
From Litter to Humus in a Norwegian Spruce Forest: Long-Term Studies on the Decomposition of Needles and Cones

Abstract
:1. Introduction
2. Material and Methods
2.1. Study Site and Habitats
2.2. Experiments
3. Results
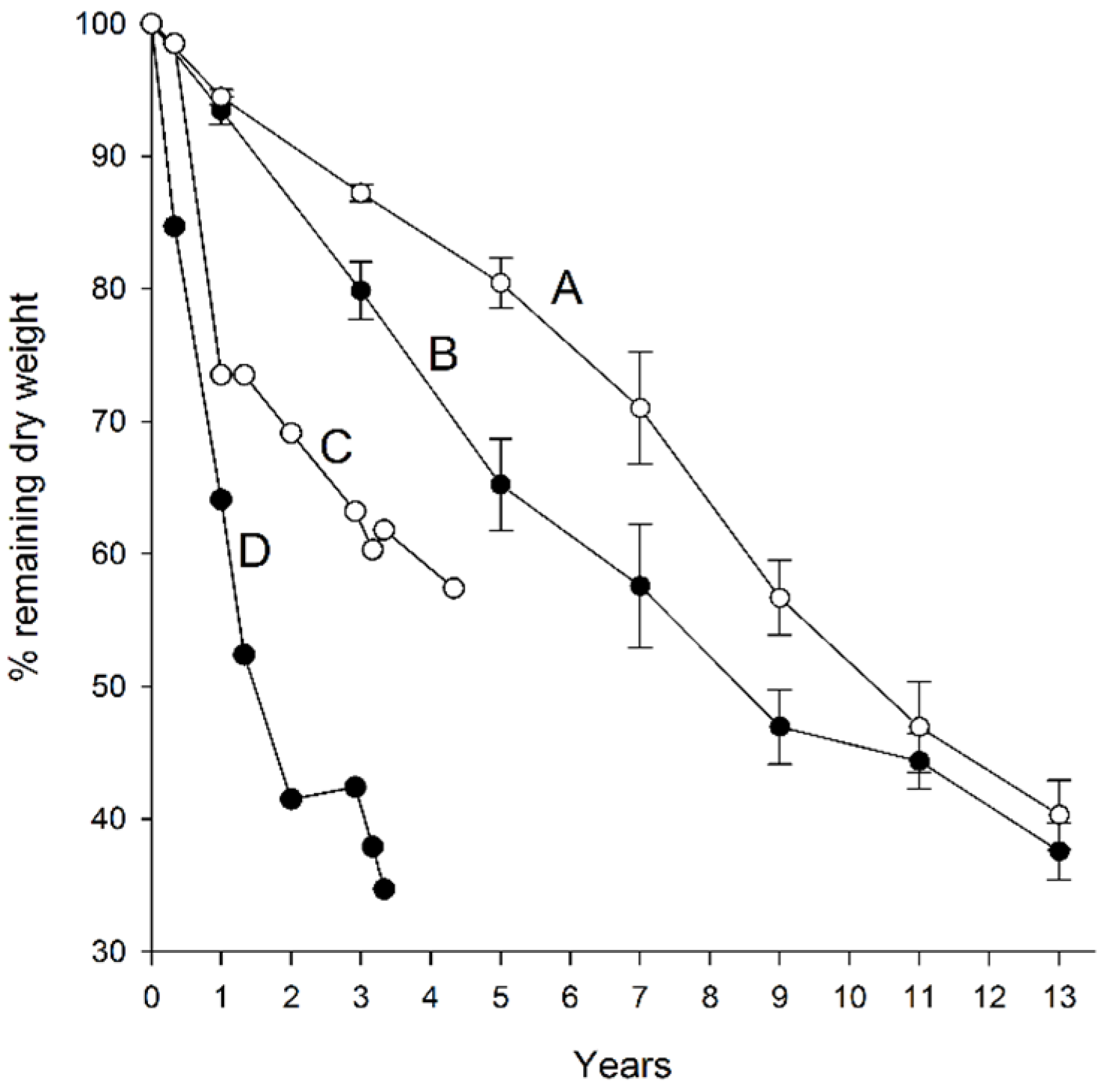
3.1. Dry Weight Loss
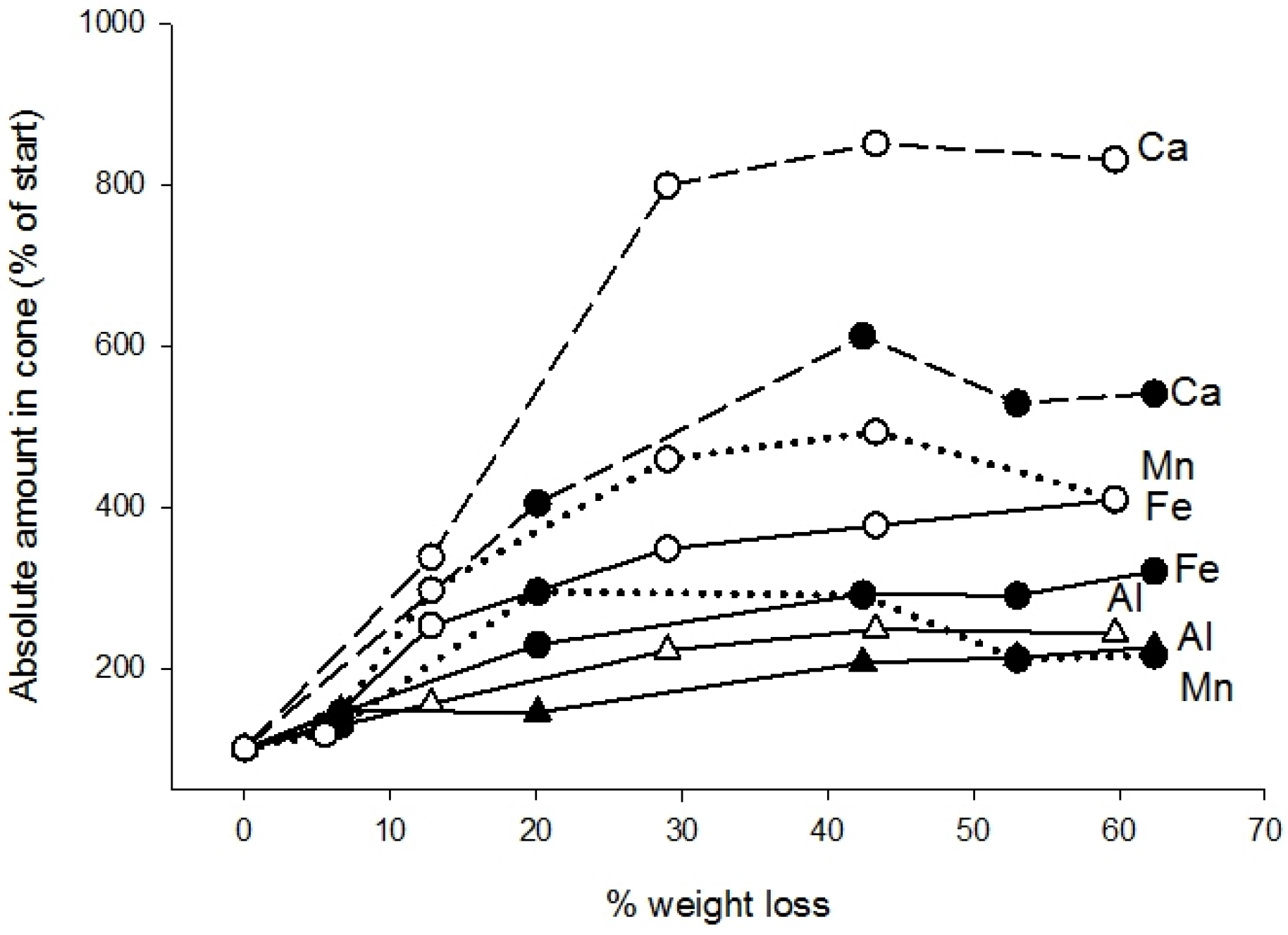
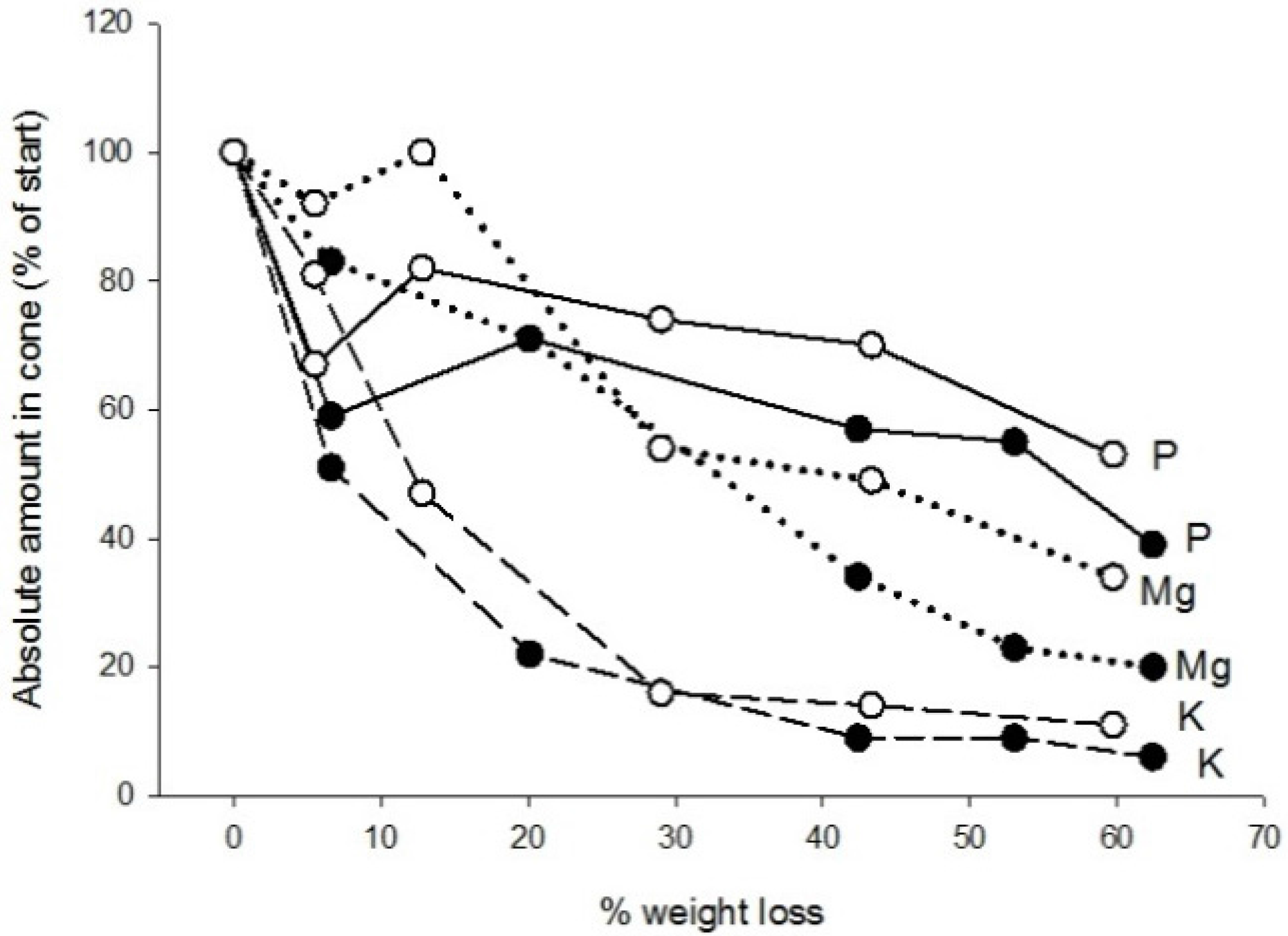
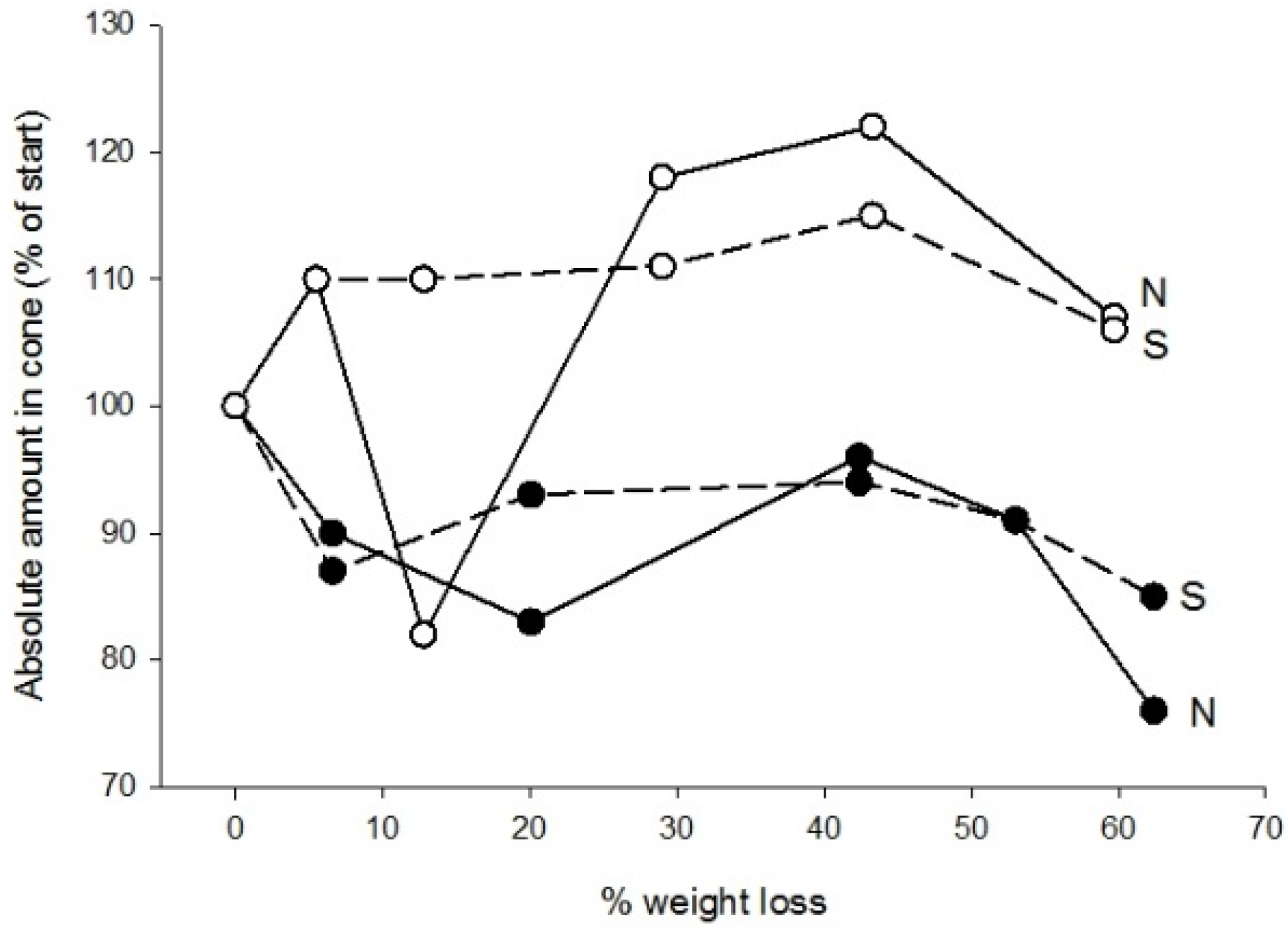
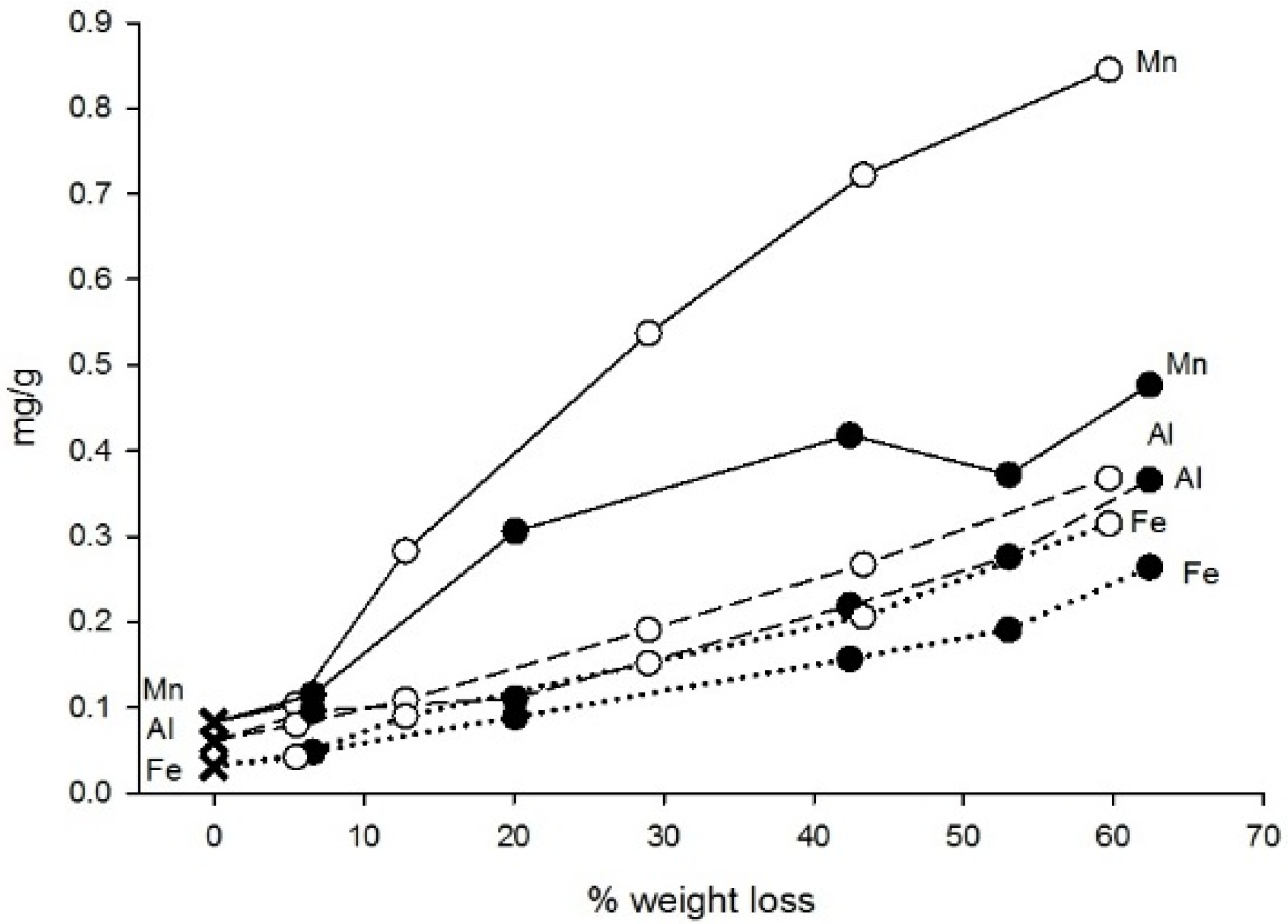
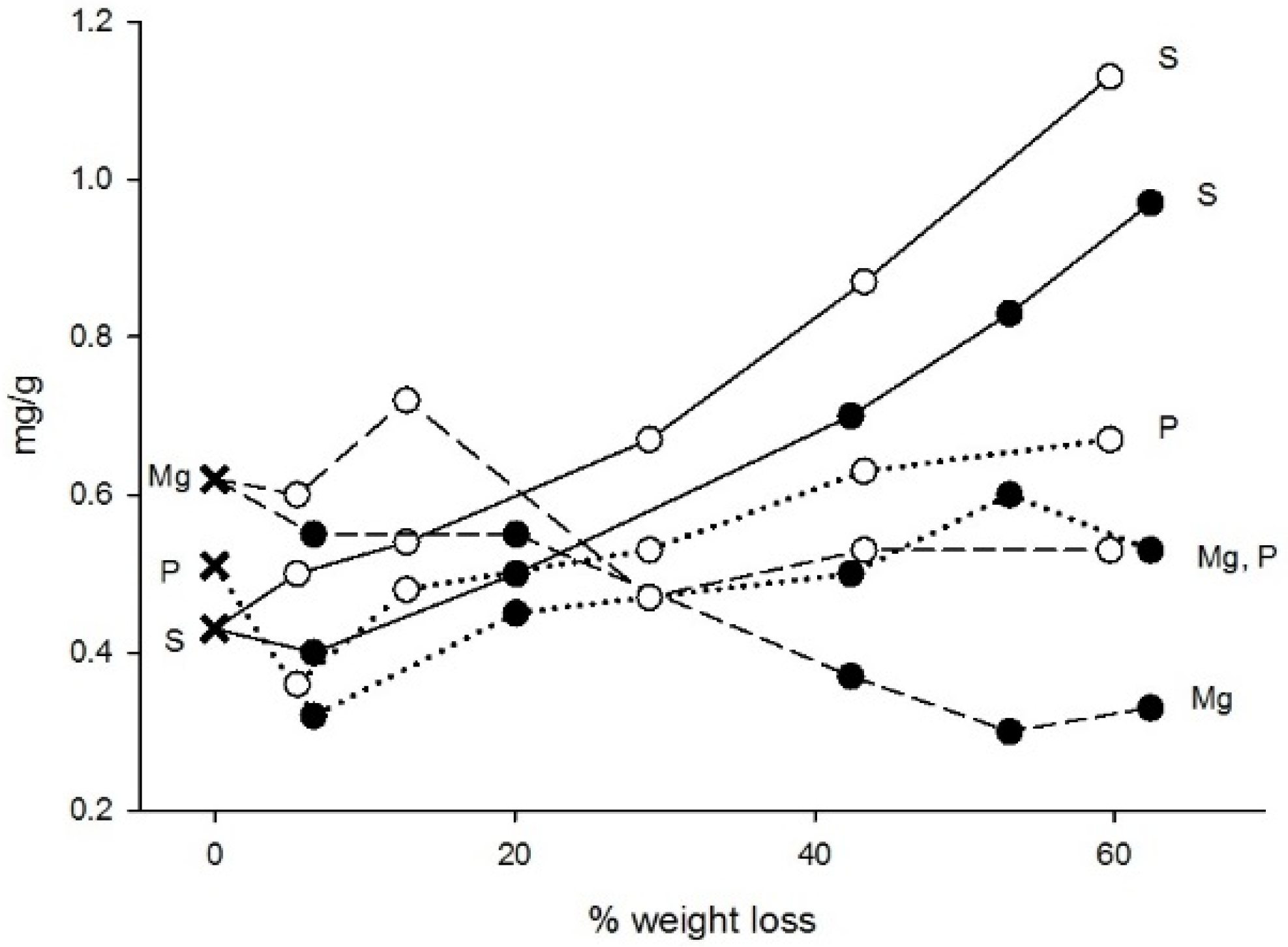
3.2. Chemistry
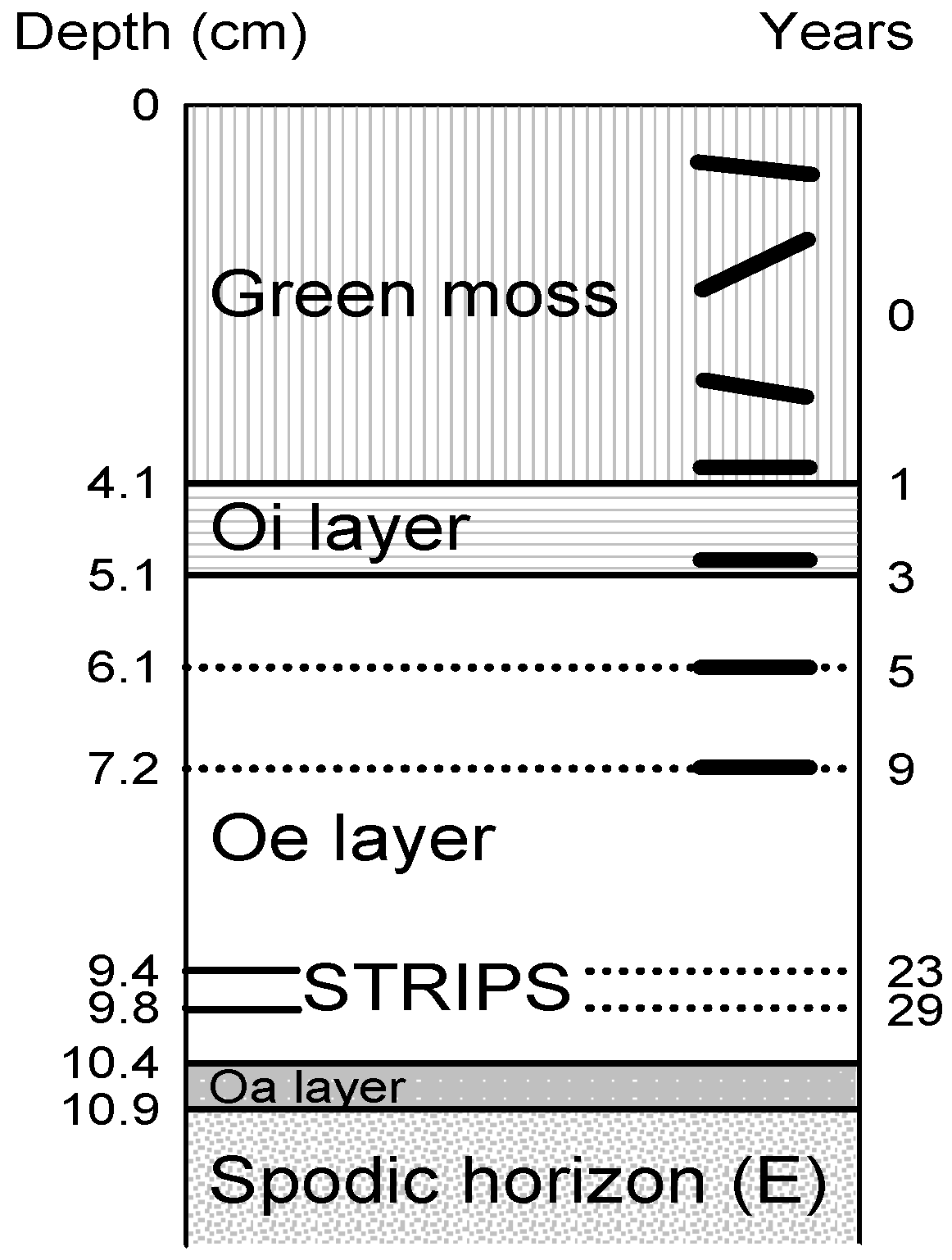
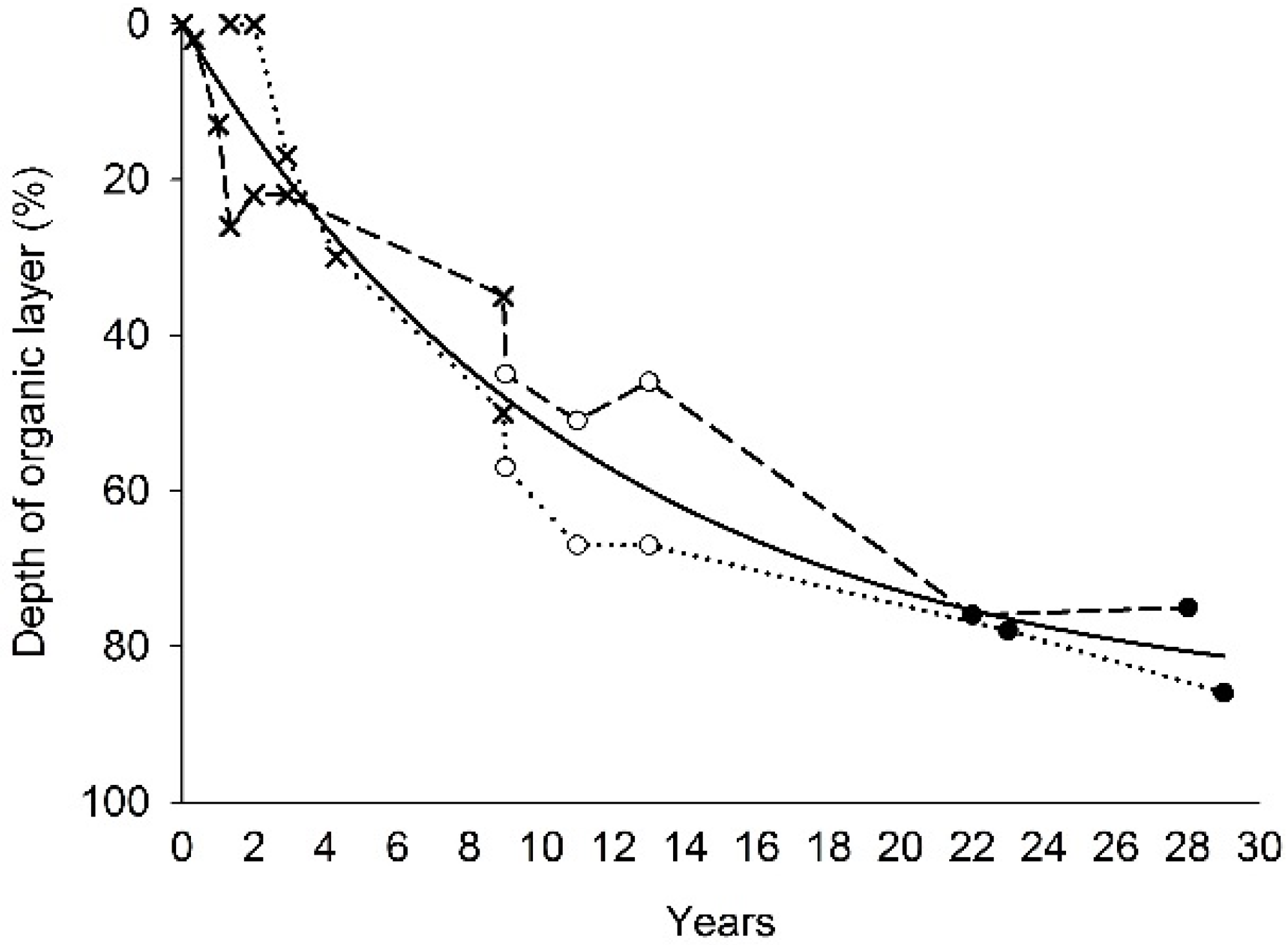
3.3. Vertical Transport
3.4. How Deep Can Fragments of Needles and Cones Be Recognized?
3.5. Soil Animals Associated with Needle Decomposition
4. Discussion
4.1. Vertical Transport in the Soil Profile
4.2. Chemistry and Decomposition Rate
4.3. Alternative Pathways from Litter to Humus
5. Conclusions
6. Suggestions for Future Research
Acknowledgments
Conflicts of Interest
References
- Berg, B.; McClaugherty, C. Plant Litter. Decomposition, Humus Formation, Carbon Sequestration; Springer: Berlin, Germany, 2003; p. 286. [Google Scholar]
