
Influence of Mature Overstory Trees on Adjacent 12-Year Regeneration and the Woody Understory: Aggregated Retention versus Intact Forest

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
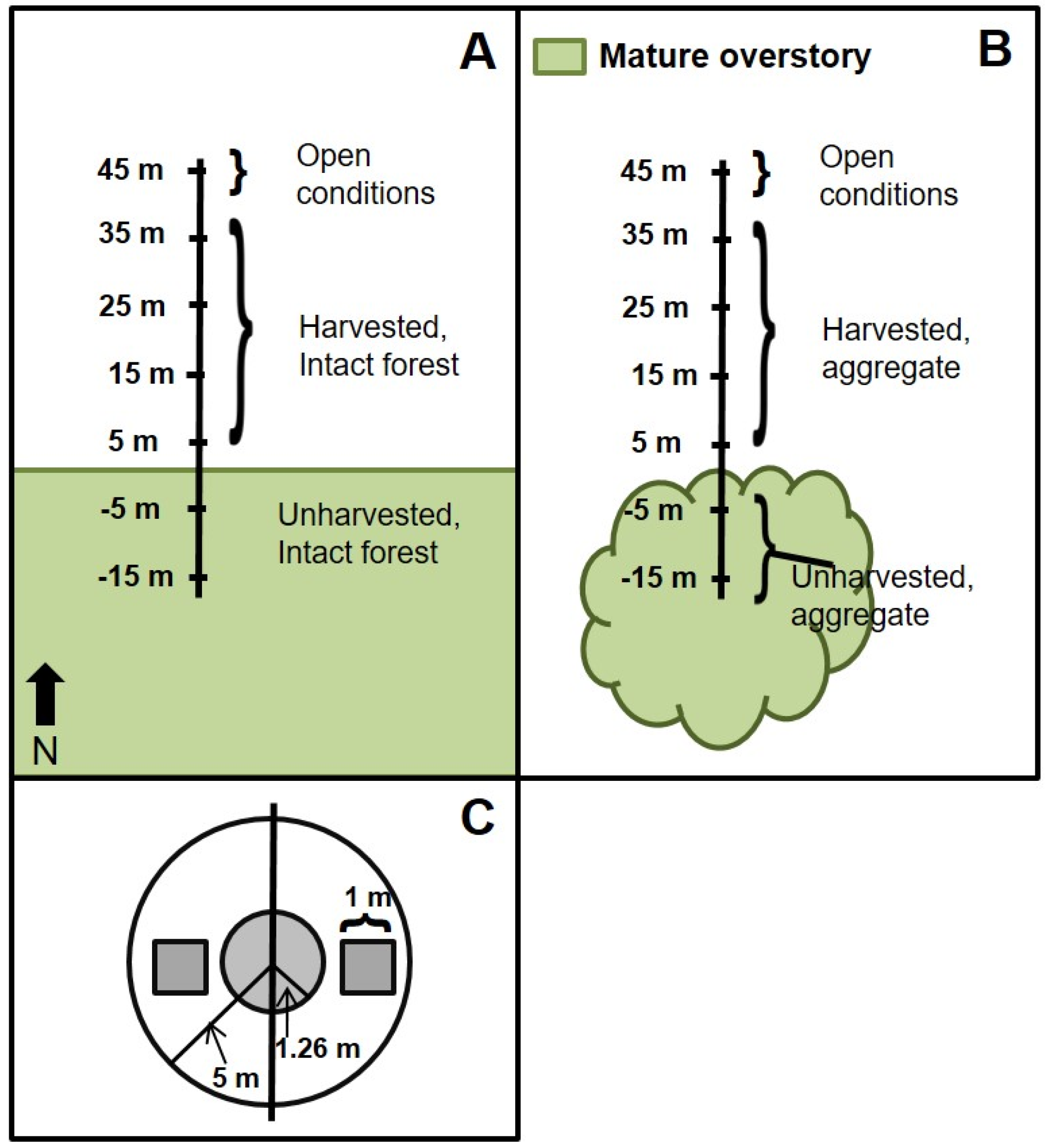
2.1. Study Area
2.2. Field Methods
2.3. Analysis
2.3.1. Understory Forest Structure and Standing Biomass
2.3.2. Forest Influence and Edge Effects
3. Results
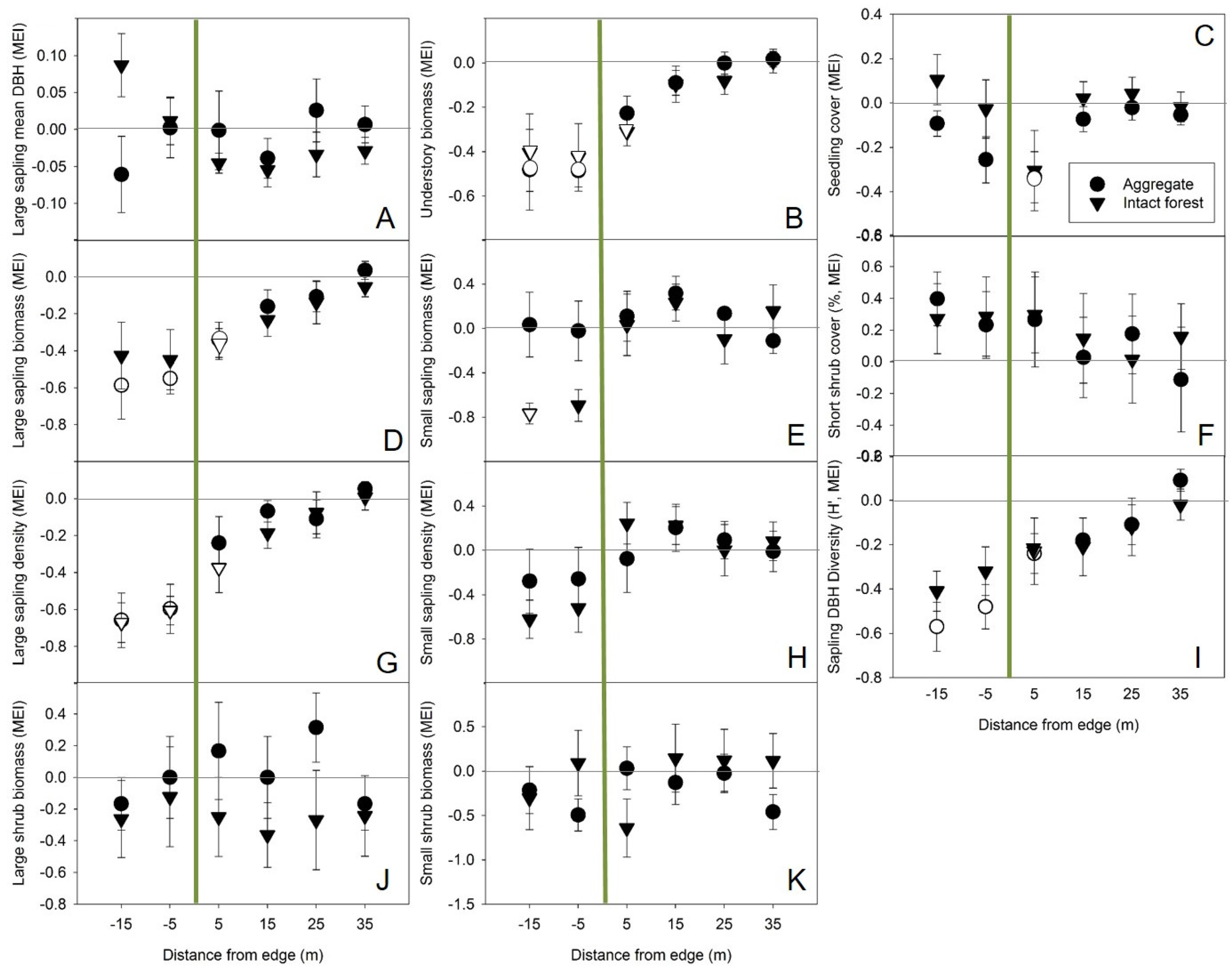
3.1. Forest Structure and Standing Biomass
3.2. Magnitude and Distance of Forest Influence and Edge Effects
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
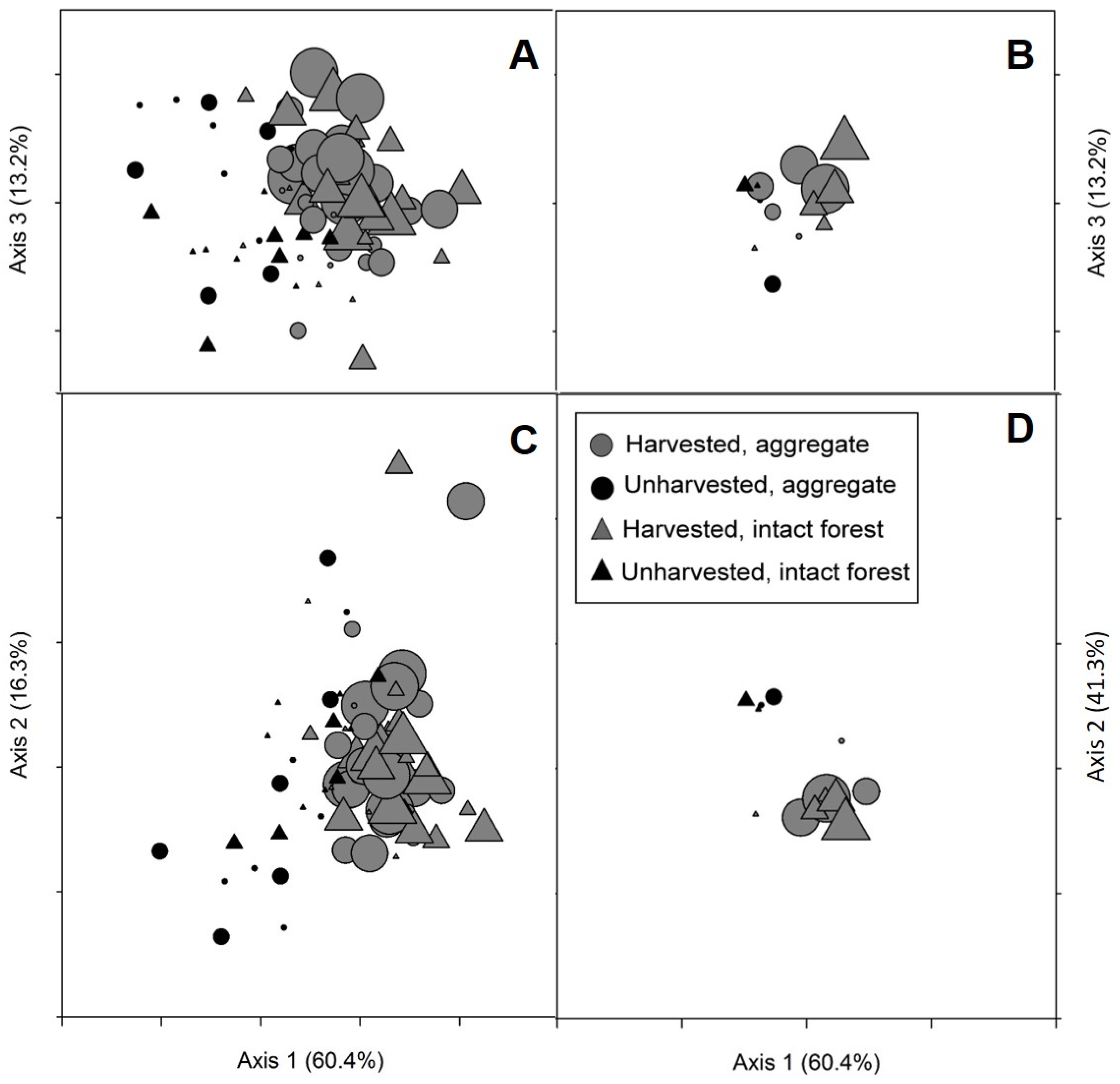
Appendix A. Effects of Overstory Condition and Distance from Harvest Edge on Plot Distribution in Species Space (NMS)

{kind=link}
{kind=link}
{kind=link}
| Effect | Df | F | p-Value |
|---|---|---|---|
| Overstory | 1 | 1.332 | 0.246 |
| Location | 5 | 15.105 | 0.0002 |
| Overstory × Location | 5 | 0.7216 | 0.615 |
| Δ Axis 1 (60.4%) | Δ Axis 2 (16.3%) | Δ Axis 3 (13.2%) | |||||
|---|---|---|---|---|---|---|---|
| Source | DF | F | p-Value | F | p-Value | F | p-Value |
| Overstory condition | 1 | 1.77 | 0.1902 | 2.35 | 0.1317 | 4.13 | 0.0477 |
| Location | 5 | 2.09 | 0.0835 | 11.48 | <0.0001 | 2.12 | 0.0787 |
| Condition × Location | 5 | 0.65 | 0.6626 | 0.78 | 0.5686 | 2.24 | 0.0658 |
| Location (m) | Δ Axis 1 | Δ Axis 2 | Δ Axis 3 |
|---|---|---|---|
| −15 | −0.61 (0.267) | 0.75 (0.109) a | −0.53 (0.119) |
| −5 | −0.61 (0.232) | 0.69 (0.107) a,b | −0.20 (0.125) |
