
The Biodiversity Benefits and Opportunity Costs of Plantation Forest Management: A Modelling Case Study of Pinus radiata in New Zealand

Abstract
:1. Introduction
2. Materials and Methods
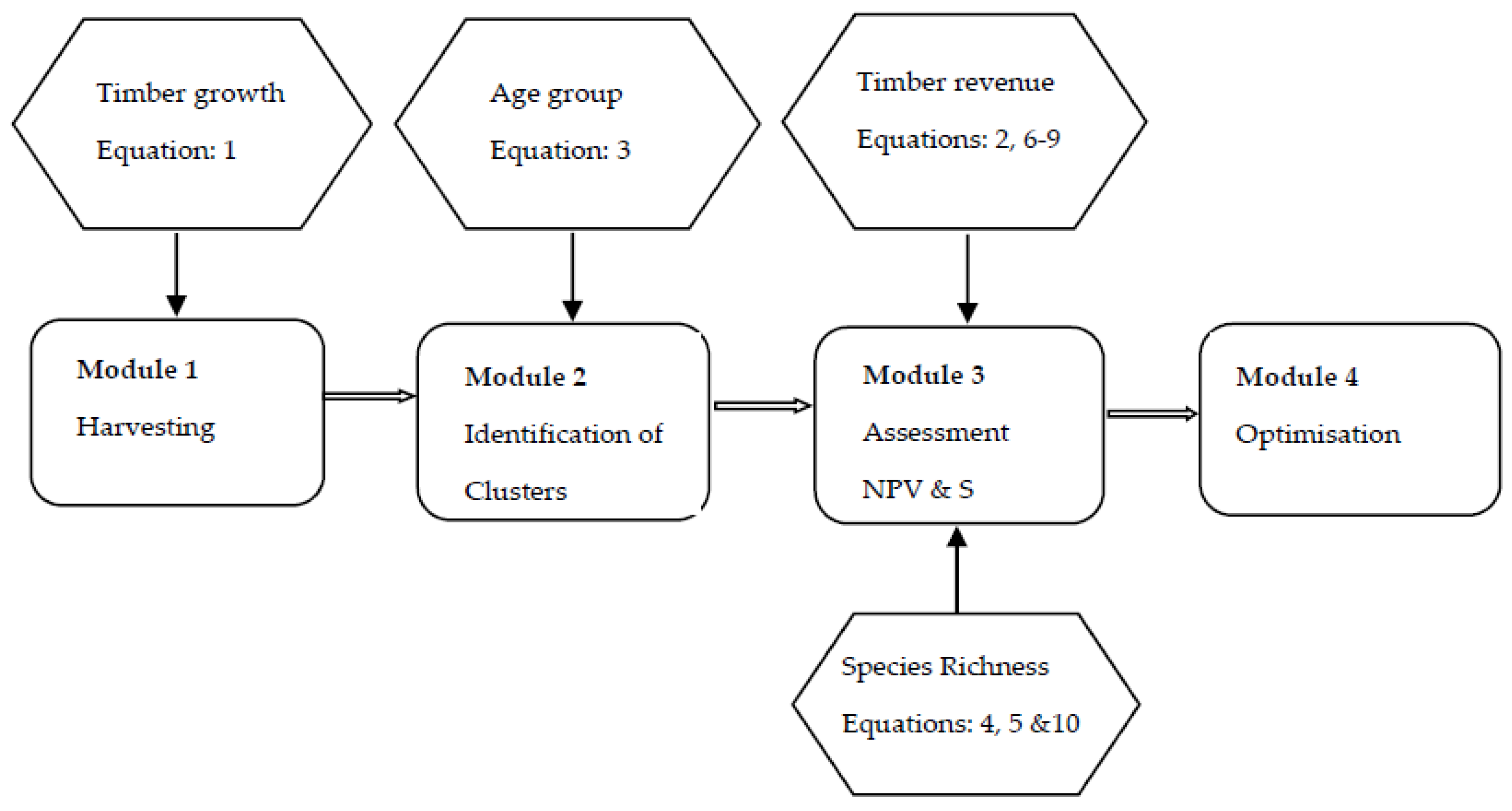
2.1. The Model
- Module 1: Involved calculating timber growth and a decision to cut or keep a stand in period t. The decisions to cut a stand vary with alternative management strategies. We let t run from 0 to T, and calculated timber volume based on Equations (1), (2), and (8).
- Module 2: Involved identifying clusters of clear-cutting area and of habitat area. We ran through stand 1 to stand n × n, ascertained whether (i.e., cut) or (i.e., habitat for plant species), and checked whether its adjacent stands are cut or aged older than A. If satisfied, clusters of clear cutting or habitat were recorded.
- Module 3: Involved calculating the NPV using Equations (6)–(9) and counting the number of species by applying Equations (5) and (10).
