
Forest Restoration Using Variable Density Thinning: Lessons from Douglas-Fir Stands in Western Oregon

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
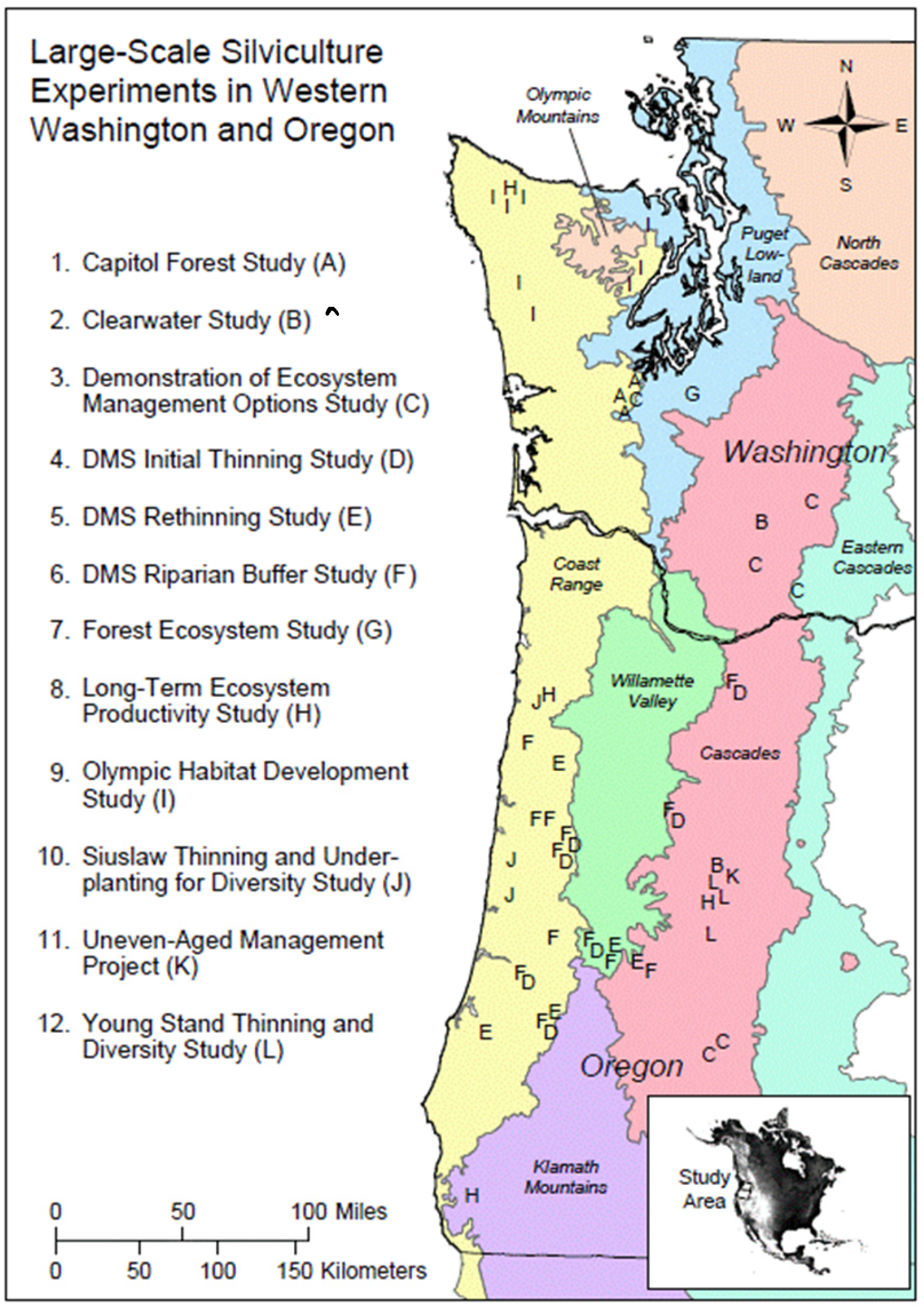
2. Materials and Methods
3. Results and Discussion
3.1. Lessons Learned about Restoration Treatments and Large-Scale Experiments
3.1.1. Financial and Operational Feasibility
3.1.2. Variability and Multiple Scales
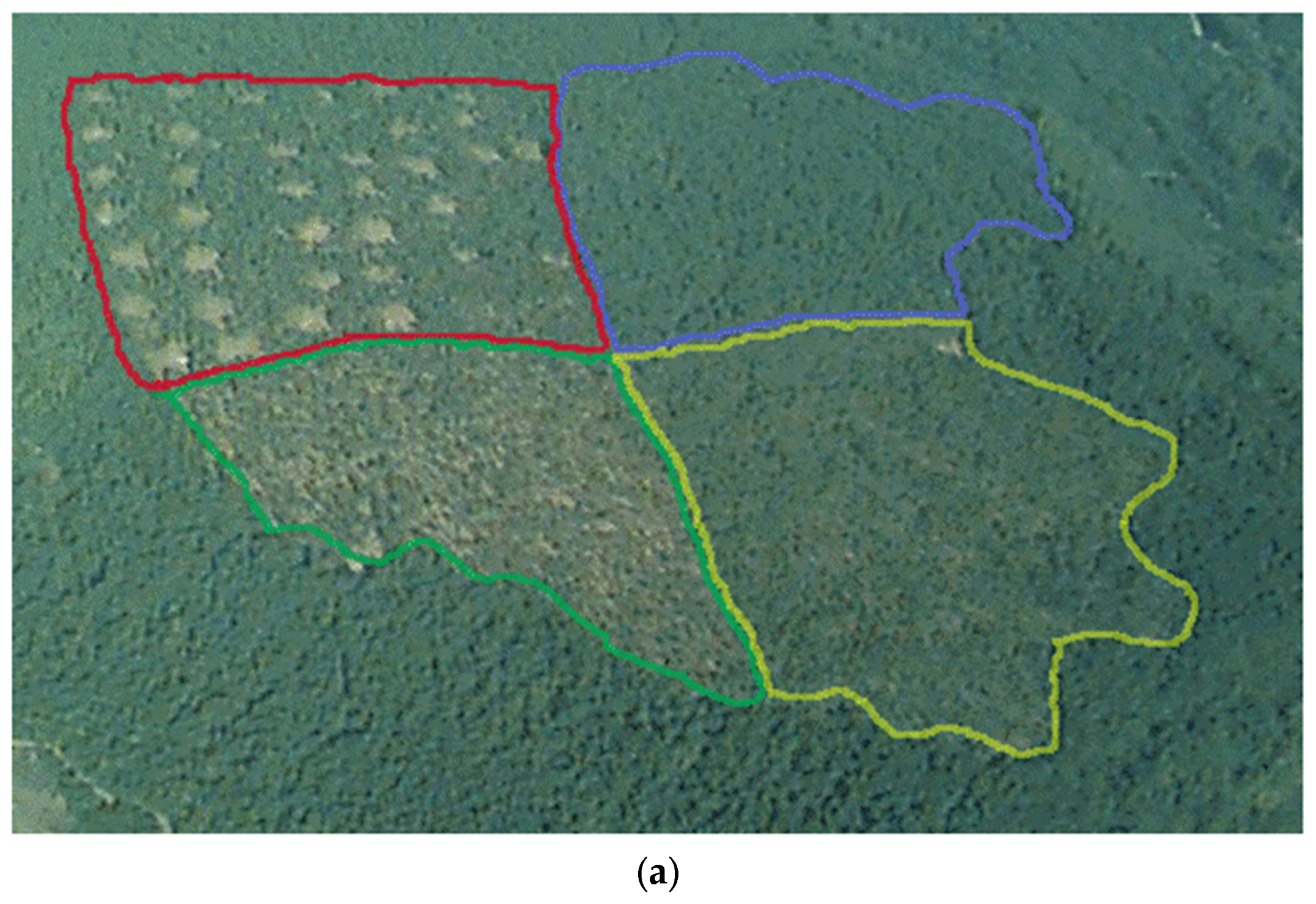
3.1.3. Treatment Choices and Definitions
3.1.4. Treatment Implementation
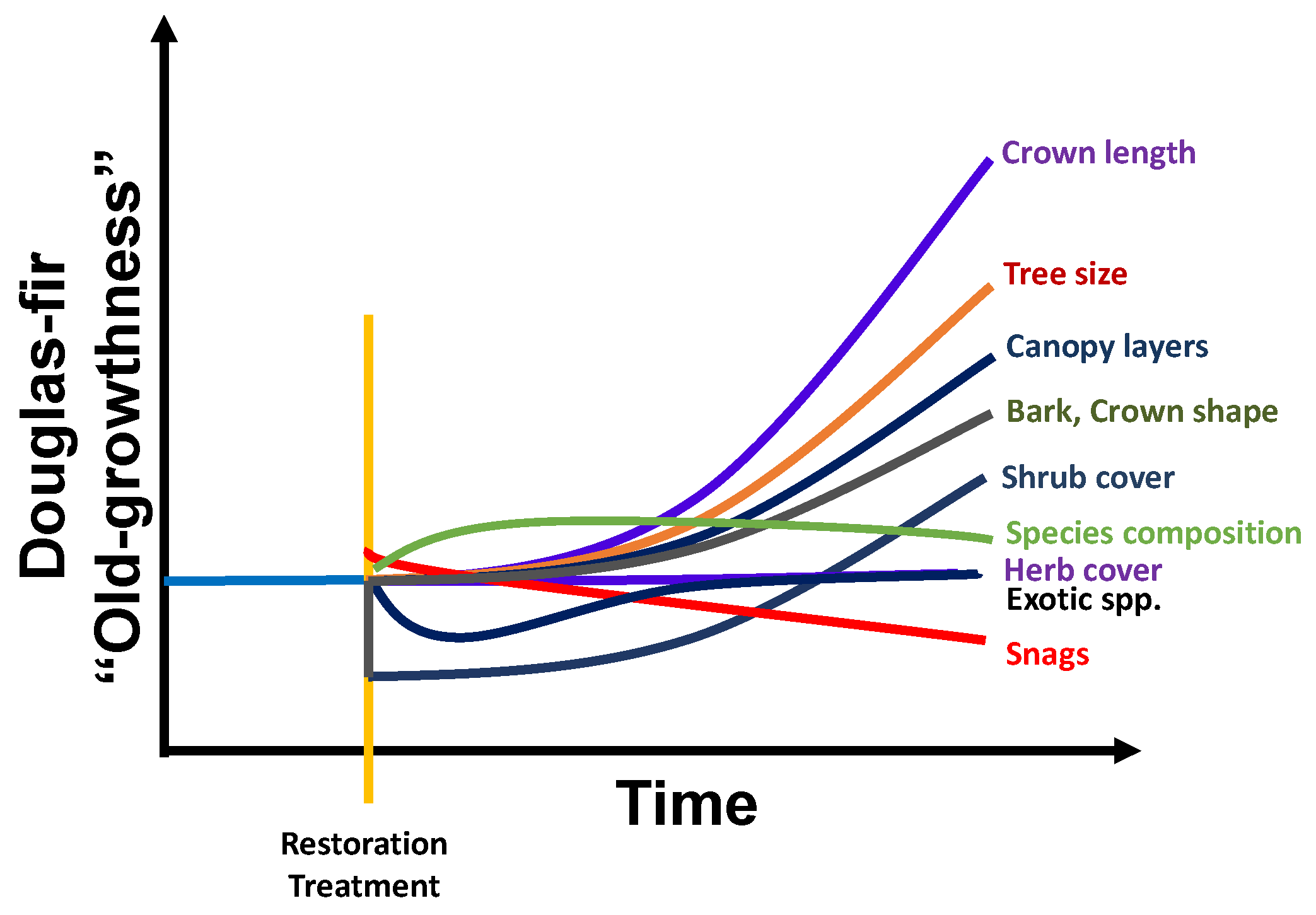
3.2. Vegetation Responses
3.2.1. Tree and Stand Growth Responses