| 5 | −0.21 (0.202) | 0.23 (0.182) b,c | −0.45 (0.081) |
| 15 | 0.09 (0.144) | −0.07 (0.128) c | −0.40 (0.184) |
| 25 | 0.04 (0.230) | 0.07 (0.124) c | −0.23 (0.152) |
| 35 | −0.14 (0.176) | −0.13 (0.090) c | −0.09 (0.128) |
| Overstory Condition | Δ Axis 1 | Δ Axis 2 | Δ Axis 3 |
|---|---|---|---|
| Aggregate | −0.12 (0.111) | 0.31 (0.096) | −0.19 (0.069) a |
| Intact forest | −0.44 (0.150) | 0.15 (0.088) | −0.44 (0.088) b |
Appendix B. Multilevel Models for Structure Response to Distance from Harvest Edge
| Response | Fixed Effects | Random Effects | ||||||
|---|---|---|---|---|---|---|---|---|
| Intercept | p | Distance | p | Aggregate (Intercept) | Intact Forest (Intercept) | p | R2 | |
| Understory woody BM (Mg/ha) | 12.17 (1.38) | 0.0003 | 0.19 (0.04) | <0.0001 | 0.20 (0.51) | −0.20 (0.51) | 0.760 | 0.44 |
| Large sapling BM (Mg/ha) | 7.75 (1.74) | 0.0343 | 0.21 (0.04) | <0.0001 | 0.88 (1.26) | −0.88 (1.26) | 0.613 | 0.58 |
| Small sapling BM (Mg/ha0.5) | 1.42 (0.28) | 0.0497 | −0.01 (0.05) | 0.12 | −0.03 (0.11) | 0.03 (0.11) | 0.827 | 0.70 |
| Large sapling density (stems/ha) | 5104.04 (698.33) | 0.0041 | 86.26 (18.13) | <0.0001 | 16.99 (131.65) | −6.99 (131.65) | 0.918 | 0.59 |
| Small sapling density(Mg/ha0.5) | 110.89 (32.47) | 0.0574 | −0.25 (0.39) | 0.53 | −2.89 (8.54) | 2.89 (8.54) | 0.792 | 0.71 |
| Diversity (H’), sapling DBH | 0.24 (0.04) | 0.0087 | 0.005 (0.001) | <0.0001 | 0.02 (0.03) | −0.02 (0.03) | 0.577 | 0.53 |
