
C Stocks in Forest Floor and Mineral Soil of Two Mediterranean Beech Forests

 ,
, 
 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Litter Fall Collection
2.3. Forest Floor Litter and Mineral Soil Sampling
2.4. Litter and Soil Analyses
2.5. Determination of Active Fungal Mycelium, Microbial C and Potential Mineralization
2.6. Statistics
3. Results
3.1. Site Features
3.2. Stand Features
3.3. Nutrients
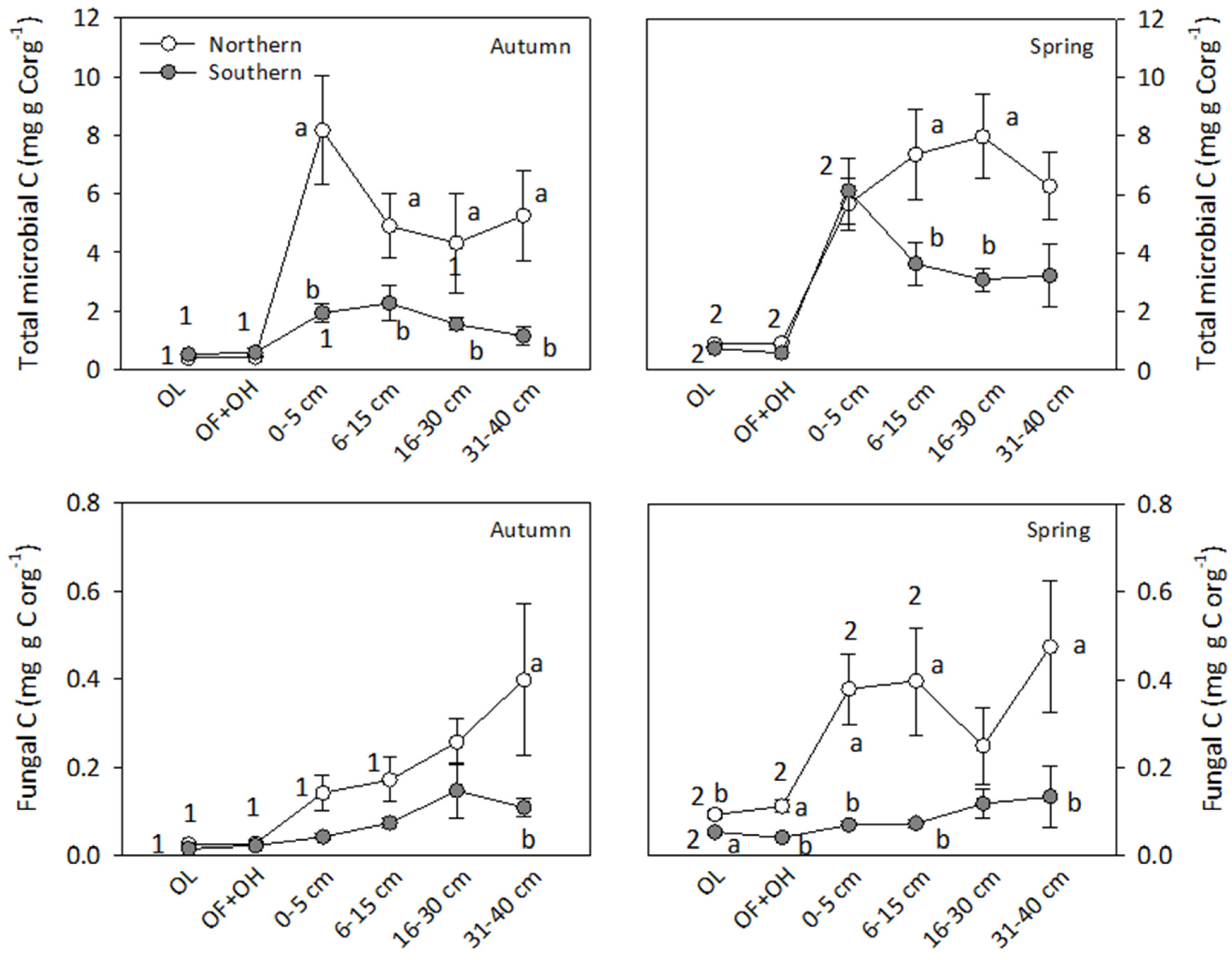
3.4. Stocks of Organic C and N, Water Soluble C and N, Microbial and Fungal C
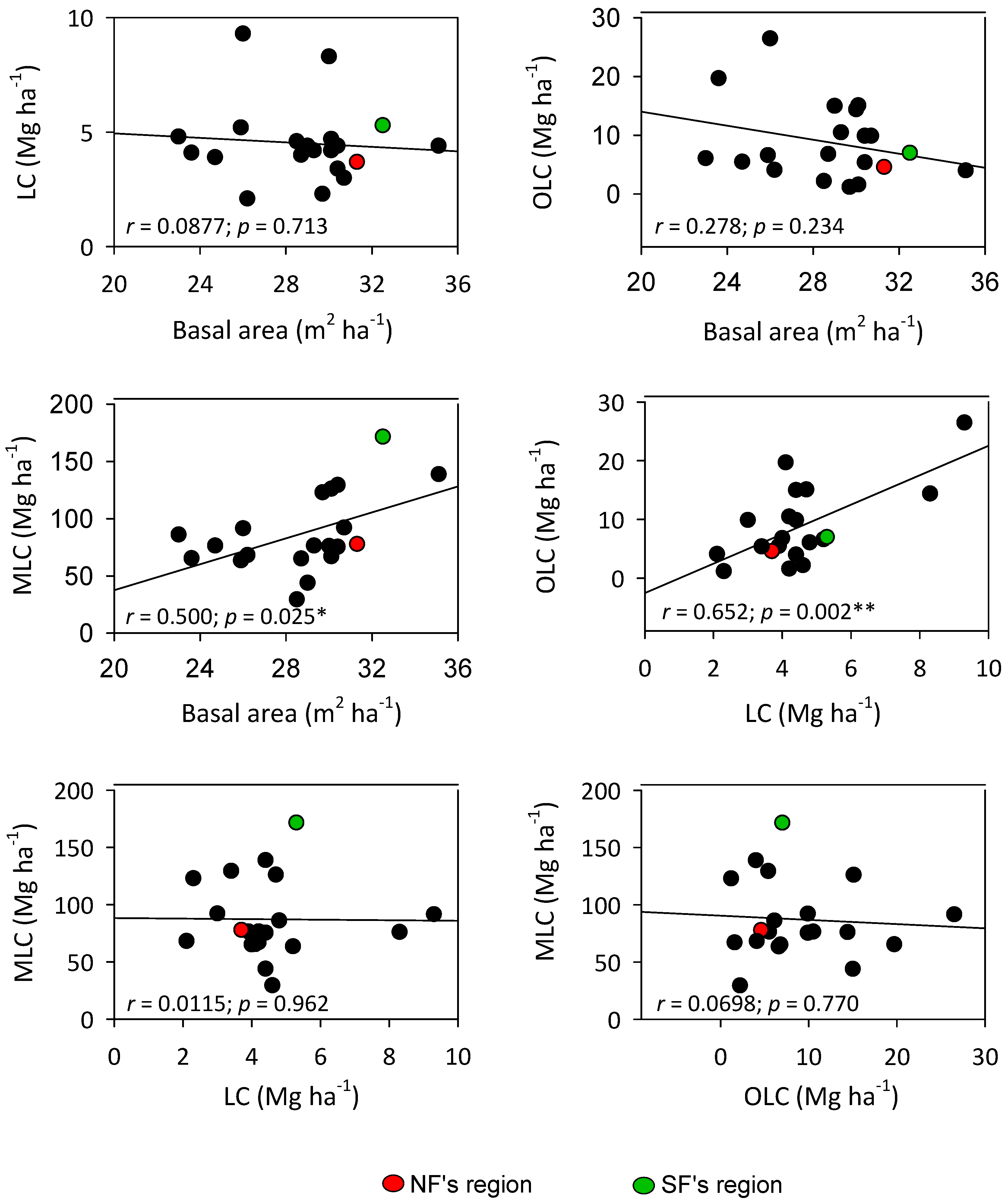
3.5. Distribution of C Stocks and Relationships with Basal Area in Italian Beech Forests (INFI Data)
4. Discussion
4.1. Basal Area, Litterfall and C and N Stocks
4.2. Nutrients
4.3. Soluble and Microbial C Fractions
4.4. Comparison of C Stocks in NF and SF with Regional Estimates of C Stocks from National Forest Inventory
5. Conclusions
- (1)
- the shorter growing season and the higher N concentration in litterfall, due to N deposition, that limit organic matter decay in the northern forest;
- (2)
- the tendency of the southern forest to produce a humus more rich in Mn, which enhances organic-matter decay.