- Ohlson, M.; Tryterud, E. Long-term spruce forest continuity—A challenge for a sustainable Scandinavian forestry. For. Ecol. Manag. 1999, 124, 27–34. [Google Scholar] [CrossRef]
- Wardle, D.A.; Zackrisson, O.; Hörnberg, G.; Gallet, C. The influence of island area on ecosystem properties. Science 1997, 277, 1296–1299. [Google Scholar] [CrossRef]
- Mork, E. Om strøfallet i våre skoger. En analyse av skogstrøets mengde, kjemiske innhold og de faktorer som innvirker på disse; Reports of The Norwegian Forest Research Institute: Aas, Norway, 1942; pp. 297–365. [Google Scholar]
- Bonnevie-Svendsen, C.; Gjems, O. Amount and chemical composition of the litter from larch, beech, Norway spruce and Scots pine stands and its effect on the soil; Reports of The Norwegian Forest Research Institute: Aas, Norway, 1957; pp. 111–174. [Google Scholar]
- Røsberg, I. Deposisjon av næringsstoff med strø og nedbør i granbestand på Nannestad, Akershus. In Symposium in Vegetation Ecology at Kongsvold 1990 and 1991; Bretten, S., Krovoll, A., Eds.; University of Trondheim; Vitenskapsmuseet: Trondheim, Norway; Rapport Botanisk Serie; 1991; Volume 2, pp. 136–155. [Google Scholar]