| POTR, large sapling BM (Mg/ha) | 4.41 (2.36) | 0.1885 | 0.29 (0.061) | <0.0001 | 0.924 (1.367) | −0.924 (1.367) | 0.622 | 0.46 |
References
- Seidl, R.; Fernandes, P.M.; Fonseca, T.F.; Gillet, F.; Jönsson, A.M.; Merganičová, K.; Netherer, S.; Arpaci, A.; Bontemps, J.D.; Bugmann, H.; et al. Modeling natural disturnaces in forest ecosytems: A review. Ecol. Model. 2011, 222, 903–924. [Google Scholar] [CrossRef]
- Dale, V.H.; Joyce, L.A.; McNulty, S.; Neilson, R.P.; Ayres, M.P.; Flannigan, M.D.; Hanson, P.J.; Irland, L.C.; Lugo, A.E.; Peterson, C.J.; et al. Climate change and forest disturbances. Bioscience 2001, 51, 723–734. [Google Scholar] [CrossRef]
- Lindner, M.; Maroschek, M.; Netherer, S.; Kremer, A.; Barbati, A.; Garcia-Gonzalo, J.; Seidl, R.; Delzon, S.; Coronoa, P.; Kolstrom, M.; et al. Climate impacts, adaptive capacity, and vulnerability of European forest ecosystems. For. Ecol. Manag. 2010, 259, 698–709. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Franklin, J.F.; Lohmus, A.; Baker, S.C.; Bauhus, J.; Beese, W.; Brodie, A.; Kiehl, B.; Kouki, J.; Martinez Pastur, G.; et al. A major shift to the retention approach for forestry can help resolve some global forest sustainability issues. Conserv. Lett. 2012, 5, 421–431. [Google Scholar] [CrossRef]
- Fedrowitz, K.; Koricheva, J.; Baker, S.C.; Lindenmayer, D.B.; Palik, B.J.; Rosenvald, R.; Beese, W.; Franklin, J.F.; Kouki, J.; Macdonald, S.E.; et al. Can retention forestry help conserve biodiversity? A meta-analysis. J. Appl. Ecol. 2014, 51, 