- Module 4: Involved all scenarios with different rotation ages being compared based on the objective variable. Then, the scenario that achieved the highest value of the NPV was chosen as the optimal management strategy in terms of timber extraction. The associated NPV and rotation length were referred to as NPV* and T*.
2.2. A Case Study of Pinus radiata in New Zealand
2.3. Sensitivity and Uncertainty Analyses
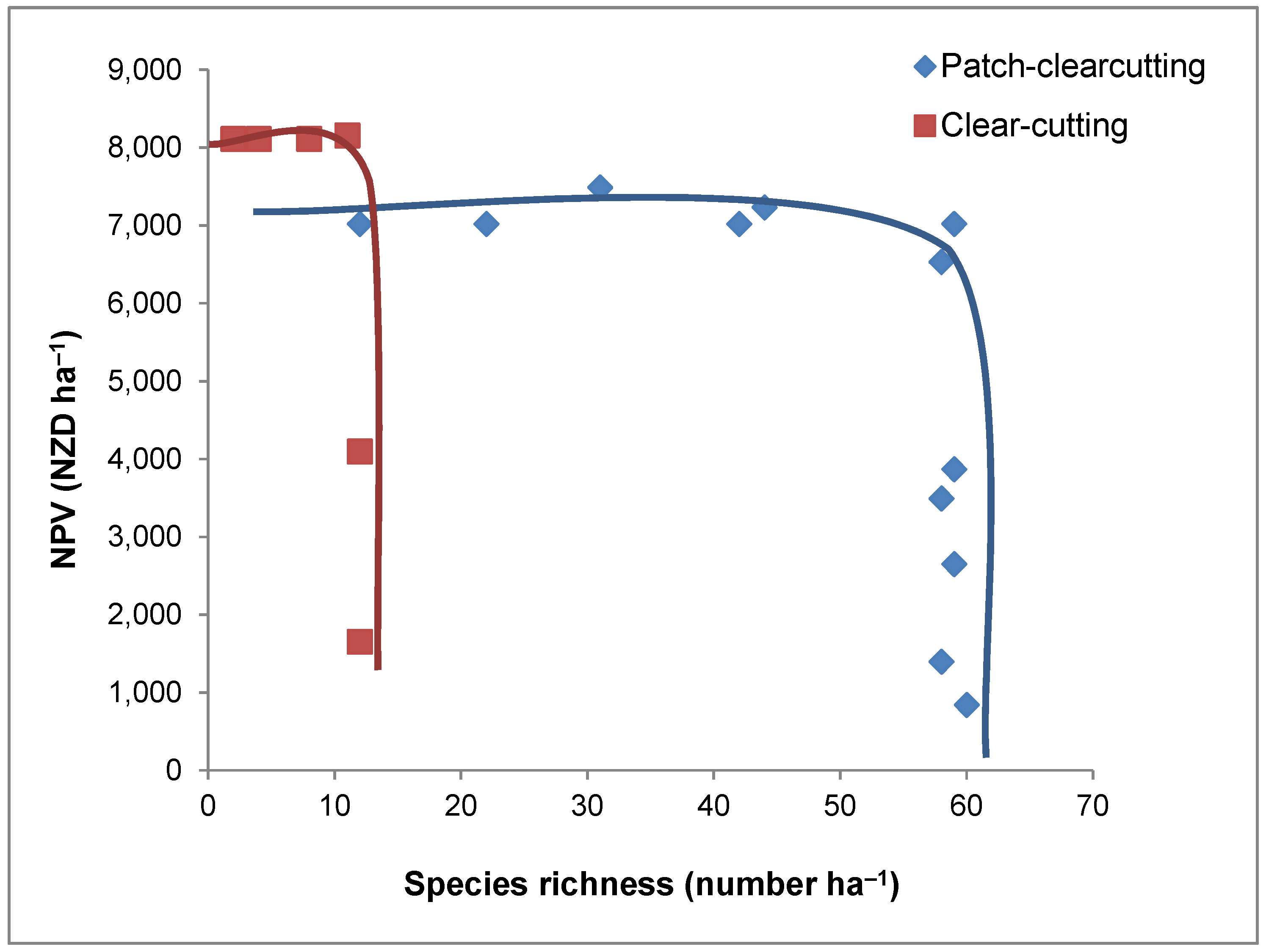
3. Results
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age (Years) | Timber Volume | a Timber Price | ||
|---|---|---|---|---|
| (m3·ha−1) | Standard Deviation (SD) (m3·ha−1) | (NZD·m−3) | SD (NZD·m−3) | |
| 1 | 0 | 0 | 77.78 | 15.56 |
| 2 | 0 | 0 | 77.78 | 15.56 |
| 3 | 0 | 0 | 77.78 | 15.56 |
| 4 | 0 | 0 | 77.78 | 15.56 |
| 5 | 2 | 1 | 77.78 | 15.56 |
| 6 | 5 | 3 | 77.78 | 15.56 |
| 7 | 12 | 7 | 77.78 | 15.56 |
| 8 | 24 | 11 | 77.78 | 15.56 |
| 9 | 39 | 17 | 77.78 | 15.56 |
| 10 | 59 | 23 | 77.78 | 15.56 |
| 11 | 82 | 30 | 77.78 | 15.56 |
| 12 | 109 | 36 | 81.46 | 16.29 |
| 13 | 137 | 43 | 83.42 | 16.68 |
| 14 | 168 | 49 | 85.70 | 17.14 |
| 15 | 201 | 54 | 87.88 | 17.58 |
| 16 | 234 | 60 | 89.91 | 17.98 |
| 17 | 268 | 65 | 91.26 | 18.25 |
| 18 | 302 | 69 | 92.48 | 18.50 |
| 19 | 337 | 74 | 93.62 | 18.72 |
| 20 | 371 | 74 | 94.14 | 18.83 |
| 21 | 403 | 76 | 94.58 | 18.92 |
| 22 | 436 | 81 | 94.95 | 18.99 |
| 23 | 469 | 82 | 95.17 | 19.03 |
| 24 | 501 | 85 | 96.93 | 19.39 |
| 25 | 527 | 88 | 99.79 | 19.96 |
| 26 | 558 | 90 | 100.36 | 20.07 |
| 27 | 587 | 93 | 100.80 | 20.16 |
| 28 | 614 | 95 | 101.13 | 20.23 |
| 29 | 640 | 97 | 101.59 | 20.32 |
| 30 | 667 | 100 | 101.81 | 20.36 |
| 31 | 693 | 101 | 102.15 | 20.43 |
| 32 | 719 | 102 | 102.21 | 20.44 |
| 33 | 745 | 104 | 102.41 | 20.48 |
| 34 | 769 | 104 | 102.60 | 20.52 |
| 35 | 793 | 105 | 102.91 | 20.58 |
| 36 | 817 | 104 | 103.10 | 20.62 |
| 37 | 840 | 103 | 103.33 | 20.67 |
| 38 | 863 | 103 | 103.45 | 20.69 |
| 39 | 888 | 101 | 103.57 | 20.71 |
| 40 | 913 | 100 | 103.65 | 20.73 |
Appendix B
| a Piece (log) Size (m3) | b Plantation Tree Size (m3) | a Harvesting Cost for a Piece (log) (NZD·m−3) | c Stand Size (ha) | c Harvesting Cost Per Stand for a Particular Stand Size on the Left Column (NZD·m−3) in 2011 |
|---|---|---|---|---|
| 0.8 | 1.1 | 10.0 | 1.0 | 40.78 |
| 1.3 | 1.3 | 8.0 | 2.0 | 32.65 |
| 2.0 | 2.5 | 7.2 | 5.0 | 29.37 |
| 3.0 | 6.0 | 6.6 | 10.0 | 26.89 |
| 4.5 | 15.1 | 6.0 | 30.0 | 25.42 |
| d 50.0 | d 24.51 |
Appendix C
| Scenarios | Patch-Clear-Cutting Strategy (P) | Clear-Cutting Strategy (C) | Differences (P − C) | OC ($) Per Extra Species | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| S* (Species·ha−1) | T* (Year) | NPV* ($·ha−1) | S* | T* | NPV* | ∆S* | ∆T* | ∆NPV* (OC $·ha−1) | ∆NPV*/∆S* | |
| Baseline | 59 | 25 | 7020 | 11 | 24 | 8157 | 48 | 1 | −1137 | 24 |
| Varying the stand size (ha)—a variable that can be modified by the foresters and planners | ||||||||||