3.2.2. Tree Mortality
3.2.3. Canopy and Crown Size and Structure
3.2.4. Bark Characteristics
3.2.5. Tree Regeneration (Natural)
3.2.6. Understory Vegetation
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Waring, R.H.; Franklin, J.F. Evergreen coniferous forests of the Pacific Northwest. Science 1979, 204, 1380–1386. [Google Scholar] [CrossRef] [PubMed]
- Agee, J.K. Fire Ecology of Pacific Northwest Forests; Island Press: Washington, DC, USA, 1993. [Google Scholar]
- Schwantes, C.A. The Pacific Northwest: An Interpretive History; University of Nebraska Press: Lincoln, NE, USA, 1996. [Google Scholar]
- Arnold, D.; Williams, G.W.; Dombeck, M. The US Forest Service in the Pacific Northwest: A History. Public Hist. 2011, 33, 115–116. [Google Scholar] [CrossRef]
- Forsman, E.D. A Preliminary Investigation of the Spotted Owl in Oregon. Master’s Thesis, Oregon State University, Corvallis, OR, USA, 1976. [Google Scholar]
- U.S. Department of Agriculture; U.S. Department of Interior. Record of Decision for Amendment to Forest Service and Bureau of Land Management Planning Documents within the Range of the Northern Spotted Owl (Northwest Forest Plan); U.S. Department of Interior, Bureau of Land Management: Portland, OR, USA, 1994.
