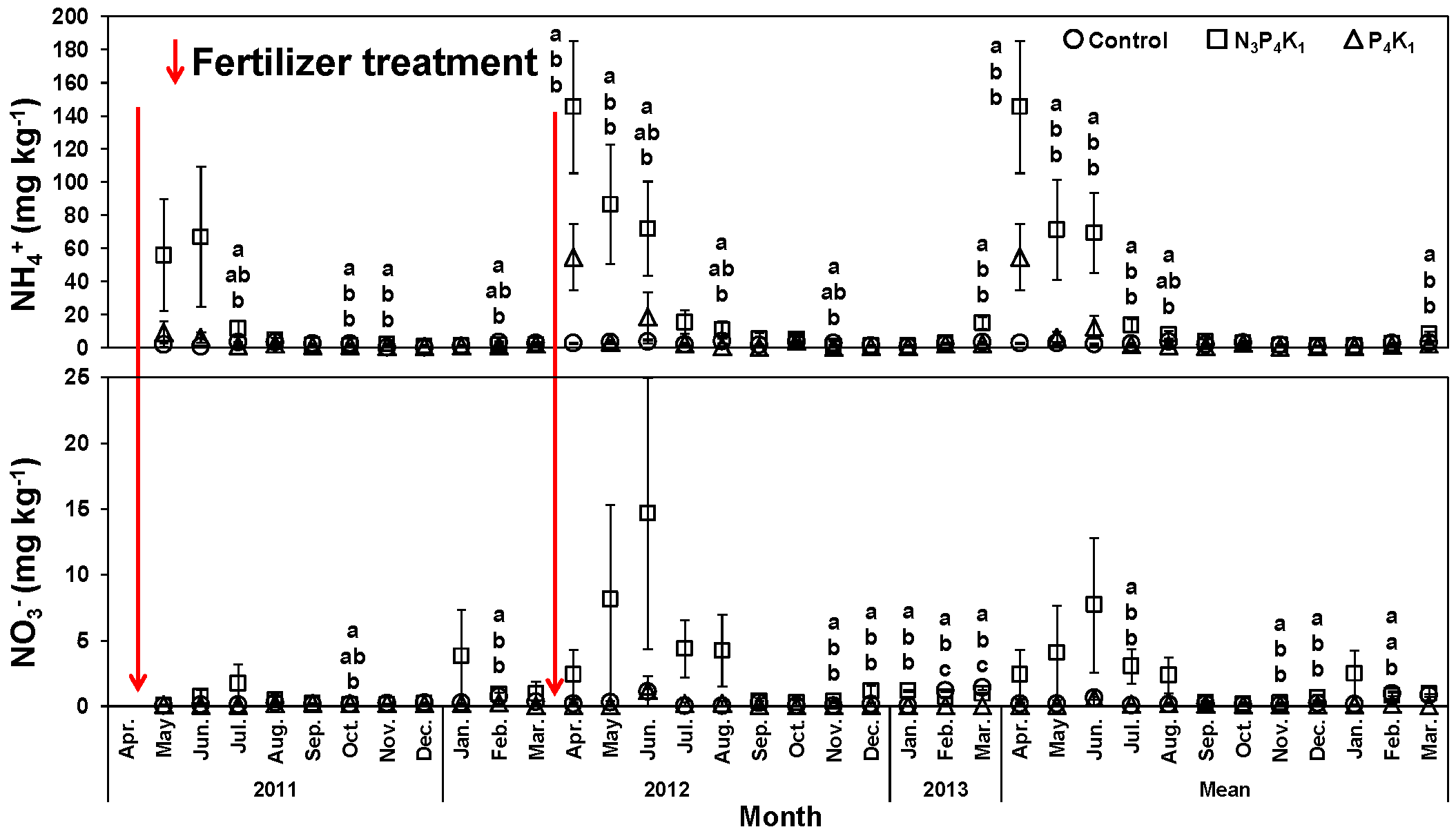
3.1. Stand and Soil Characteristics
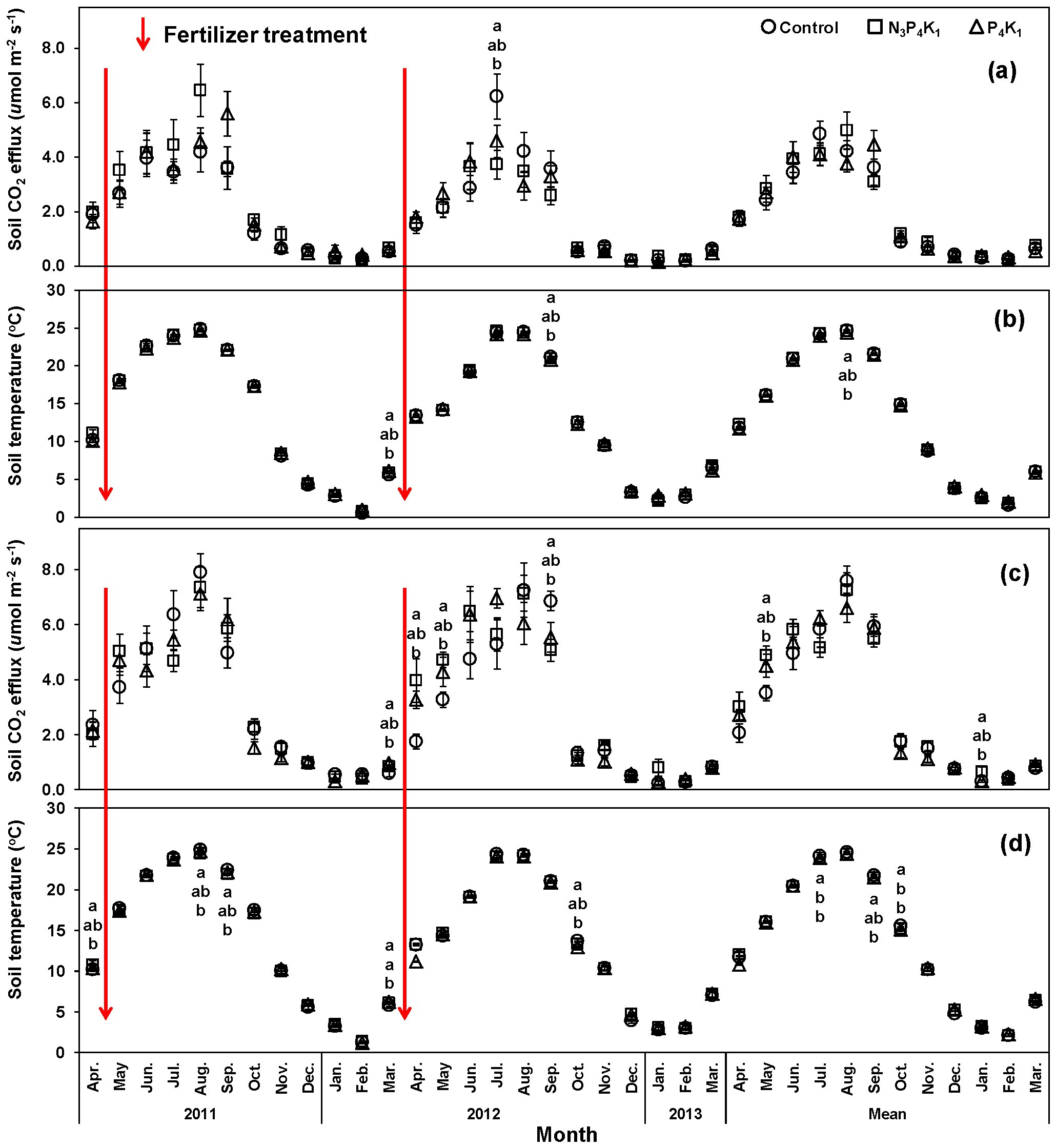
3.2. Monthly Variation of Rh and Rs Rates
3.3. Annual Rh and Rs Rates
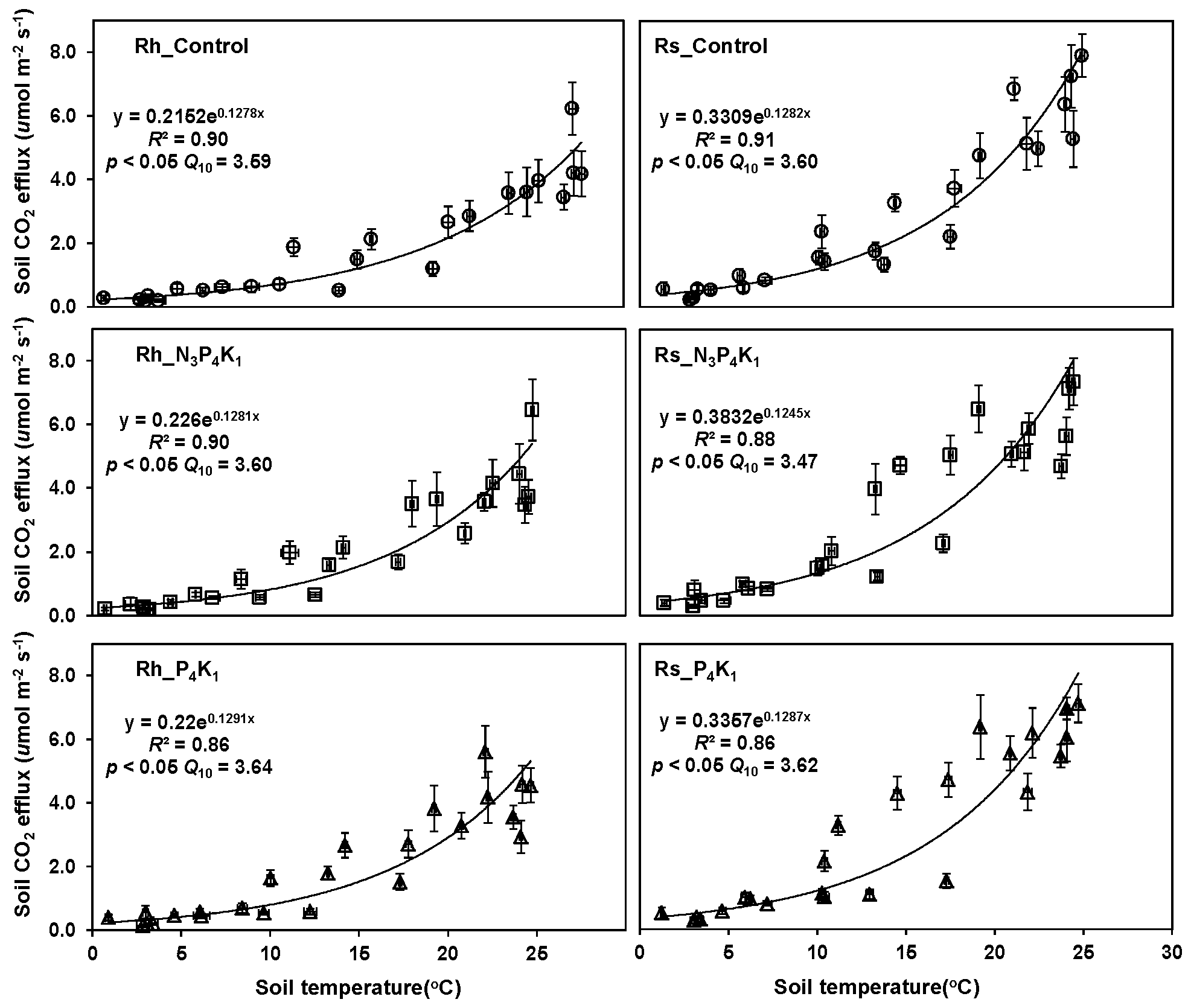
3.4. Temperature Dependency of Rh and Rs
3.5. Relationships between Rs and Rh Rates
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Jandl, R.; Lindner, M.; Vesterdal, L.; Bauwens, B.; Baritz, R.; Hagedorn, F.; Johnson, D.W.; Minkkinen, K.; Byrne, K.A. How strongly can forest management influence soil carbon sequestration? Geoderma 2007, 137, 253–268. [Google Scholar] [CrossRef]
- Kim, C. Soil carbon storage, litterfall and CO2 efflux in fertilized and unfertilized larch (Larix leptolepis) plantations. Ecol. Res. 2008, 23, 757–763. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, W.; Sun, T.; Chen, L.; Pang, X.; Wang, Y.; Xiao, F. N and P fertilization reduced soil autotrophic and heterotrophic respiration in young Cunninghamia lanceolate forest. Agric. For. Meteorol. 2017, 232, 66–73. [Google Scholar] [CrossRef]
- Hasselquist, N.J.; Metcalfe, D.B.; Högberg, P. Contrasting effects of low and high nitrogen additions on soil CO2 flux components and ectomycorrhizal fungal sporocarp production in a boreal forest. Glob. Chang. Biol. 2012, 18, 3596–3605. [Google Scholar] [CrossRef]
- Haynes, B.E.; Gower, S.T. Belowground carbon allocation in unfertilized and fertilized red pine plantations in northern Wisconsin. Tree Physiol. 1995, 15, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Olsson, P.; Linder, S.; Giesler, R.; Högberg, P. Fertilization of boreal forest reduces both autotrophic and heterotrophic soil respiration. Glob. Chang. Biol. 2005, 11, 1745–1753. [Google Scholar] [CrossRef]