| a = 5 | 31 | 25 | 7490 | 11 b | 24 b | 8157 b | 20 | 1 | −667 | 33 |
| a = 3 | 44 | 25 | 7234 | 11 b | 24 b | 8157 b | 34 | 1 | −923 | 28 |
| a = 1.5 | 72 | 25 | 6861 | 11 b | 24 b | 8157 b | 61 | 1 | −1295 | 21 |
| Varying the stand age that is assumed to create a suitable habitat for understorey plants (years) | ||||||||||
| A = 8 | 42 | 25 | 7020 | 8 | 24 | 8112 | 34 | 1 | −1093 | 33 |
| A = 16 | 22 | 25 | 7020 | 4 | 24 | 8112 | 18 | 1 | −1093 | 62 |
| A = 20 | 12 | 25 | 7020 | 2 | 24 | 8112 | 10 | 1 | −1093 | 111 |
| Varying the exponent of the harvesting cost | ||||||||||
| η = −0.2 | 59 | 25 | 7273 | 12 | 20 | 8913 | 47 | 5 | −1640 | 35 |
| η = −0.3 | 59 | 25 | 7564 | 12 | 20 | 9682 | 47 | 5 | −2118 | 45 |
| η = −0.4 | 59 | 25 | 7835 | 12 | 20 | 10,251 | 47 | 5 | −2417 | 51 |
| η = −0.5 | 59 | 25 | 8087 | 12 | 20 | 10,674 | 47 | 5 | −2586 | 55 |
| Sustainable management scenario with varied minimum harvesting ages (years) | ||||||||||
| T: 0–40 | 58 | 17 | 6531 | 11 b | 24 b | 8157 b | 47 | −7 c | −1626 | 34 |
| T: 25–40 | 59 | 25 | 3870 | 11 b | 24 b | 8157 b | 48 | 1 | −4287 | 89 |
| T: 28–40 | 59 | 28 | 2649 | 11 b | 24 b | 8157 b | 48 | 4 | −5507 | 114 |
| T: 35–40 | 60 | 35 | 839 | 11 b | 24 b | 8157 b | 49 | 11 | −7318 | 150 |
References
- Costanza, R.; D’Arge, R.; de Groot, R.; Farber, S.; Grasso, M.; Hannon, B.; Limburg, K.; Naeem, S.; O’Neill, R.V.; Paruelo, J.; et al. The value of the world’s ecosystem services and natural capital. Nature 1997, 387, 253–260. [Google Scholar] [CrossRef]
- Hassan, R.M.; Scholes, R.; Ash, N. Ecosystems and human well-being: Current state and trends. In The Millennium Ecosystem Assessment; Island Press: Washington, DC, USA, 2005; Volume 1, p. 23. [Google Scholar]
- Brockerhoff, E.G.; Ecroyd, C.E.; Leckie, A.C.; Kimberley, M.O. Diversity and succession of adventive and indigenous vascular understorey plants in Pinus radiata plantation forests in New Zealand. For. Ecol. Manag. 2003, 185, 307–326. [Google Scholar] [CrossRef]
- Laurance, W.F. Have we overstated the tropical biodiversity crisis? Trends Ecol. Evol. 2007, 22, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization of the United Nations (FAO). State of the World’s Forests 2007; Food and Agriculture Organization of the United Nations: Rome, Italy, 2007. [Google Scholar]
- Brockerhoff, E.G.; Jactel, H.; Parrotta, J.A.; Quine, C.P.; Sayer, J. Plantation forests and biodiversity: Oxymoron or opportunity? Biodivers. Conserv. 2008, 17, 925–951. [Google Scholar] [CrossRef]
- Barlow, J.; Gardner, T.A.; Araujo, I.S.; Ávila-Pires, T.C.; Bonaldo, A.B.; Costa, J.E.; Esposito, M.C.; Ferreira, L.V.; Hawes, J.; Hernandez, M.I.M.; et al. Quantifying the biodiversity value of tropical primary, secondary, and plantation forests. Proc. Natl. Acad. Sci. USA 2007, 104, 18555–18560. [Google Scholar] [CrossRef] [PubMed]
- Nyland, R.D. Silviculture: Concepts and Applications, 2nd ed.; McGraw-Hill Higher Education: New York, NY, USA, 2002; p. 704. [Google Scholar]