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Jobbágy, E.G.; Jackson, R.B. The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecol. Appl. 2000, 10, 423–436. [Google Scholar] [CrossRef]
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A large and persistent carbon sink in the world’s forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef] [PubMed]
- Vesterdal, L.; Schmidt, I.K.; Callesen, I.; Ola Nilsson, L.; Gundersen, P. Carbon and nitrogen in forest floor and mineral soil under six common European tree species. For. Ecol. Manag. 2008, 255, 35–48. [Google Scholar] [CrossRef]
- Gasparini, P.; Di Cosmo, L. Forest carbon in Italian forests: Stocks, inherent variability and predictability using NFI data. For. Ecol. Manag. 2015, 337, 186–195. [Google Scholar] [CrossRef]
- Hobbie, S.E.; Reich, P.B.; Oleksyn, J.; Ogdahl, M.; Zytkowiak, R.; Hale, C.; Karolewski, P. Tree species effects on decomposition and forest floor dynamics in a common garden. Ecology 2006, 87, 2288–2397. [Google Scholar] [CrossRef]
- Davidson, E.A.; Janssens, I.A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature 2006, 440, 165–173. [Google Scholar] [CrossRef] [PubMed]
- FRA. Global Forest Resources Assessment Country Report Italy; FRA 2010/101 Forestry Department, Food and Agriculture Organization of the United Nations: Rome, Italy, 2010. [Google Scholar]
- Berg, B.; McClaugherty, C. Plant Litter Decomposition, Humus formation, Carbon Sequestration, 3rd ed.; Springer-Verlag: Berlin, Germany, 2014. [Google Scholar]
- Berg, B.; Johansson, M.B.; Ekbohm, G.; McClaugherty, C.; Rutigliano, F.A.; Virzo De Santo, A. Maximum decomposition limits of forest litter types: A synthesis. Can. J. Bot. 1996, 74, 659–672. [Google Scholar] [CrossRef]
- Berg, B.; Davey, M.P.; De Marco, A.; Emmett, B.; Faituri, M.; Hobbie, S.E.; Johansson, M.-B.; Liu, C.; McClaugherty, C.; Norell, L.; et al. Factors influencing limit values for pine needle litter decomposition: A synthesis for boreal and temperate pine forest system. Biogeochemistry 2010, 100, 57–73. [Google Scholar] [CrossRef]
- Hofrichter, M. Review: Lignin conversion by manganese peroxidase (MnP). Enzyme Microb. Technol. 2002, 30, 454–466. [Google Scholar] [CrossRef]
- Trum, F.; Titeux, H.; Ponette, Q.; Berg, B. Influence of manganese on decomposition of common beech (Fagus sylvatica L.) leaf litter during field incubation. Biogeochemistry 2015, 125, 349–358. [Google Scholar] [CrossRef]
- Mooshammer, M.; Wanek, W.; Schnecker, R.G.; Wild, B.; Leitner, S.; Hofhans, F.; Blo, A.; Hammerle Frank, A.H.; Fuchslueger, L.; Keiblinger, K.M.; et al. Stoichiometric controls of nitrogen and phosphorus cycling in decomposing beech leaf litter. Ecology 2012, 93, 770–782. [Google Scholar] [CrossRef] [PubMed]
- Berg, B.; Meentemeyer, V. Litter quality in a north European transect versus carbon storage potential. Plant Soil 2002, 242, 83–92. [Google Scholar] [CrossRef]
- Liu, C.J.; Berg, B.; Kutsch, W.; Westman, C.J.; Ilvesniemi, H.; Shen, X.; Shen, G.; Chen, X. Leaf litter nitrogen concentration as related to climatic factors in Eurasian forests. Glob. Ecol. Biogeogr. 2006, 15, 438–444. [Google Scholar] [CrossRef]