- Phillips, R.P.; Fahey, T.J. Fertilization effects on fineroot biomass, rhizosphere microbes and respiratory fluxes in hardwood forest soils. New Phytol. 2007, 176, 655–664. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.H.; Jose, S. Soil respiration, fine root production, and microbial biomass in cottonwood and loblolly pine plantations along a nitrogen fertilization gradient. For. Ecol. Manag. 2003, 185, 263–273. [Google Scholar] [CrossRef]
- Raich, J.W.; Tufekciogul, A. Vegetation and soil respiration: Correlations and controls. Biogeochemistry 2000, 48, 71–90. [Google Scholar] [CrossRef]
- He, T.; Wang, Q.; Wang, S.; Zhang, F. Nitrogen addition altered the effect of belowground C allocation on soil respiration in a subtropical forest. PLoS ONE 2016, 11, e0155881. [Google Scholar] [CrossRef] [PubMed]
- Samuelson, L.; Mathew, R.; Stokes, T.; Feng, Y.; Aubrey, D.; Coleman, M. Soil and microbial respiration in a loblolly pine plantation in response to seven years of irrigation and fertilization. For. Ecol. Manag. 2009, 258, 2431–2438. [Google Scholar] [CrossRef]
- Franklin, O.; Högberg, P.; Ekblad, A.; Ågren, G.I. Pine forest floor carbon accumulation in response to N and PK additions: Bomb 14C modelling and respiration studies. Ecosystems 2003, 6, 644–658. [Google Scholar] [CrossRef]
- Hwang, J.; Son, Y.; Kim, C.; Yi, M.J.; Kim, Z.S.; Lee, W.K.; Hong, S.K. Fine root dynamics in thinned and limed pitch pine and Japanese larch plantations. J. Plant Nutr. 2007, 30, 1821–1839. [Google Scholar] [CrossRef]