- Hagner, S. Cone crop fluctuations in Scots pine and Norway spruce. Stud. For. Suec. 1965, 33, 1–21. [Google Scholar]
- Johansson, M.-B. Decomposition of leaf and root litters from some coniferous and broad-leaved trees at sites located in different parts of Sweden. In Reports in Forest Ecology and Forest Soils; Swedish University of Agricultural Sciences: Uppsala, Sweden, 1984; Volume 42, p. 28. [Google Scholar]
- Johansson, M.-B. Chemical composition and decomposition pattern of leaf litters from forest trees in Sweden with special reference to methodological aspects and site properties. In Reports in Forest Ecology and Forest Soils; Doctoral Dissertation; Swedish University of Agricultural Sciences: Uppsala, Sweden, 1986; Volume 56. [Google Scholar]
- Johansson, M.-B. The influence of soil scarification on the turn-over rate of slash needles and nutrient release. Scand. J. For. Res. 1994, 9, 170–179. [Google Scholar] [CrossRef]
- Johansson, M.-B. The chemical composition of needle and leaf litter from Scots pine, Norway spruce and white birch in Scandinavian forests. Forestry 1995, 68, 49–62. [Google Scholar] [CrossRef]
- Johansson, M.-B.; Grälls, C. Quantity, decomposition and nutrient dynamics of litterfall in some forest stands at Hålebäck, southern Sweden. In Reports in Forest Ecology and Forest Soils; Swedish University of Agricultural Sciences: Uppsala, Sweden, 1989; Volume 62, p. 26. [Google Scholar]
- Berg, B. Nutrient release from litter and humus in coniferous forest soils—A mini review. Scand. J. For. Res. 1986, 1, 359–369. [Google Scholar] [CrossRef]
- Hågvar, S. Succession of microarthropods in decomposing spruce needles: A litter bag study over ten years. In Advances in Management and Conservation of Soil Fauna; Veeresh, G.K., Rajagopal, D., Viraktamath, C.A., Eds.; Oxford & IBH Publishing Co. Pvt. Ltd.: New Delhi, India, 1991; pp. 485–489. [Google Scholar]
- Hågvar, S. Mites (Acari) developing inside decomposing spruce needles: Biology and effect on decomposition rate. Pedobiologia 1998, 42, 358–377. [Google Scholar]