1669–1679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gustafsson, L.; Kouki, J.; Sverdrup-Thygeson, A. Tree retention as a conservation measure in clear-cut forests of northern Europe: A review of ecological consequences. Scand. J. For. Res. 2010, 25, 295–308. [Google Scholar] [CrossRef]
- Seymour, R.; Hunter, M.L., Jr. Principles of ecological forestry. In Composition in Managing Forests for Biodiversity; Hunter, M.L., Jr., Ed.; Cambridge University Press: New York, NY, USA, 2002; pp. 22–61. [Google Scholar]
- Franklin, J.F.; Spies, T.A.; Van Pelt, R.; Carey, A.B.; Thornburgh, D.A.; Berg, D.R.; Lindenmayer, D.B.; Harmon, M.E.; Keeton, W.S.; Shaw, D.C. Disturbances and structural development of natural forest ecosystems with silvicultural implications, using Douglas-fir forests as an example. For. Ecol. Manag. 2002, 155, 399–423. [Google Scholar] [CrossRef]
- Hunter, M.L., Jr. Maintaining biodiversity in forest ecosystems. In Composition in Managing Forests for Biodiversity; Hunter, M.L., Jr., Ed.; Cambridge University Press: New York, NY, USA, 2002; pp. 3–21. [Google Scholar]
- Halpern, C.B.; Halaj, J.; Evans, S.A.; Doviak, M. Level and pattern of overstory retention interact to shape long-term responses of understories to timber harvest. Ecol. Appl. 2012, 22, 2049–2064. [Google Scholar] [CrossRef] [PubMed]
- McElhinny, C. Forest and Woodland Structure as an Index of Biodiversity: A Review; A literature review; NSW National Parks and Wildlife Service: Sydney, Australia, 2002; p. 84.