- Monserud, R.A. Large-scale management experiments in the moist maritime forests of the Pacific Northwest. Landsc. Urban Plan. 2002, 59, 159–180. [Google Scholar] [CrossRef]
- Poage, N.J.; Anderson, P.D. Large-Scale Silviculture Experiments of Western Oregon and Washington; General Technical Report PNW-GTR-713; USDA Forest Service: Portland, OR, USA, 2007; pp. 1–44.
- Anderson, P.D.; Ronnenberg, K.L. (Eds.) Density Management in the 21st Century: West Side Story; General Technical Report PNW-GTR-880; U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 2013; pp. 1–249.
- Omernik, J.M. Ecoregions of the contiguous United States. Ann. Assoc. Am. Geogr. 1987, 77, 118–125. [Google Scholar] [CrossRef]
- Manning, T.; Friesen, C. The Young Stand Thinning & Diversity Study (YSTDS): Establishment Report, Study Plan, and Key Findings. 2013. Available online: http://ecoshare.info/projects/central-cascade-adaptive-management-partnership/forest-studies/young-stand-thinning-and-diversity-study/ (accessed on 4 November 2016).
- Cissel, J.H.; Anderson, P.D.; Olson, D.; Puettmann, K.J.; Berryman, S.; Chan, S.; Thompson, C. BLM Density Management and Riparian Buffer Study: Establishment Report and Study Plan; Scientific Investigations Report 2006-5087; U.S. Geological Survey: Washington, DC, USA, 2006; pp. 1–151.
- Oliver, C.D.; Larson, B.C. Forest Stand Dynamics; John Wiley and Sons: New York, NY, USA, 1996; p. 520. [Google Scholar]
- Puettmann, K.J.; Dodson, E.K.; Ares, A.; Berger, C.A. Short-term responses of overstory and understory vegetation to thinning treatments: A tale of two studies. In Density Management in the 21st Century: West Side Story; General Technical Report PNW-GTR-880; U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 2013; pp. 1–249. [Google Scholar]
- Dodson, E.K.; Ares, A.; Puettmann, K.J. Early responses to thinning treatments designed to accelerate late successional forest structure in young coniferous stands of western Oregon, USA. Can. J. For. Res. 2012, 42, 345–355. [Google Scholar] [CrossRef]
- Kuehne, C.; Puettmann, K.J. Natural regeneration in thinned Douglas-fir stands in western Oregon. J. Sustain. For. 2008, 27, 246–274. [Google Scholar] [CrossRef]
- Neill, A.; Puettmann, K.J. Managing for adaptive capacity: Thinning improves food availability for wildlife and insect pollinators under climate change conditions. Can. J. For. Res. 2013, 43, 428–440. [Google Scholar] [CrossRef]
- Dodson, E.K.; Burton, J.I.; Puettmann, K.J. Multiscale controls on natural regeneration dynamics after partial overstory removal in Douglas-fir Forests in western Oregon, USA. For. Sci. 2014, 60, 953–961. [Google Scholar] [CrossRef]