- Hagner, S. Forest Management in Temperate and Boreal Borests: Current Practices and the Scope for Implementing Sustainable Forest Management; Working Paper: Fao/fpirs/03; Food and Agriculture Organization of the United Nations (FAO), Forestry Policy and Planning Division: Rome, Italy, 1999; p. 47. [Google Scholar]
- Maclaren, J.P. Environmental Effects of Planted Forests in New Zealand: The Implications of Continued Afforestation of Pasture; New Zealand Forest Research Institute: Rotorua, New Zealand, 1996; Volume 198, p. 180. [Google Scholar]
- Applegate, G.; Putz, F.E.; Snook, L.K. Who Pays for and Who Benefits from Improved Timber Harvesting Practices in the Tropics? Lessons Learned and Information Gaps; Center for International Forestry Research: Jakarta, Indonesia, 2004; p. 35. [Google Scholar]
- Nasi, R.; Putz, F.E.; Pacheco, P.; Wunder, S.; Anta, S. Sustainable forest management and carbon in tropical latin america: The case for redd+. Forests 2011, 2, 200–217. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Hobbs, R.J. Fauna conservation in australian plantation forests—A review. Biol. Conserv. 2004, 119, 151–168. [Google Scholar] [CrossRef]
- Maynard, D.G.; MacIsaac, D.A. Soil nutrient and vegetation response to patch clear-cutting of an aspen forest near Meadow Lake, Saskatchewan. Can. J. Soil Sci. 1998, 78, 59–68. [Google Scholar] [CrossRef]
- Torras, O.; Saura, S. Effects of silvicultural treatments on forest biodiversity indicators in the Mediterranean. For. Ecol. Manag. 2008, 255, 3322–3330. [Google Scholar] [CrossRef]
- Pawson, S.; Brockerhoff, E.; Didham, R.; Norton, D. Clearfell harvest size: A key issue for biodiversity conservation in New Zealand’s plantation forests. N. Z. J. For. 2005, 50, 29–32. [Google Scholar]
- Nguyen, T.T.; Nghiem, N. Optimal forest rotation for carbon sequestration and biodiversity conservation by farm income levels. For. Policy Econ. 2016, 73, 185–194. [Google Scholar] [CrossRef]
- Calkin, D.E.; Montgomery, C.A.; Schumaker, N.H.; Polasky, S.; Arthur, J.L.; Nalle, D.J. Developing a production possibility set of wildlife species persistence and timber harvest value. Can. J. For. Res. 2002, 32, 1329–1342. [Google Scholar] [CrossRef]
- Hurme, E.; Kurttila, M.; Monkkonen, M.; Heinonen, T.; Pukkala, T. Maintenance of flying squirrel habitat and timber harvest: A site-specific spatial model in forest planning calculations. Landsc. Ecol. 2007, 22, 243–256. [Google Scholar] [CrossRef]
- Loomis, J.B.; White, D.S. Economic benefits of rare and endangered species: Summary and meta-analysis. Ecol. Econ. 1996, 18, 197–206. [Google Scholar] [CrossRef]
- Bann, C. An overview of valuation techniques: Advantages and limitations. Asean Biodivers. 2002, 2, 8–16. [Google Scholar]
- Martin-Lopez, B.; Montes, C.; Benayas, J. Economic valuation of biodiversity conservation: The meaning of numbers. Conserv. Biol. 2008, 22, 624–635. [Google Scholar] [CrossRef] [PubMed]