- Hågvar, S. Decomposing cones of Norway spruce (Picea abies (L.) H. Karst.): Dry weight loss, chemical changes, and vertical transport in a Norwegian raw humus soil profile. Scand. J. For. Res. 2015, 30, 643–652. [Google Scholar]
- Hågvar, S.; Edsberg, E. Vertical transport of decomposing spruce needles during nine years in a raw humus soil profile in southern Norway. Pedobiologia 2000, 44, 119–131. [Google Scholar] [CrossRef]
- Abrahamsen, G. Ecological study of Lumbricidae (Oligochaeta) in Norwegian coniferous forest soils. Pedobiologia 1972, 12, 267–281. [Google Scholar]
- Olson, J.S. Energy storage and the balance of producers and decomposers in ecological systems. Ecology 1963, 44, 322–331. [Google Scholar] [CrossRef]
- Berg, B. Research notes on decomposition. 2: Transport of pine needles through moss communities of the S-layer in a 120-year-old stand of Pinus silvestris (Ih VA); Swedish Coniferous Forest Project, Internal Report 63; Swedish Agriculture University: Uppsala, Sweden, 1977; pp. 15–19. [Google Scholar]
- Roland, C.; Schmidt, J.; Johnstone, J. Climate sensitivity of reproduction in a mast-seeding boreal conifer across its distributional range from lowland to treeline forests. Oecologia 2014, 174, 665–677. [Google Scholar] [CrossRef] [PubMed]