- Rosenvald, R.; Lõhmus, A. For what, when, and where is green-tree retention better than clear-cutting? A review of the biodiversity aspects. For. Ecol. Manag. 2008, 255, 1–15. [Google Scholar] [CrossRef]
- Heithecker, T.D.; Halpern, C.B. Edge-related gradients in microclimate in forest aggregates following structural retention harvests in western Washington. For. Ecol. Manag. 2007, 248, 163–173. [Google Scholar] [CrossRef]
- Baker, T.P.; Jordan, G.J.; Steel, E.A.; Fountain-Jones, M.M.; Wardlaw, T.J.; Baker, S.C. Microlimate through space and time: Microclimatic variation at the edge of regeneration forests over daily, yearly and decadal time scales. For. Ecol. Manag. 2014, 334, 174–184. [Google Scholar] [CrossRef]
- Baker, S.C.; Halpern, C.B.; Wardlaw, T.J.; Crawford, R.L.; Bigley, R.E.; Edgar, G.J.; Evans, S.A.; Franklin, J.F.; Jordan, G.J.; Karpievitch, Y.; et al. Short- and long-term benefits for forest biodiversity of retaining unlogged patches in harvested areas. For. Ecol. Manag. 2015, 353, 187–195. [Google Scholar] [CrossRef]
- Hansen, A.J.; Garman, S.L.; Weigand, J.F.; Urban, D.L.; McComb, W.C.; Raphael, M.G. Alternative silvicultural regeimes in Pacific Northwest: Simulations of ecological and economic effects. Ecol. Appl. 1995, 5, 535–554. [Google Scholar] [CrossRef]
- Halpern, C.B.; Evans, S.A.; Nelson, C.R.; McKenzie, D.; Ligouri, D.A. Response of forest vegetation to varying levels and patterns of green-tree retention: An overview of a long-term experiment. Northwest Sci. 1999, 73, 27–44. [Google Scholar]
- Franklin, J.F.; Berg, D.R.; Thornburgh, D.A.; Tappeiner, J.C. Alternative silvicultural approaches to timber harvesting: Variable retention harvest systems. In Creating a Forestry for the 21st Century; Kohm, K.A., Franklin, J.F., Eds.; Island Press: Washington, DC, USA, 1997; pp. 111–139. [Google Scholar]
- Gustafsson, L.; Baker, S.C.; Bauhus, J.; Beese, W.J.; Brodie, A.; Kouki, J.; Lindenmayer, D.B.; Lõhmus, A.; Guillermo, M.P.; Messier, C.; et al. Retention forestry to maintain multifunctional forests: A world perspective. Bioscience 2012, 62, 633–645. [Google Scholar]
- Palik, B.J.; Engstrom, R.T. Species composition. In Managing for Biodiversity; Hunter, M.L., Jr., Ed.; Cambridge University Press: New York, NY, USA, 2002; pp. 65–94. [Google Scholar]
- Wright, E.F.; Coates, K.D.; Canham, C.D.; Bartemucci, P. Species variability in growth response to light across climatic regions in northwestern British Columbia. Can. J. For. Res. 1998, 28, 871–886. [Google Scholar] [CrossRef]
- Newsome, T.A.; Henieman, J.L.; Nemec, A.F.L.; Comeau, P.G.; Arsenault, A.; Waterhouse, M. Ten-year regeneration responses to varying levels of overstory retention in two productive southern British Columbia ecosystems. For. Ecol. Manag. 2010, 260, 132–145. [Google Scholar] [CrossRef]
- Boyden, S.; Montgomery, R.; Reich, P.B.; Palik, B. Seeing the forest for the heterogeneous trees: Stand-scale resource distributions emerge from tree-scale structure. Ecol. Appl. 2012, 22, 1578–1588. [Google Scholar] [CrossRef] [PubMed]
- Palik, B.J.; Mitchell, R.J.; Pecot, S.; Battaglia, M.; Mou, P. Spatial distribution of overstory retention influences resources and growth of longleaf pine seedlings. Ecol. Appl. 2003, 13, 674–686. [Google Scholar] [CrossRef]
- Aubry, K.B.; Halpern, C.B.; Maguire, D.A. Ecological effects of variable-retention harvests in the north-western United States: The DEMO study. For. Snow Landsc. Res. 2004, 78, 119–137. [Google Scholar]
- Mitchell, A.K.; Koppenall, R.; Goodmanson, G.; Benton, R.; Brown, T. Regenerating montane conifers with variable retention systems in a coastal British Columbia forest: 10-year results. For. Ecol. Manag. 2007, 246, 240–250. [Google Scholar] [CrossRef]