- Bandara, R.; Tisdell, C. Comparison of rural and urban attitudes to the conservation of Asian elephants in Sri Lanka: Empirical evidence. Biol. Conserv. 2003, 110, 327–342. [Google Scholar] [CrossRef]
- Eiswerth, M.E.; Van Kooten, G.C. The ghost of extinction: Preservation values and minimum viable population in wildlife models. Ecol. Econ. 2009, 68, 2129–2136. [Google Scholar] [CrossRef]
- Nghiem, T.H.N. Optimal Forest Management for Carbon Sequestration and Biodiversity Maintenance. Ph.D. Thesis, Massey University, Manawatu, New Zealand, 2011. [Google Scholar]
- Convention on Biological Diversity. Forest Biological Diversity. Available online: http://www.cbd.int/decisions/cop-06.shtml?m=COP-06&id=7196&lg=0 (accessed on 25 September 2007).
- Nghiem, N. Optimal forest management for timber value and carbon sequestration benefits in tropical planted forests: A case study of household foresters in Vietnam. Environ. Dev. Econ. 2015, 20, 746–766. [Google Scholar] [CrossRef]
- Nghiem, N. Optimal rotation age for carbon sequestration and biodiversity conservation in Vietnam. For. Policy Econ. 2014, 38, 56–64. [Google Scholar] [CrossRef]
- Nghiem, N. Biodiversity conservation attitudes and policy tools for promoting biodiversity in tropical planted forests. Biodivers. Conserv. 2013, 22, 373–403. [Google Scholar] [CrossRef]
- Woollons, R.; Whyte, A. Analysis of growth and yield from three kaingaroa thinning experiments. N. Z. For. 1989, 34, 12–15. [Google Scholar]
- Keenan, R.; Lamb, D.; Woldring, O.; Irvine, T.; Jensen, R. Restoration of plant biodiversity beneath tropical tree plantations in northern australia. For. Ecol. Manag. 1997, 99, 117–131. [Google Scholar] [CrossRef]
- Lugo, A.E. Comparison of tropical tree plantations with secondary forests of similar age. Ecol. Monogr. 1992, 62, 1–41. [Google Scholar] [CrossRef]
- Munro, N.T.; Fischer, J.; Wood, J.; Lindenmayer, D.B. Revegetation in agricultural areas: The development of structural complexity and floristic diversity. Ecol. Appl. 2009, 19, 1197–1210. [Google Scholar] [CrossRef] [PubMed]
- Bremer, L.L.; Farley, K.A. Does plantation forestry restore biodiversity or create green deserts? A synthesis of the effects of land-use transitions on plant species richness. Biodivers. Conserv. 2010, 19, 3893–3915. [Google Scholar] [CrossRef]
- Preston, F.W. The canonical distribution of commonness and rarity: Part I. Ecology 1962, 43, 185–215. [Google Scholar] [CrossRef]
- Roy, M.; Pascual, M.; Levin, S.A. Competitive coexistence in a dynamic landscape. Theor. Popul. Biol. 2004, 66, 341–353. [Google Scholar] [CrossRef] [PubMed]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Fonseca, G.A.B.D.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- The New Zealand Forest Owners Association. New Zealand Plantation Forestry. Available online: http://www.nzfoa.org.nz/index.php?/Home/New_Zealand_plantation_forestry (accessed on 29 January 2009).