- Burton, J.I.; Ganio, L.M.; Puettmann, K.J. Multi-scale spatial controls of understory vegetation in Douglas-fir–western hemlock forests of western Oregon, USA. Ecosphere 2014, 5, 1–34. [Google Scholar] [CrossRef]
- Puettmann, K.J.; Tappeiner, J.C. Multi-scale assessments highlight silvicultural opportunities to increase species diversity and spatial variability in forests. Forestry 2014, 87, 1–10. [Google Scholar] [CrossRef]
- Hayes, J.; Weikel, J.; Huso, M. Response of birds to thinning young Douglas-fir forests. Ecol. Appl. 2003, 13, 1222–1232. [Google Scholar] [CrossRef]
- Yegorova, S.; Betts, M.G.; Hagar, J.; Puettmann, K.J. Bird-vegetation associations in thinned and unthinned young Douglas-fir forests 10 years after thinning. For. Ecol. Manag. 2013, 310, 1057–1070. [Google Scholar] [CrossRef]
- Ruzicka, K.J.; Puettmann, K.J.; Olson, D.H. Management of Riparian Buffers: Upslope Thinning with Downslope Impacts. For. Sci. 2014, 60, 881–892. [Google Scholar] [CrossRef]
- Cole, E.; Newton, M. Tenth-year survival and size of underplanted seedlings in the Oregon Coast Range. Can. J. For. Res. 2009, 39, 580–595. [Google Scholar] [CrossRef]
- Kellogg, L.D. A comparison of logging planning, felling, and skyline yarding costs between clearcutting and five group-selection harvesting methods. West. J. Appl. For. 1996, 11, 90–96. [Google Scholar]
- Lutz, J.A.; Halpern, C.B. Tree mortality during early forest development: A long-term study of rates, causes, and consequences. Ecol. Monogr. 2006, 76, 257–275. [Google Scholar] [CrossRef]
- Aussenac, G.; Granier, A. Effects of thinning on water stress and growth in Douglas-fir. Can. J. For. Res. 1988, 18, 100–105. [Google Scholar] [CrossRef]
- Brooks, J.R.; Mitchell, A.K. Interpreting tree responses to thinning and fertilization using tree-ring stable isotopes. New Phytol. 2011, 190, 770–782. [Google Scholar] [CrossRef] [PubMed]
- Davis, L.R.; Puettmann, K.J.; Tucker, G.F. Overstory response to alternative thinning treatments in young Douglas-fir forests of western Oregon. Northwest Sci. 2007, 81, 1–14. [Google Scholar] [CrossRef]
- Roberts, S.D.; Harrington, C.A. Individual tree growth response to variable-density thinning in coastal Pacific Northwest forests. For. Ecol. Manag. 2008, 255, 2771–2781. [Google Scholar] [CrossRef]
- Franklin, J.F.; Spies, T.A. Ecological Definitions of Old-Growth Douglas-Fir Forests; USDA Forest Service General Technical Report PNW-GTR-285; Pacific Northwest Research Station: Portland, OR, USA, 1991.
- Marshall, D.D.; Curtis, R.O. Levels-of-Growing-Stock Cooperative Study in Douglas-Fir: Report No. 15—Hoskins: 1963–1998; Research Paper PNW-RP-337; Pacific Northwest Research Station: Portland, OR, USA, 2002.