- Kim, C.; Jeong, J.; Park, J.H.; Ma, H.S. Growth and nutrient status of foliage as affected by tree species and fertilization in a fire-disturbed urban forest. Forests 2015, 6, 2199–2213. [Google Scholar] [CrossRef]
- Noh, N.J.; Son, Y.; Lee, S.K.; Yoon, T.K.; Seo, K.W.; Kim, C.; Lee, W.K.; Bae, S.W.; Hwang, J. Influence of stand density on soil CO2 efflux for a Pinus densiflora forest in Korea. J. Plant Res. 2010, 123, 411–419. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.; Koo, J.W.; Noh, N.J.; Kim, J.; Son, Y. Autotrophic and heterotrophic respiration in needle fir and Quercus-dominated stands in a cool-temperate forest, central Korea. J. Plant Res. 2010, 123, 485–495. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.; Jeong, J.; Bolan, N.S.; Naidu, R. Short-term effects of fertilizer application on soil respiration in red pine stands. J. Ecol. Field Biol. 2012, 35, 307–311. [Google Scholar] [CrossRef]
- Joo, J.H.; Lee, W.K.; Kim, T.H.; Lee, C.Y.; Jin, I.S.; Park, S.K.; Oh, M.Y. Studies on fertilization in pruning and thinning stands. The Res. Rep. For. Res. Insti., Korea 1983, 30, 155–189. [Google Scholar]
- Bond-Lamberty, B.; Bronson, D.; Bladyka, E.; Gower, S.T. A comparison of trenched plot techniques for partitioning soil respiration. Soil Biol. Biochem. 2011, 43, 2108–2114. [Google Scholar] [CrossRef]
- ArchMiller, A.A.; Samuelson, L.J. Partitioning longleaf pine soil respiration into its heterotrophic and autotrophic components through root exclusion. Forests 2016, 7, 39. [Google Scholar] [CrossRef]