- Ministry of Agriculture and Forestry. New Zealand Forest Industry Facts & Figures 2007/2008. Available online: http://www.maf.govt.nz/statistics/forestry/other-forestry-releases/facts-figures/ (accessed on 29 January 2009).
- Ministry of Agriculture and Forestry. A National Exotic Forest Description 2007. Available online: http://www.maf.govt.nz/mafnet/publications/nefd/national-exotic-forest-2007/index.htm (accessed on 29 January 2009).
- Food and Agriculture Organization of the United Nations (FAO). Global Forest Resources Assessment 2005; Food and Agriculture Organization of the United Nations: Rome, Italy, 2006. [Google Scholar]
- Ministry of Agriculture and Forestry. Future Drivers for New Zealand Forestry. Available online: http://www.maf.govt.nz/forestry/publications/future-drivers/ (accessed on 21 January 2009).
- Ausseil, A.G.E.; Dymond, J.R.; Kirschbaum, M.U.F.; Andrew, R.M.; Parfitt, R.L. Assessment of multiple ecosystem services in New Zealand at the catchment scale. Environ. Model. Softw. 2013, 43, 37–48. [Google Scholar] [CrossRef]
- Ministry for Primary Industries. National Exotic Forest Regional Yield Tables. Available online: http://www.mpi.govt.nz/news-resources/statistics-forecasting/statistical-publications/nefd-regional-yield-tables.aspx (accessed on 22 October 2012).
- Ministry of Agriculture and Forestry. Fact and Figure 2008/2009; Ministry of Agriculture and Forestry: Wellington, New Zealand, 2009.
- Murphy, G. Functions to predict average piece size and average haul volume for New Zealand clearfell cable logging operations. N. Z. J. For. 1984, 29, 60–65. [Google Scholar]
- Horgan, G. Financial Returns and Forestry Planting Rates; New Zealand Ministry of Agriculture and Forestry: Wellington, New Zealand, 2007.
- Ministry for Primary Industries. Indicative New Zealand Radiata Pine Log Prices. Available online: http://www.mpi.govt.nz/news-resources/statistics-forecasting/forestry/indicative-new-zealand-radiata-pine-log-prices (accessed on 15 July 2012).
- Dymond, J.R.; Ausseil, A.G.; Shepherd, J.D.; Janssen, H. A landscape approach for assessing the biodiversity value of indigenous forest remnants: Case study of the Manawatu/Wanganui region of New Zealand. Ecol. Econ. 2007, 64, 82–91. [Google Scholar] [CrossRef]
- Manley, B. Discount rates used for forest valuation—Results of 2007 survey. N. Z. J. For. 2007, 52, 21–27. [Google Scholar]
- Ogden, J.; Braggins, J.; Stretton, K.; Anderson, S. Plant species richness under Pinus radiata stands on the central North Island Volcanic Plateau, New Zealand. N. Z. J. Ecol. 1997, 21, 17–29. [Google Scholar]
- Ministry of Forestry. National Exotic Forest Description Regional Yield Tables as at 1 April 1995; Ministry of Forestry: Wellington, New Zealand, 1996.
- GAMS 23.6.5. Available online: http://www.gams.com/ (accessed on 5 November 2016).
- Heath, L.S.; Smith, J.E. An assessment of uncertainty in forest carbon budget projections. Environ. Sci. Policy 2000, 3, 73–82. [Google Scholar] [CrossRef]
- Eyvindson, K.; Kangas, A. Evaluating the required scenario set size for stochastic programming in forest management planning: Incorporating inventory and growth model uncertainty. Can. J. For. Res. 2015, 46, 340–347. [Google Scholar] [CrossRef]
- Berg, P. Radiata Pine—Growing and Harvesting the Forest, TE Ara—The Encyclopedia of New Zealand. Available online: http://www.TeAra.govt.nz/en/radiata-pine/page-4 (accessed on 22 October 2016).