- Staaf, H. Plant nutrient changes in beech leaves during senescence as influenced by site characteristics. Acta Oecol. Plant 1982, 3, 161–170. [Google Scholar]
- Berg, B.; Erhagen, B.; Johansson, M.; Vesterdal, L.; Faituri, M.; Sanborn, P.; Nilsson, M. Manganese dynamics in decomposing needle and leaf litter—A synthesis. Can. J. For. Res. 2013, 43, 1127–1136. [Google Scholar] [CrossRef]
- Burrows, W.H.; Hoffmann, M.B.; Compton, J.F.; Back, P.V.; Tait, L.J. Allometric relationships and community biomass estimates for some dominant eucalypts in Central Queensland woodlands. Aust. J. Bot. 2000, 48, 707–714. [Google Scholar] [CrossRef]
- Guckland, A.; Jacob, M.; Flessa, H.; Thomas, F.M.; Leuschner, C. Acidity, nutrient stocks, and organic-matter content in soils of a temperate deciduous forest with different abundance of European beech (Fagus sylvatica L.). J. Plant Nutr. Soil Sci. 2009, 172, 500–511. [Google Scholar] [CrossRef]
- Keiblinger, K.M.; Schneider, T.; Roschitzki, B.; Schmid, E.; Eberl, L.; Hämmerle, L.I.; Leitner, S.; Richter, A.; Wanek, W.; Riedel, K.; et al. Effects of stoichiometry and temperature perturbations on beech leaf litter decomposition, enzyme activities and protein expression. Biogeosciences 2012, 9, 4537–4551. [Google Scholar] [CrossRef] [Green Version]
- Koranda, M.; Kaiser, C.; Fuchslueger, L.; Kitzler, B.; Sessitsch, A.; Zechmeister-Boltenstern, S.; Richter, A. Seasonal variation in functional properties of microbial communities in beech forest soil. Soil Biol. Biochem. 2013, 60, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Malý, S.; Fiala, P.; Reininger, D.; Obdržálková, E. The relationships among microbial parameters and the rate of organic matter mineralization in forest soils, as influenced by forest type. Pedobiologia 2014, 57, 235–244. [Google Scholar] [CrossRef]
- Buzek, F.; Paces, T.; Jackova, I. Production of dissolved organic carbon in forest soils along the north–south European transect. Appl. Geochem. 2009, 24, 1686–1701. [Google Scholar] [CrossRef]
- IPCC. Good Practice Guidance for Land Use, Land Use Change and Forestry; Institute for Global Environmental Strategies: Hayama, Japan, 2003; Available online: http://www.ipcc-nggip.iges.or.jp/public/gpglulucf/gpglulucf.html (accessed on 25 April 2016).
- Pignatti, S. I piani di vegetazione in Italia. G. Bot. Ital. 1979, 113, 411–428. [Google Scholar] [CrossRef]
- Gasparini, P.; Di Cosmo, L.; Pompei, E. IL contenuto di carbonio delle foreste italiane. In Inventario Nazionale delle Foreste e dei Serbatoi Forestali di Carbonio INFC 2005; CRA, Ed.; Metodi e risultati dell’indagine integrative; Consiglio per la Ricerca e la sperimentazione in Agricoltura: Trento, Italy, 2013. [Google Scholar]
- Fontaine, S.; Mariotti, A.; Abbadie, L. The priming effect of organic matter: A question of microbial competition? Soil Biol. Biochem. 2003, 35, 837–843. [Google Scholar] [CrossRef]
- Berg, B.; McClaugherty, C.; De Virzo Santo, A. Practicalities of estimating carbon sequestration—Boreal coniferous forests. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2008, 3, 84. [Google Scholar] [CrossRef]
- Sykes, M.T.; Prentice, I.C.; Cramer, W. A bioclimatic model for the potential distributions of north European tree species under present and future climates. J. Biogeogr. 1996, 23, 203–233. [Google Scholar]