- Statistical Analysis System (SAS) Institute Inc. SAS/STAT Statistical Software, version 9.1; SAS Publishing: Cary, NC, USA, 2003.
- Davidson, E.A.; Janssens, I.A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature 2006, 440, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Suseela, V.; Conant, R.T.; Wallenstein, M.D.; Dukes, J.S. Effects of soil moisture on the temperature sensitivity of heterotrophic respiration vary seasonally in an old-field climate change experiment. Glob. Chang. Biol. 2012, 18, 336–348. [Google Scholar] [CrossRef]
- Yuste, J.C.; Baldocchi, D.D.; Gershenson, A.; Goldstein, A.; Misson, L.; Wong, S. Microbial soil respiration and its dependency on carbon inputs, soil temperature and moisture. Glob. Chang. Biol. 2007, 13, 2018–2035. [Google Scholar]
- Kim, C.; Jeong, J. Comparison of soil CO2 efflux rates in Larix leptolepis, Pinus densiflora and P. rigitaeda plantations in Southern Korea. Dendrobiology 2016, 76, 51–60. [Google Scholar] [CrossRef]
- Davidson, E.A.; Belk, E.; Boone, R.D. Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest. Glob. Chang. Biol. 1998, 4, 217–227. [Google Scholar] [CrossRef]
- Bowden, R.D.; Davidson, E.; Savage, K.; Arabia, C.; Steudler, P. Chronic nitrogen additions reduce total soil respiration and microbial respiration in temperate forest soils at the Harvard Forest. For. Ecol. Manag. 2004, 196, 43–56. [Google Scholar] [CrossRef]