- Rivas Palma, R.M. Environmental and Social Values from Plantation Forests: A Study in New Zealand with Focus on the Hawke’s Bay Region. Ph.D. Thesis, University of Canterbury, Christchurch, New Zealand, 2008. [Google Scholar]
- Rüger, N.; Gutiérrez, Á.; Kissling, W.D.; Armesto, J.J.; Huth, A. Ecological impacts of different harvesting scenarios for temperate evergreen rain forest in southern chile—A simulation experiment. For. Ecol. Manag. 2007, 252, 52–66. [Google Scholar] [CrossRef]
- Costanza, R.; Farber, S.C.; Maxwell, J. Valuation and management of wetland ecosystems. Ecol. Econ. 1989, 1, 335–361. [Google Scholar] [CrossRef]
- Fisher, A.C.; Krutilla, J.V. Resource conservation, environmental preservation, and the rate of discount. Q. J. Econ. 1975, 89, 358–370. [Google Scholar] [CrossRef]
- Howarth, R.B.; Norgaard, R.B. Intergenerational transfers and the social discount rate. Environ. Resour. Econ. 1993, 3, 337–358. [Google Scholar] [CrossRef]
- Rambaud, S.C.; Torrecillas, M.J.M. Some considerations on the social discount rate. Environ. Sci. Policy 2005, 8, 343–355. [Google Scholar] [CrossRef]
- Weitzman, M.L. On the “environmental” discount rate. J. Environ. Econ. Manag. 1994, 26, 200–209. [Google Scholar] [CrossRef]
- Millennium Ecosystem Assessment. Ecosystems and Human Well-Being: General Synthesis; Island Press: Washington, DC, USA, 2005. [Google Scholar]
- Milne, S.; Niesten, E. Direct payments for biodiversity conservation in developing countries: Practical insights for design and implementation. Oryx 2009, 43, 530–541. [Google Scholar] [CrossRef]
- Turpie, J.K.; Marais, C.; Blignaut, J.N. The working for water programme: Evolution of a payments for ecosystem services mechanism that addresses both poverty and ecosystem service delivery in South Africa. Ecol. Econ. 2008, 65, 788–798. [Google Scholar] [CrossRef]
- Madsen, B.; Carroll, N.; Brands, K.M. State of Biodiversity Markets: Offset and Compensation Programs Worldwide; Ecosystem Marketplace: Washington, DC, USA, 2010. [Google Scholar]
- Kinzig, A.; Perrings, C.; Chapin, F., III; Polasky, S.; Smith, V.; Tilman, D.; Turner, B., II. Paying for ecosystem services—Promise and peril. Science 2011, 334, 603–604. [Google Scholar] [CrossRef] [PubMed]
- Wellington Regional Council. Hutt River Floodplain Management Plan—For the Hutt River and Its Environment; Wellington Regional Council: Wellington, New Zealand, 2001.
- Busch, J.; Godoy, F.; Turner, W.R.; Harvey, C.A. Biodiversity co-benefits of reducing emissions from deforestation under alternative reference levels and levels of finance. Conserv. Lett. 2011, 4, 101–115. [Google Scholar] [CrossRef]
- Carnus, J.M.; Parrotta, J.; Brockerhoff, E.G.; Arbez, M.; Jactel, H.; Kremer, A.; Lamb, D.; O’Hara, K.; Walters, B. Planted forests and biodiversity. In IUFRO Occasional Paper; IUFRO: Vienna, Austria, 2003; pp. 31–50. [Google Scholar]
- Evans, J. The multiple roles of planted forests. In Planted Forests. Uses, Impacts and Sustainability; CAB International and FAO: Wallingford, UK, 2009; pp. 61–90. [Google Scholar]
- Seaton, R. The Ecological Requirements of the New Zealand Falcon (Falco novaeseelandiae) in Plantation Forestry. Ph.D. Thesis, Massey University, Palmerston North, New Zealand, 2007. [Google Scholar]
- Hanski, I. Metapopulation dynamics. Nature 1998, 396, 41–49. [Google Scholar] [CrossRef]
- Lindenmayer, D.; Franklin, J.F. Conserving Forest Biodiversity: A Comprehensive Multiscaled Approach, 2nd ed.; Island Press: Washington, DC, USA, 2002; p. 351. [Google Scholar]
- Allen, R.B.; Platt, K.H.; Coker, R.E.J. Understorey species composition patterns in a Pinus radiata plantation on the central north island volcanic plateau, New Zealand. N. Z. J. For. Sci. 1995, 25, 301–317. [Google Scholar]
- Terlesk, C.J. Harvesting: Pinus radiata production and cost trends. N. Z. J. For. 1980, 25, 172–183. [Google Scholar]
- Long, A. Environmentally Sound Forest Harvesting; School of Forest Resources and Conservation, Florida Cooperative Extension Service, Institute of Food and Agricultural Sciences, University of Florida Gainesville: Gainesville, FL, USA, 1994; Volume SS-FOR-6, p. 11. [Google Scholar]
- Ministry of Forestry. Small forest management. In Harvesting a Small Forest; Ministry of Forestry and New Zealand Logging Industry Research Organisation: Wellington, New Zealand, 1996; p. 39. [Google Scholar]
- Dickson, A.; Bloomberg, M. An economic feasibility study of partial harvesting in a radiata pine plantation in canterbury, New Zealand. In Proceedings of the Conference Joint Australia and New Zealand Institute of Forestry, Queenstown, New Zealand, 27 April–1 May 2003; Mason, E., Perley, C., Eds.; Ministry of Agriculture and Forestry: Wellington, New Zealand, 2003; pp. 370–378. [Google Scholar]
- Russell, F.; Mortimer, D. A Review of Small-Scale Harvesting Systems in Use Worldwide and Their Potential Application in Irish Forestry; COFORD: Dublin, Ireland, 2005; p. 48. [Google Scholar]
- Zhang, Y.; Zhang, D.; Schelhas, J. Small-scale non-industrial private forest ownership in the United States: Rationale and implications for forest management. Silva Fenn. 2005, 39, 443–454. [Google Scholar] [CrossRef]
- OECD. Key Short-Term Economic Indicators Metadata: Consumer Prices—Annual Inflation. Available online: http://stats.oecd.org/index.aspx?queryid=21761 (accessed on 15 June 2011).