- Berg, B.; Laskowski, R. Litter Decomposition: A Guide to Carbon and Nutrient Turnover; Elsevier: San Diego, CA, USA, 2006. [Google Scholar]
- Klironomos, J.N.; Rillig, M.C.; Allen, M.F. Designing belowground field experiments with the help of semi-variance and power analyses. Appl. Soil Ecol. 1999, 12, 227–238. [Google Scholar] [CrossRef]
- Ministero delle Politiche Agricole e Forestali—Osservatorio Nazionale Pedologico e per la Qualità del Suolo. Metodi Ufficiali di Analisi Chimica del Suolo; Violante, P., International Union of Soil Science, Società Italiana della Scienza del Suolo, Eds.; Franco Angeli Milano: Milan, Italy, 2000. (In Italian) [Google Scholar]
- García, C.; Hernández, T.; Costa, F. Study on water extract of sewage sludge composts. Soil Sci. Plant Nutr. 1991, 37, 399–408. [Google Scholar] [CrossRef]
- Bussotti, F.; Prancrazi, M.; Matteucci, G.; Gerosa, G. Leaf morphology and chemistry in Fagus sylvatica (beech) trees as affected by site factors and ozone: Results from CONECOFOR permanent monitoring plots in Italy. Tree Physiol. 2005, 25, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Olson, F.C.W. Quantitative estimates of filamentous algae. Trans. Am. Microsc. Soc. 1950, 69, 272–279. [Google Scholar] [CrossRef]
- Sundman, V.; Sivelä, S. A comment on the membrane filter technique for estimation of length of fungal hyphae in soil. Soil Biol. Biochem. 1978, 10, 399–401. [Google Scholar] [CrossRef]
- Berg, B.; Söderström, B. Fungal biomass and nitrogen in decomposing Scots pine needle litter. Soil Biol. Biochem. 1979, 11, 339–341. [Google Scholar] [CrossRef]
- Killham, K. Soil Ecology; Cambridge University Press: Cambridge, UK, 1994. [Google Scholar]
- Swift, M.J.; Heal, O.W.; Anderson, J.M. Decomposition in Terrestrial Ecosystem; Blackwell Scientific Publications: Oxford, UK, 1979. [Google Scholar]
- Anderson, T.H.; Domsch, K.H. A physiological method for the quantitative measurements of microbial biomass in soil. Soil Biol. Biochem. 1978, 10, 215–221. [Google Scholar] [CrossRef]
- Sparling, G.P. The soil biomass. In Soil Organic Matter and Biological Activity; Vaughan, D., Malcolm, R.E., Eds.; Nijhoff/Junk: Dordrecht, The Netherlands, 1995. [Google Scholar]
- Degens, B.P.; Schipper, L.A.; Sparling, G.P.; Vojvodic-Vukovic, M. Decreases in organic C reserves in soils can reduce the catabolic diversity of soil microbial communities. Soil Biol. Biochem. 2000, 32, 189–196. [Google Scholar] [CrossRef]
- Froment, A. Soil respiration in a mixed oak forest. Oikos 1972, 23, 273–277. [Google Scholar] [CrossRef]
- Balderas Torres, A.; Jon, C.; Lovett, J.C. Using basal area to estimate aboveground carbon stocks in forests: La Primavera Biosphere’s Reserve, Mexico. Forestry 2012, 86, 267–281. [Google Scholar] [CrossRef]
- Fichtner, A.; Sturm, K.; Rickert, C.; Hȁrdtle, W.; Schrautzer, J. Competition response of European beech Fagus sylvatica L. varies with tree size and abiotic stress: Minimizing anthropogenic disturbances in forests. J. Appl. Ecol. 2012, 49, 1306–1315. [Google Scholar] [CrossRef]
- Leith, H. Primary productivity in ecosystems: Comparative analysis of global patterns. In Unifying Concepts in Ecology; van Dobben, W.H., McConnel, L., Eds.; Junk: The Hague, The Netherlands, 1975. [Google Scholar]