- Newton, M.; Cole, L. Overstory development in Douglas-fir-dominant forests thinned to enhance late-seral features. For. Sci. 2015, 61, 809–816. [Google Scholar] [CrossRef]
- Fairweather, S.E. Recommendation of growth model and tree volume estimation system to the H&H core team for the Tillamook, Forest Grove, Astoria, West Oregon, and Western Lane districts. In Oregon Department of Forestry Harvest Scheduling Model Yield Table Creation Project; Mason, Bruce and Girard, Inc.: Portland, OR, USA, 2004; pp. 1–23. [Google Scholar]
- Burton, J.I.; Ares, A.; Mulford, S.E.; Olson, D.H.; Puettmann, K.J. Above-ground carbon storage, downed wood, and understory plant species richness after thinning in western Oregon. In Density Management in the 21st Century: West Side Story; Anderson, P.D., Ronnenberg, K.L., Eds.; General Technical Report PNW-GTR-880; U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 2013; pp. 1–249. [Google Scholar]
- Lewis, J.C. Creating snags and wildlife trees in commercial forest landscapes. West. J. Appl. For. 1998, 13, 97–101. [Google Scholar]
- Aussenac, G. Interactions between forest stands and microclimate: Ecophysiological aspects and consequences for silviculture. Ann. For. Sci. 2000, 57, 287–301. [Google Scholar] [CrossRef]
- Swanson, M.E.; Franklin, J.F.; Beschta, R.L.; Crisafulli, C.M.; DellaSala, D.A.; Hutto, R.L.; Lindenmayer, D.B.; Swanson, F.J. The forgotten stage of forest succession: Early-successional ecosystems on forest sites. Front. Ecol. Environ. 2011, 9, 117–125. [Google Scholar] [CrossRef]
- Brandeis, T.J.; Newton, M.; Cole, E. A comparison of overstory density measures for describing understory conifer growth. For. Ecol. Manag. 2001, 152, 149–157. [Google Scholar] [CrossRef]
- Chan, S.S.; Larson, D.J.; Maas-Hebner, K.G.; Emmingham, W.H.; Johnston, S.R.; Mikowski, D.A. Overstory and understory development in thinned and underplanted Oregon Coast Range Douglas-fir stands. Can. J. For. Res. 2006, 36, 2696–2711. [Google Scholar] [CrossRef]
- Wilson, J.S.; Oliver, C.D. Stability and density management in Douglas-fir plantations. Can. J. For. Res. 2000, 30, 910–920. [Google Scholar] [CrossRef]
- Weiskittel, A.R.; Maguire, D.A.; Monserud, R.A. Modeling crown structural responses to competing vegetation control, thinning, fertilization, and Swiss needle cast in coastal Douglas-fir of the Pacific Northwest, USA. For. Ecol. Manag. 2007, 245, 96–109. [Google Scholar] [CrossRef]
- Maguire, D.A.; Kershaw, J.A., Jr.; Hann, D.W. Predicting the effects of silvicutlural regime on branch size and crown wood core in Douglas-fir. For. Sci. 1991, 37, 1409–1428. [Google Scholar]
- Seidel, D.; Ruzicka, K.J.; Puettmann, K. Canopy gaps affect the shape of Douglas-fir crowns in the western Cascades, Oregon. For. Ecol. Manag. 2016, 363, 31–38. [Google Scholar] [CrossRef]
- Michel, A.K.; Winter, S. Tree microhabitat structures as indicators of biodiversity in Douglas-fir forests of different stand ages and management histories in the Pacific Northwest, USA. For. Ecol. Manag. 2009, 257, 1453–1464. [Google Scholar] [CrossRef]
- Michel, A.K.; Winter, S.; Linde, A. The effect of tree dimension on the diversity of bark microhabitat structures and bark use in Douglas-fir (Pseudotsuga menziesii var. menziesii). Can. J. For. Res. 2011, 41, 300–308. [Google Scholar] [CrossRef]
- Sheridan, C.D.; Puettmann, K.J.; Huso, M.M.P.; Hagar, J.C.; Falk, K.R. Management, morphological, and environmental factors influencing Douglas-fir bark furrows in the Oregon Coast Range. West. J. Appl. For. 2013, 28, 97–106. [Google Scholar] [CrossRef]
- Franklin, J.F.; Van Pelt, R. Spatial aspects of structural complexity in old-growth forests. J. For. 2004, 102, 22–28. [Google Scholar]
- Spies, T.A. Ecological concepts and diversity of old-growth forests. J. For. 2004, 102, 14–20. [Google Scholar]