- Bécard, G.; Piché, Y. Fungal growth stimulation by CO2 and root exudates in vesicular-arbuscular mycorrhizal symbiosis. Appl. Environ. Microbiol. 1989, 55, 2320–2325. [Google Scholar] [PubMed]
- Bücking, H.; Abubaker, J.; Govindarajulu, M.; Tala, M.; Pfeffer, P.E.; Nagahashi, G.; Lammers, P.; Shachar-Hill, Y. Root exudates stimulate the uptake and metabolism of organic carbon in germinating spores of Glomus intraradices. New Phytol. 2008, 180, 684–695. [Google Scholar] [CrossRef] [PubMed]
- Seastedt, T.R. The role of microarthropods in decomposition and mineralization processes. Ann. Rev. Entomol. 1984, 29, 25–46. [Google Scholar] [CrossRef]
- Hågvar, S.; Kjøndal, B.R. Succession, diversity and feeding habits of microarthropods in decomposing birch leaves. Pedobiologia 1981, 22, 385–408. [Google Scholar]
- Hågvar, S.; Kjøndal, B.R. Decomposition of birch leaves: Dry weight loss, chemical changes, and effects of artificial acid rain. Pedobiologia 1981, 22, 232–245. [Google Scholar]
- Gourbière, F.; Lions, J.C.; Repin, R. Activité et développement d’Adoristes ovatus (C.L. Koch, 1839) (Acarien, Oribate) dans les aiguilles d’Abies alba Mill. Relations avec la décomposition et les microflores fongiques. Rev. Ecol. Biol. Sol. 1985, 22, 57–73. [Google Scholar]
- Lions, J.C.; Gourbière, F. Populations adultes et immatures d’Adoristes ovatus (Acarien, Oribate) dans les aiguilles de la litière d’Abies alba. Rev. Ecol. Biol. Sol. 1988, 25, 343–352. [Google Scholar]
- Lions, J.C.; Gourbière, F. Populations d’Adoristes ovatus (Acarien, Oribate) vivant à l’extérieur des aiguilles dans la litière d’Abies alba. Rev. Ecol. Biol. Sol. 1989, 26, 213–223. [Google Scholar]
- Wallwork, J.A. Acari. In Soil Biology; Burges, A., Raw, F., Eds.; Academic Press: London, UK; New York, USA, 1967; pp. 363–396. [Google Scholar]
- Harding, D.J.L.; Easton, S.M. Development of two species of Phthiracarid mites in beech cupules. Acarology 1984, VI, 860–870. [Google Scholar]
- Webb, N.R. Observations on Steganacarus magnus. General biology and life cycle. Acarologia 1977, XIX, 686–696. [Google Scholar]