- Urgenson, L.S.; Halpern, C.B.; Anderson, P.D. Twelve-year responses of planted and natural regenerating conifers to variable-retention harvest in the Pacific Northwest, USA. Can. J. For. Res. 2013, 43, 46–55. [Google Scholar] [CrossRef]
- Mitchell, S.J.; Beese, W.J. The retention system: Reconciling variable retention with the principles of silvicultural systems. For. Chron. 2002, 78, 397–403. [Google Scholar] [CrossRef]
- Frey, B.R.; Lieffers, V.J.; Landhausser, S.M.; Comeau, P.G.; Greenway, K.J. An analysis of sucker regeneration of trembling aspen. Can. J. For. Res. 2003, 33, 1169–1179. [Google Scholar] [CrossRef]
- Luoma, D.L.; Stockdale, C.A.; Molina, R.; Eberhart, J.L. The spatial influence of Pseudotsuga menziesii retention trees on ectomycorrhizal diversity. Can. J. For. Res. 2006, 36, 2561–2573. [Google Scholar] [CrossRef]
- Bose, A.K. Sapling recruitment and mortality dynamics following partial harvesting in aspen-dominated mixedwoods in eastern Canada. For. Ecol. Manag. 2004, 329, 37–48. [Google Scholar] [CrossRef]
- Royo, A.A.; Carson, W.P. On the formation of dense understory layers in forests worldwide: Consequences and implcations for forest dynamics, biodiversity, and succession. Can. J. For. Res. 2006, 36, 1345–1362. [Google Scholar] [CrossRef]
- Dovčiak, M.; Reich, P.B.; Frelich, L.E. Seed rain, safe sites, competing vegetation and soil resources spatially structure white pine regeneration and recruitment. Can. J. For. Res. 2003, 33, 1892–1904. [Google Scholar] [CrossRef]
- Montgomery, R.A.; Palik, B.J.; Boyden, S.B.; Reich, P.B. New cohort growth and survival in variable retention harvests of a pine ecosystem in Minnesota, USA. For. Ecol. Manag. 2013, 310, 327–335. [Google Scholar] [CrossRef]
- MacDonald, G.B.; Cherry, M.L.; Thompson, D.J. Effect of harvest intensity on development of natural regeneration and shrubs in an Ontario boreal mixedwood stand. For. Ecol. Manag. 2004, 189, 207–222. [Google Scholar] [CrossRef]
- Pre´vost, M.; Pothier, D. Partial cuts in a trembling aspen conifer stand: Effects on microenvironmental conditions and regeneration dynamics. Can. J. For. Res. 2003, 33, 1–15. [Google Scholar] [CrossRef]
- Brais, S.; Harvey, B.D.; Bergeron, Y.; Messier, C.; Greene, D.; Belleau, A.; Pare, D. Testing forest ecosystem management in boreal mixedwoods of northwestern Quebec: Initial response of aspen stands to different levels of harvesting. Can. J. For. Res. 2004, 34, 431–446. [Google Scholar] [CrossRef]
- Palik, B.; Cease, K.; Egeland, L.; Blinn, C. Aspen regeneration in riparian management zones in northern Minnesota: Effects of residual overstory and harvest method. North. J. Appl. For. 2003, 20, 79–84. [Google Scholar]
- Man, R.; Kayahara, G.J.; Rice, J.A.; MacDonald, G.B. Eleven-year responses of a boreal mixedwood stand to partial harvesting: Light, vegetation, and regeneration dynamics. For. Ecol. Manag. 2008, 255, 697–706. [Google Scholar] [CrossRef]
- Bradshaw, F.J. Quantifying edge effect and patch size for multiple-use silviculture—A discussion paper. For. Ecol. Manag. 1992, 48, 249–264. [Google Scholar] [CrossRef]
- Keenan, R.J.; Kimmins, J.P. The ecological effects of clear-cutting. Environ. Rev. 1993, 1, 121–144. [Google Scholar] [CrossRef]
- Baker, S.C.; Spies, T.A.; Wardlaw, T.J.; Balmer, J.; Franklin, J.F.; Jordan, G.J. The harvested side of edges: Effect of retained forests on the re-establishment of biodiversity in adjacent harvested areas. For. Ecol. Manag. 2013, 302, 107–121. [Google Scholar] [CrossRef]