- Kantola, M. Role of Forestry in Small Farms. Unasylva 1967, 21. Available online: http://www.Fao.Org/docrep/59708e/59708e03.Htm (accessed on 5 November 2016). [Google Scholar]




| Parameters | Unit | Values | Sensitivity Analyses | Uncertainty Analyses |
|---|---|---|---|---|
| Forest harvest size (a) | ha | 2 | 1.5–5 | No |
| Total stand (n) | number | 400 | No | No |
| Time horizon (T) | year | 100 | No | No |
| Yield (q) | m3 | See Appendix A | No | Normal distribution |
| Exponent of the species area curve (z) | numeric | 0.3 | 0.15–0.32 | No |
| Biodiversity age (A) | year | 0 | 8–20 | No |
| Cost exponent (η) | numeric | −0.118 (see Appendix B) | (−0.5)–(−0.2) | No |
| Price (p) | NZD | See Appendix A | No | Normal distribution |
| Discount rate (r) | rate | 0.07 | 0–0.15 | No |
| Scenarios | Patch-Clear-Cutting Strategy (P) | Clear-Cutting Strategy (C) | Differences (P–C) | OC ($) Per Extra Species | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| S* (Species·ha−1) | T* (Year) | NPV* ($·ha−1) | S* | T* | NPV* | ∆S* | ∆T* | ∆NPV* (OC $·ha−1) | ∆NPV*/∆S* | |
| Baseline | 59 | 25 | 7020 | 11 | 24 | 8157 | 48 | 1 | −1137 | 24 |
| Varying the discount rate | ||||||||||
| r = 0 | 60 | 33 | 136,954 | 12 | 33 | 157,473 | 48 | 0 | −20,519 | 429 |
| r = 0.03 | 59 | 26 | 26,894 | 12 | 26 | 31,064 | 47 | 0 | −4170 | 88 |
| r = 0.1 | 58 | 19 | 3493 | 12 | 19 | 4098 | 47 | 0 | −605 | 13 |
| r = 0.15 | 58 | 17 | 1396 | 12 | 16 | 1651 | 46 | 1 | −255 | 5 |
| Scenarios | Patch-Clear-Cutting (P) | Clear-Cutting (C) | Differences (P − C) | OC ($) Per Extra Species ∆NPV*/∆S* | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| S* (Species·ha−1) | T* (Year) | NPV* ($·ha−1) | S* | T* | NPV* | ∆S* | ∆T* | ∆NPV* (OC $·ha−1) | ||
| Mean | 58.59 | 22.5 | 10,103 | 11.68 | 22.1 | 11,362 | 46.90 | 0.4 | −1258 | 27 |
| Median | 58.48 | 23.0 | 9985 | 11.67 | 22.0 | 11,239 | 46.81 | 1.0 | −1257 | 27 |
| Lower 95% SI | 57.25 | 15.0 | 7875 | 11.42 | 15.0 | 8940 | 45.83 | 0.0 | −1066 | 23 |
| Upper 95% SI | 59.4 | 32.0 | 13,276 | 11.79 | 31.0 | 14,747 | 47.61 | 1.0 | −1471 | 31 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nghiem, N.; Tran, H. The Biodiversity Benefits and Opportunity Costs of Plantation Forest Management: A Modelling Case Study of Pinus radiata in New Zealand. Forests 2016, 7, 297. https://doi.org/10.3390/f7120297
Nghiem N, Tran H. The Biodiversity Benefits and Opportunity Costs of Plantation Forest Management: A Modelling Case Study of Pinus radiata in New Zealand. Forests. 2016; 7(12):297. https://doi.org/10.3390/f7120297
Chicago/Turabian StyleNghiem, Nhung, and Hop Tran. 2016. "The Biodiversity Benefits and Opportunity Costs of Plantation Forest Management: A Modelling Case Study of Pinus radiata in New Zealand" Forests 7, no. 12: 297. https://doi.org/10.3390/f7120297
APA StyleNghiem, N., & Tran, H. (2016). The Biodiversity Benefits and Opportunity Costs of Plantation Forest Management: A Modelling Case Study of Pinus radiata in New Zealand. Forests, 7(12), 297. https://doi.org/10.3390/f7120297