- Vogt, K.A.; Grier, C.C.; Vogt, D.J. Production, turnover, and nutrient dynamics of above- and belowground detritus of world forests. Adv. Ecol. Res. 1986, 15, 303–377. [Google Scholar]
- Rasse, D.P.; Rumpel, C.; Dignac, M.-F. Is soil carbon mostly root carbon? Mechanisms for a specific stabilisation. Plant Soil 2005, 269, 341–356. [Google Scholar] [CrossRef]
- Olsen, J.S. Energy storage and the balance of producers and decomposers in ecological systems. Ecology 1963, 44, 322–331. [Google Scholar] [CrossRef]
- Andrén, O.; Paustian, K. Barley straw decomposition in the field: A comparison of models. Ecology 1987, 68, 1190–1200. [Google Scholar] [CrossRef]
- Trumbore, S.E. Comparison of carbon dynamics in tropical and temperate soils using radiocarbon measurements. Glob. Biogeochem. Cycles 1993, 7, 275–290. [Google Scholar] [CrossRef]
- Mosello, R.; Brizzio, M.C.; Kotzias, D.; Marchetto, A.; Rembges, D.; Tartari, G. The chemistry of atmospheric deposition in Italy in the framework of the National Programme for Forest Ecosystems Control (CONECOFOR). J. Limnol. 2002, 61, 77–92. [Google Scholar] [CrossRef]
- Coûteaux, M.M.; Bottner, P.; Berg, B. Litter decomposition, climate and litter quality. Trends Ecol. Evol. 1995, 10, 63–66. [Google Scholar] [CrossRef]
- Coûteaux, M.M.; McTiernan, K.; Berg, B.; Szuberla, D.; Dardenner, P. Chemical composition and carbon mineralisation potential of Scots pine needles at different stages of decomposition. Soil Biol. Biochem. 1998, 30, 583–595. [Google Scholar] [CrossRef]
- Tian, X.; Takeda, H.; Azuma, J. Dynamics of organic chemical components in leaf litters during a 3.5-year decomposition. Eur. J. Soil Biol. 2000, 36, 81–89. [Google Scholar] [CrossRef]
- Fioretto, A.; Di Nardo, C.; Papa, S.; Fuggi, A. Lignin and cellulose degradation and nitrogen dynamics during decomposition of three leaf litter species in a Mediterranean ecosystem. Soil Biol. Biochem. 2005, 37, 1083–1091. [Google Scholar] [CrossRef]
- Michel, K.; Matzner, E. Nitrogen content of forest floor Oa layers affects carbon pathways and nitrogen mineralization. Soil Biol. Biochem. 2002, 34, 1807–1813. [Google Scholar] [CrossRef]
- Berg, B.; Erhagen, B.; Johansson, M.-B.; Nilsson, M.; Stendahl, J.; Trum, F.; Vesterdal, L. Manganese in the litter fall-forest floor continuum of boreal and temperate pine and spruce forest ecosystems—A review. For. Ecol. Manag. 2015, 358, 248–260. [Google Scholar] [CrossRef]
- Innangi, M.; Schenk, M.K.; d’Alessandro, F.; Pinto, S.; Menta, C.; Papa, S.; Fioretto, A. Field and microcosms decomposition dynamics of European beech leaf litter: Influence of climate, plant material and soil with focus on N and Mn. Appl. Soil Ecol. 2015, 93, 88–97. [Google Scholar] [CrossRef]
- Joergensen, R.G.; Scholle, G.A.; Wolters, V. Dynamics of mineral components in the forest floor of an acidic beech (Fagus sylvatica L.) forest. Eur. J. Soil Biol. 2009, 45, 285–289. [Google Scholar] [CrossRef]
- Liverness, J.; Smith, T.D. The role of manganese in photosynthesis. Biochemistry 1982, 48, 1–44. [Google Scholar]