- Baker, T.P.; Jordan, G.J.; Baker, S.C. Microclimatic edge effects in a recently harvested forest: Do remnant forest patches create the same impact as large forest areas? For. Ecol. Manag. 2016, 365, 128–136. [Google Scholar] [CrossRef]
- Harper, K.A.; Drapeau, P.; Lesiur, D.; Bergeron, Y. Negligible structural development and edge influence on the understory at 16–17-yr-old clear-cut edges in black spruce forest. Appl. Veg. Sci. 2016, 19, 462–473. [Google Scholar] [CrossRef]
- Scott, R.E.; Mitchell, S.J. Empirical modelling of windthrow risk in partially harvested stands using tree, neighbourhood, and stand attributes. For. Ecol. Manag. 2005, 218, 193–209. [Google Scholar] [CrossRef]
- Bladon, K.D.; Lieffers, V.J.; Silins, U.; Landhausser, S.M.; Blenis, P.V. Elevated mortality of residual trees following structural retention harvesting in boreal mixedwoods. For. Chron. 2008, 84, 70–75. [Google Scholar] [CrossRef]
- Wood, M.J.; Scott, R.; Voker, P.W.; Mannes, D.J. Windthrow in Tasmania, Australia: Monitoring, prediction and management. Forestry 2008, 81, 415–427. [Google Scholar] [CrossRef]
- LePage, P.T.; Canham, C.D.; Coates, K.D.; Bartemucci, P. Seed abundance versus substrate limitation of seedling recruitment in northern temperate forests of British Columbia. Can. J. For. Res. 2000, 30, 415–427. [Google Scholar] [CrossRef]
- Baker, S.C.; Strengbom, J.; Wardlaw, T.J.; Kern, C.; Edgar, G.J.; Thomson, R.J.; Bigley, R.E.; Franklin, J.F.; Gandhi, K.J.K.; Gustafsson, L.; et al. A cross-continental study of plant and beetle responses to retention of forest patches during timber harvest. Ecol. Appl. 2016, 26, 2493–2504. [Google Scholar] [CrossRef] [PubMed]
- Palik, B.J.; Kastendick, D. Response of seasonal pond plant communities to upland forest harvest in northern Minnesota forests, USA. For. Ecol. Manag. 2010, 260, 628–637. [Google Scholar] [CrossRef]
- Perala, D.A.; Alban, D. Allometric Biomass Estimators for Aspen-Dominated Ecosystems in the Upper Great Lakes; Research Paper NC-314; USDA Forest Service, North Central Experiment Station: St. Paul, MN, USA, 1994.
- Kruskal, J.B. Multidimensional scaling by optimizing goodness of fit to a nonmetric hypothesis. Psychometrika 1964, 29, 1–27. [Google Scholar] [CrossRef]
- McCune, B.; Grace, J.B. Analysis of Ecological Communities; MjM Software Design: Gleneden Beach, OR, USA, 2002; p. 300. [Google Scholar]
- Oksanen, J.; Blanchet, G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.4-1. 2016. Available online: Https://CRAN-R-project.org/package=vegan (accessed on 09 July 2016).
- Harper, K.A.; MacDonald, E.; Burton, P.J.; Chen, J.; Brosofske, K.D.; Saunders, S.C.; Euskirchen, E.S.; Roberts, D.; Jaiteh, M.S.; Esseen, P. Edge influence on forest structure and composition in fragmented landscapes. Conserv. Biol. 2005, 19, 768–782. [Google Scholar] [CrossRef]
- Harper, K.A.; Macdonald, S.E. Quantifying distance of edge influence: A comparison of methods and a new randomization method. Ecosphere 2011, 2, 1–17. [Google Scholar] [CrossRef]
- Dodonov, P.; Harper, K.A.; Silva-Matos, D.M. The role of edge contrast and forest structure in edge influence: Vegetation and microclimate at edges in the Brazilian cerrado. Plant Ecol. 2013, 214, 1345–1399. [Google Scholar] [CrossRef]
- Dupuch, A.; Fortin, D. The extent of edge effects increases during post-harvesting forest succession. Biol. Conserv. 2013, 162, 9–16. [Google Scholar] [CrossRef]
- Means, J.E.; Helm, M.E. Height Growth and Site Index Curves for Douglas-Fir on Dry Sites in the Willamette National Forest; PNW-341 Research Paper; USDA Forest Service, Pacific Northwest Forest and Range Experiment Station: Portland, OR, USA, 1995.