- Wei, W.; Weile, C.; Shaopeng, W. Forest soil respiration and its heterophic and autotrophic components: Global patterns and responses to temperate and precipitation. Soil Biol. Biochem. 2010, 42, 1236–1244. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Stand Density | DBH | Basal Area | Sand | Silt | Clay | C | N | P | K+ | Ca2+ | Mg2+ |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (trees·ha−1) | (cm) | (m2·ha−1) | (%) | (%) | (%) | (%) | (%) | (mg·kg−1) | (cmolc·kg−1) | |||
| Control | 1217 (133) a | 15.74 (0.84) a | 22.37 (1.95) a | 45 (3.5) a | 43 (3.0) a | 12 (1.0) a | 2.40 (0.28) a | 0.07 (0.01) a | 3.9 (0.40) a | 0.09 (0.01) a | 1.35 (0.19) b | 0.43 (0.05) b |
| N3P4K1 | 1150 (193) a | 15.89 (1.10) a | 20.56 (2.42) a | 42 (2.9) a | 44 (1.8) a | 14 (1.0) a | 2.82 (0.21) a | 0.09 (0.01) a | 6.5 (0.73) a | 0.09 (0.01) a | 1.77 (0.17) ab | 0.54 (0.04) ab |
| P4K1 | 1150 (152) a | 16.46 (1.46) a | 22.62 (2.00) a | 42 (2.0) a | 45 (1.9) a | 13 (1.8) a | 2.66 (0.27) a | 0.08 (0.01) a | 5.8 (1.61) a | 0.09 (0.01) a | 2.10 (0.26) a | 0.65 (0.05) a |
| Year | Treatment | df | Soil Respiration Rates (µmol·m−2·s−1) | Soil Temperature (°C) | ||
|---|---|---|---|---|---|---|
| Rh | Rs | Rh | Rs | |||
| 2011 | Control | - | 1.93 (0.20) | 3.08 (0.21) | 13.4 (0.10) | 13.7 (0.06) |
| N3P4K1 | - | 2.37 (0.26) | 3.05 (0.11) | 13.5 (0.04) | 13.7 (0.05) | |
| P4K1 | - | 2.20 (0.13) | 2.95 (0.12) | 13.4 (0.05) | 13.7 (0.03) | |
| Mean | - | 2.17 (0.12) | 3.02 (0.88) | 13.4 (0.04) | 13.7 (0.03) | |
| 2012 | Control | - | 1.91 (0.19) | 2.82 (0.20) | 12.8 (0.10) | 13.1(0.07) |
| N3P4K1 | - | 1.64 (3.19) | 3.19 (0.23) | 12.8 (0.10) | 13.2 (0.08) | |
| P4K1 | - | 1.77 (0.12) | 3.50 (0.16) | 12.8 (0.14) | 12.9 (0.15) | |
| Mean | - | 1.77 (0.09) | 3.02 (0.11) | 12.8 (0.06) | 13.1 (0.06) | |