- Cleveland, C.C.; Townsend, A.R.; Schmidt, S.K. Phosphorus limitation of microbial processes in moist tropical forests: Evidence from short-term laboratory incubations and field studies. Ecosystems 2002, 5, 680–691. [Google Scholar] [CrossRef]
- Güsewell, S.; Gessner, M.O. N: P ratios influence litter decomposition and colonization by fungi and bacteria in microcosms. Funct. Ecol. 2009, 23, 211–219. [Google Scholar] [CrossRef]
- Smith, V.H. Effects of resource supply ratios on the structure and function of microbial communities. Antonie van Leeuwenhoek 2002, 81, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Rousk, J.; Brookes, P.C.; Bååth, E. Contrasting soil pH effects on fungal and bacterial growth suggest functional redundancy in carbon mineralization. Appl. Environ. Microb. 2009, 75, 1589–1596. [Google Scholar] [CrossRef] [PubMed]
- Kalbitz, K.; Solinger, S.; Park, J.H.; Michalzik, B.; Matzner, E. Controls on the dynamics of dissolved organic matter in soils: A review. Soil Sci. 2000, 165, 277–304. [Google Scholar] [CrossRef]
- Marschner, B.; Kalbitz, K. Controls of bioavailability and biodegradability of dissolved organic matter in soils. Geoderma 2003, 113, 211–235. [Google Scholar] [CrossRef]
- Nakanishi, T.; Atarashi-Andoha, M.; Koarashia, J.; Saito-Kokubub, Y.; Hiraic, K. Carbon isotopes of water-extractable organic carbon in a depth profile of forest soil imply a dynamic relationship with soil carbon. Eur. J. Soil Sci. 2012, 63, 495–500. [Google Scholar] [CrossRef]
- Schöning, I.; Kögel-Knabner, I. Chemical composition of young and old carbon pools throughout Cambisol and Luvisol profiles under forests. Soil Biol. Biochem. 2006, 38, 2411–2424. [Google Scholar] [CrossRef]
- Fontaine, S.; Barot, S.; Barré, P.; Bdioui, N.; Mary, B.; Rumpel, C. Stability of organic carbon in deep soil layers controlled by fresh carbon supply. Nature 2007, 450, 277–280. [Google Scholar] [CrossRef] [PubMed]
- Kögel-Knabner, I.; Guggenberger, G.; Kleber, M.; Kandeler, E.; Kalbitz, K.; Scheu, S.; Eusterhues, K.; Leinweber, P. Organo-mineral associations in temperate soils: Integrating biology, mineralogy, and organic matter chemistry. J. Plant Nutr. Soil Sci. 2008, 171, 61–82. [Google Scholar] [CrossRef]
- Matus, F.; Rumpel, C.; Neculman, R.; Panichini, M.; Mora, M.L. Soil carbon storage and stabilisation in andic soils: A review. Catena 2014, 120, 102–110. [Google Scholar] [CrossRef]
- Anderson, T.H. Microbial ecophysiological indicators to assess soil quality. Agric. Ecosyst. Environ. 2003, 98, 285–293. [Google Scholar] [CrossRef]
- Taylor, J.P.; Wilson, B.; Mills, M.S.; Burns, R.G. Comparison of microbial numbers and enzymatic activities in surface soils and subsoils using various techniques. Soil Biol. Biochem. 2002, 34, 387–401. [Google Scholar] [CrossRef]
- Sotomayor-Ramirez, D.; Espinoza, Y.; Acosta-Martinez, V. Land use effects on microbial biomass C, β-glucosidase and β-glucosaminidase activities, and availability, storage, and age of organic C in soil. Biol. Fertil. Soils 2009, 45, 487–497. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Northern Forest | Southern Forest | |
|---|---|---|
| Latitude/Longitude | 43°23′ N; 10°01′ E | 40°48′ N; 15°07′ E |
| Altitude (meters above sea level) | 1350 | 1150 |
| * Mean annual temperature (MAT) | 6.0 °C | 8.7 °C |