- Spies, T.A.; Franklin, J.F. Old growth and forest dynamics in the douglas-fir region of western Oregon and Washington. Nat. Areas J. 1988, 8, 190–201. [Google Scholar]
- Urgenson, L.S.; Halpern, C.B.; Anderson, P.D. Twelve-year responses of planted and naturally regenerating conifers to variable-retention harvest in the Pacific Northwest, USA. Can. J. For. Res. 2012, 43, 46–55. [Google Scholar] [CrossRef]
- Newton, M.; Cole, E.C. Harvesting impacts on understory regeneration in two-storied Douglas-fir stands. West. J. Appl. For. 2006, 21, 14–18. [Google Scholar]
- Shatford, J.P.A.; Bailey, J.D.; Tappeiner, J.C. Understory tree development with repeated stand density treatments in coastal Douglas-fir forests of Oregon. West. J. Appl. For. 2009, 24, 11–16. [Google Scholar]
- Donato, D.C.; Campbell, J.L.; Franklin, J.F. Multiple successional pathways and precocity in forest development: Can some forests be born complex? J. Veg. Sci. 2012, 23, 576–584. [Google Scholar] [CrossRef]
- Puettmann, K.J.; Berger, C.A. Development of tree and understory vegetation in young Douglas-fir plantations in western Oregon. West. J. Appl. For. 2006, 21, 94–101. [Google Scholar]
- Ares, A.; Neill, A.R.; Puettmann, K.J. Understory abundance, species diversity and functional attribute response to thinning in coniferous stands. For. Ecol. Manag. 2010, 260, 1104–1113. [Google Scholar] [CrossRef]
- Fahey, R.T.; Puettmann, K.J. Patterns in spatial extent of gap influence on understory plant communities. For. Ecol. Manag. 2008, 255, 2801–2810. [Google Scholar] [CrossRef]
- Fahey, R.T.; Puettmann, K.J. Ground-layer disturbance and initial conditions influence gap partitioning of understory vegetation. J. Ecol. 2007, 95, 1098–1109. [Google Scholar] [CrossRef]
- Ares, A.; Berryman, S.D.; Puettmann, K.J. Understory vegetation response to thinning disturbance of varying complexity in coniferous stands. Appl. Veg. Sci. 2009, 12, 472–487. [Google Scholar] [CrossRef]
- Davis, L.R.; Puettmann, K.J. Initial response of understory vegetation to three alternative thinning treatments. J. Sustain. For. 2009, 28, 904–934. [Google Scholar] [CrossRef]
- Hagar, J.C. Wildlife species associated with non-coniferous vegetation in Pacific Northwest conifer forests: A review. For. Ecol. Manag. 2007, 246, 108–122. [Google Scholar] [CrossRef]
- Wilson, D.S.; Anderson, P.D.; Puettmann, K.J. Evaluating the consistency of understorey vegetation response to forest thinning through synthetic analysis of operational-scale experiments. Forestry 2009, 82, 583–596. [Google Scholar] [CrossRef]
- Brady, T.J.; Monleon, V.J.; Gray, A.N. Calibrating vascular plant abundance for detecting future climate change in Oregon and Washington, USA. Ecol. Indic. 2010, 10, 657–667. [Google Scholar] [CrossRef]
- Burton, J.I.; Ares, A.; Olson, D.H.; Puettmann, K.J. Management trade-off between aboveground carbon storage and understory plant species richness in temperate forests. Ecol. Appl. 2013, 23, 1297–1310. [Google Scholar] [CrossRef] [PubMed]





© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puettmann, K.J.; Ares, A.; Burton, J.I.; Dodson, E.K. Forest Restoration Using Variable Density Thinning: Lessons from Douglas-Fir Stands in Western Oregon. Forests 2016, 7, 310. https://doi.org/10.3390/f7120310
Puettmann KJ, Ares A, Burton JI, Dodson EK. Forest Restoration Using Variable Density Thinning: Lessons from Douglas-Fir Stands in Western Oregon. Forests. 2016; 7(12):310. https://doi.org/10.3390/f7120310
Chicago/Turabian StylePuettmann, Klaus J., Adrian Ares, Julia I. Burton, and Erich Kyle Dodson. 2016. "Forest Restoration Using Variable Density Thinning: Lessons from Douglas-Fir Stands in Western Oregon" Forests 7, no. 12: 310. https://doi.org/10.3390/f7120310
APA StylePuettmann, K. J., Ares, A., Burton, J. I., & Dodson, E. K. (2016). Forest Restoration Using Variable Density Thinning: Lessons from Douglas-Fir Stands in Western Oregon. Forests, 7(12), 310. https://doi.org/10.3390/f7120310