- Scott, R.E.; Neyland, M.G.; McElwee, D.J. Early regeneration results following aggregated retention harvesting of wet eucalypt forests in Tasmania, Australia. For. Ecol. Manag. 2013, 302, 254–263. [Google Scholar] [CrossRef]



| Structural Component | Size Class and Guild | Axis 1 (60.4%) | Axis 2 (16.3%) | Axis 3 (13.2%) | |||
|---|---|---|---|---|---|---|---|
| τ | p-Value | τ | p-Value | τ | p-Value | ||
| Mean diameter | Large sapling (DBH) * | 0.122 | 0.104 | 0.172 | 0.022 | 0.203 | 0.007 |
| Small sapling (D15) | −0.02 | 0.818 | −0.44 | <0.0001 | −0.21 | 0.006 | |
| Density | Medium shrubs | 0.507 | <0.0001 | 0.124 | 0.108 | −0.25 | 0.001 |
| Large shrubs | 0.436 | <0.0001 | 0.225 | 0.01 | 0.298 | 0.001 | |
| Small saplings * | 0.012 | 0.881 | −0.58 | <0.0001 | −0.23 | 0.003 | |
| Large saplings * | 0.436 | <0.0001 | 0.225 | 0.01 | 0.298 | 0.001 | |
| Biomass | Medium shrubs * | 0.645 | <0.0001 | 0.116 | 0.126 | −0.2 | 0.01 |
| Large shrubs | 0.457 | <0.0001 | 0.236 | 0.007 | 0.302 | 0.001 | |
| Small saplings | 0.02 | 0.794 | −0.45 | <0.0001 | 0.243 | 0.001 | |
| Large saplings | 0.166 | 0.027 | −0.31 | <0.0001 | 0.253 | 0.001 | |
| Woody understory * (>1 m height, <10 cm DBH) | 0.324 | <0.0001 | −0.31 | <0.0001 | 0.173 | 0.021 | |
| DBH diversity (H’) | All saplings * | 0.052 | 0.488 | −0.27 | 0.001 | 0.368 | <0.0001 |
| Cover (%) | Small shrubs * | 0.054 | 0.471 | 0.35 | <0.0001 | −0.34 | <0.0001 |
| Seedlings * | −0.02 | 0.752 | −0.28 | 0 | −0.03 | 0.668 | |
| POTR mean diameter | Large saplings (DBH) | 0.294 | 0 | 0.037 | 0.641 | 0.159 | 0.044 |
| POTR biomass (Mg/ha) | Small saplings | 0.055 | 0.486 | −0.34 | <0.0001 | 0.157 | 0.053 |
| Large saplings | 0.074 | 0.346 | −0.27 | 0.001 | 0.265 | 0.001 | |
| POTR density (stems/ha) | Small saplings | 0.095 | 0.254 | −0.34 | <0.0001 | −0.15 | 0.076 |
| Large saplings | 0.009 | 0.913 | −0.33 | <0.0001 | 0.159 | 0.044 | |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Curzon, M.T.; Baker, S.C.; Kern, C.C.; Palik, B.J.; D’Amato, A.W. Influence of Mature Overstory Trees on Adjacent 12-Year Regeneration and the Woody Understory: Aggregated Retention versus Intact Forest. Forests 2017, 8, 31. https://doi.org/10.3390/f8020031
Curzon MT, Baker SC, Kern CC, Palik BJ, D’Amato AW. Influence of Mature Overstory Trees on Adjacent 12-Year Regeneration and the Woody Understory: Aggregated Retention versus Intact Forest. Forests. 2017; 8(2):31. https://doi.org/10.3390/f8020031
Chicago/Turabian StyleCurzon, Miranda T., Susan C. Baker, Christel C. Kern, Brian J. Palik, and Anthony W. D’Amato. 2017. "Influence of Mature Overstory Trees on Adjacent 12-Year Regeneration and the Woody Understory: Aggregated Retention versus Intact Forest" Forests 8, no. 2: 31. https://doi.org/10.3390/f8020031
APA StyleCurzon, M. T., Baker, S. C., Kern, C. C., Palik, B. J., & D’Amato, A. W. (2017). Influence of Mature Overstory Trees on Adjacent 12-Year Regeneration and the Woody Understory: Aggregated Retention versus Intact Forest. Forests, 8(2), 31. https://doi.org/10.3390/f8020031