| Mean | Control | - | 1.92 (0.14) | 2.95 (0.14) | 13.1 (0.07) | 13.4 (0.04) |
| N3P4K1 | - | 2.01 (0.16) | 3.12 (0.12) | 13.1 (0.03) | 13.4 (0.04) | |
| P4K1 | - | 1.98 (0.10) | 3.00 (0.10) | 13.1 (0.07) | 13.3 (0.07) | |
| p-value | Year (Y) | 1 | 0.009 | 0.973 | <0.001 | <0.001 |
| Treatment (F) | 2 | 0.882 | 0.598 | 0.733 | 0.504 | |
| Y × F | 2 | 0.152 | 0.447 | 0.806 | 0.253 | |
| Year | Treatment | Rh | Ra | Rs |
|---|---|---|---|---|
| (%) | ||||
| 2011 | Control | 63 | 37 | 100 |
| N3P4K1 | 78 | 22 | 100 | |
| P4K1 | 75 | 25 | 100 | |
| Mean | 72 | 28 | 100 | |
| 2012 | Control | 68 | 32 | 100 |
| N3P4K1 | 51 | 49 | 100 | |
| P4K1 | 58 | 42 | 100 | |
| Mean | 59 | 41 | 100 | |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, J.; Bolan, N.; Kim, C. Heterotrophic Soil Respiration Affected by Compound Fertilizer Types in Red Pine (Pinus densiflora S. et Z.) Stands of Korea. Forests 2016, 7, 309. https://doi.org/10.3390/f7120309
Jeong J, Bolan N, Kim C. Heterotrophic Soil Respiration Affected by Compound Fertilizer Types in Red Pine (Pinus densiflora S. et Z.) Stands of Korea. Forests. 2016; 7(12):309. https://doi.org/10.3390/f7120309
Chicago/Turabian StyleJeong, Jaeyeob, Nanthi Bolan, and Choonsig Kim. 2016. "Heterotrophic Soil Respiration Affected by Compound Fertilizer Types in Red Pine (Pinus densiflora S. et Z.) Stands of Korea" Forests 7, no. 12: 309. https://doi.org/10.3390/f7120309
APA StyleJeong, J., Bolan, N., & Kim, C. (2016). Heterotrophic Soil Respiration Affected by Compound Fertilizer Types in Red Pine (Pinus densiflora S. et Z.) Stands of Korea. Forests, 7(12), 309. https://doi.org/10.3390/f7120309