| * Mean temperature of the warmest month (TM) | 15.4 °C | 17.4 °C |
| * Mean temperature of the coldest month (Tm) | −2.7 °C | 0.3 °C |
| * Mean annual precipitation (MAP) | 2900 mm | 2300 mm |
| * Maximum monthly precipitation (PM) | 415 mm | 430 mm |
| * Minimum monthly precipitation (Pm) | 42 mm | 31 mm |
| Number of growing degree days | 178 | 238 |
| Soil parental substrate | Arenaceous, Lithological formation: Macigno | Calcareous covered by pyroclastic material |
| Soil 0–15 cm depth | ||
| Texture (%): | ||
| Sand | 34.2 | 29.7 |
| Silt | 40.4 | 47.4 |
| Clay | 25.4 | 22.8 |
| water content (% d.w.) | 72.4 ± 10.7 | 78.2 ± 2.6 |
| bulk density (g·cm−3) | 1.06 ± 0.06 | 0.99 ± 0.05 |
| organic matter content (%) | 20.2 ± 1.0 | 22.8 ± 1.2 |
| CEC (cmol·kg−1) | 29.4 ± 3.6 | 35.7 ± 2.8 |
| pH (H2O) | 3.9 ± 0.1 | 5.6 ± 0.1 |
| N (mg·g−1 d.w.) | 6.52 ± 1.75 | 9.38 ± 0.58 |
| P (mg·g−1 d.w.) | 0.04 ± 0.007 | 0.16 ± 0.01 |
| C/N | 14.7 ± 2.1 | 12.7 ± 0.7 |
| C/P | 3742 ± 348 | 1118 ± 247 |
| Northern Forest | Southern Forest | |
|---|---|---|
| Slope (degree) | 13.98° | 12.13° |
| Tree density (number·ha−1) | 1100 ± 55 | 800 ± 40 |
| Diameter at breast height (cm) | 25.4 ± 1.3 | 34.4 ± 2.2 |
| Basal area (m2·ha−1) | 55.7 | 74.3 |
| Litter input (Mg·ha−1) (n = 9) | 2.50 ± 0.18 | 4.31 ± 0.43 |
| Forest floor litter stock (Mg·ha−1) | 23.89 ± 2.12 | 21.18 ± 2.69 |
| Ratio litter stock to litter input | 9.6 | 4.9 |
| Nutrient content of newly shed leaves (mg·g−1) (n = 9) | ||
| C | 472.0 ± 1.45 | 466.7 ± 0.90 |
| N | 13.06 ± 0.05 | 10.17 ± 0.22 |
| C/N | 36 | 46 |
| P | 0.40 ± 0.06 | 0.62 ± 0.06 |
| Mn | 0.14 ± 0.005 | 0.06 ± 0.001 |
| Nutrient fluxes with litterfall (kg·ha−1) | ||
| N | 32.65 | 43.83 |
| P | 1.00 | 2.67 |
| Mn | 0.35 | 0.26 |
| Main features of forest floor litter (OL and OF + OH) * | ||
| pH (H2O) | 5.82 ± 0.10 | 6.18 ± 0.03 |
| water content (% d.w.) | 191.4 ± 13.0 | 224.7 ± 21.3 |
| OM (mg·g−1 d.w.) | 910.1 ± 15.4 | 728.1 ± 30.9 |
| N (mg·g−1 d.w.) | 21.21 ± 1.42 | 15.68 ± 1.27 |
| P (mg·g−1 d.w.) | 0.18 ± 0.01 | 0.38 ± 0.03 |
| Mn (mg g−1 d.w.) | 0.16 ± 0.02 | 0.25 ± 0.03 |
| C/N | 24 | 32 |
| C/P | 2925 | 1328 |
| C/Mn | 3690 | 2184 |
| C/Ca | 53 | 26 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Marco, A.; Fioretto, A.; Giordano, M.; Innangi, M.; Menta, C.; Papa, S.; Virzo De Santo, A. C Stocks in Forest Floor and Mineral Soil of Two Mediterranean Beech Forests. Forests 2016, 7, 181. https://doi.org/10.3390/f7080181
De Marco A, Fioretto A, Giordano M, Innangi M, Menta C, Papa S, Virzo De Santo A. C Stocks in Forest Floor and Mineral Soil of Two Mediterranean Beech Forests. Forests. 2016; 7(8):181. https://doi.org/10.3390/f7080181
Chicago/Turabian StyleDe Marco, Anna, Antonietta Fioretto, Maria Giordano, Michele Innangi, Cristina Menta, Stefania Papa, and Amalia Virzo De Santo. 2016. "C Stocks in Forest Floor and Mineral Soil of Two Mediterranean Beech Forests" Forests 7, no. 8: 181. https://doi.org/10.3390/f7080181
APA StyleDe Marco, A., Fioretto, A., Giordano, M., Innangi, M., Menta, C., Papa, S., & Virzo De Santo, A. (2016). C Stocks in Forest Floor and Mineral Soil of Two Mediterranean Beech Forests. Forests, 7(8), 181. https://doi.org/10.3390/f7080181