- Berg, B.; Lohm, U.; Lundkvist, H.; Wirén, A. Influence of soil animals on decomposition of scots pine needle litter. In Ecological Bulletins 32: Structure and Function of Northern Coniferous Forests: An Ecosystem Study; Persson, T., Ed.; Oikos Editorial Office: Lund, Sweden, 1980; pp. 401–409. [Google Scholar]
- Pande, Y.D.; Berthet, P. Studies on the food and feeding habits of soil Oribatei in a black pine plantation. Oecologia 1973, 12, 413–426. [Google Scholar] [CrossRef]
- Zachariae, G. Was leisten Collembolen für den Waldhumus? In Soil Organism; Doeksen, J., van der Drift, J., Eds.; North-Holland: Amsterdam, The Netherlands, 1963; pp. 109–124. [Google Scholar]
- Culliney, T.W. Role of arthropods in maintaining soil fertility. Agriculture 2013, 3, 629–659. [Google Scholar] [CrossRef]
- Fenton, N.J.; Bergeron, Y.; Paré, D. Decomposition rates of bryophytes in managed boreal forests: Influence of bryophyte species and forest harvesting. Plant Soil 2010, 336, 499–508. [Google Scholar] [CrossRef]
- Turetsky, M.R.; Mack, M.C.; Hollingsworth, T.N.; Harden, J.W. The role of mosses in ecosystem succession and function in Alaska’s boreal forest. Can. J. For. Res. 2010, 40, 1237–1264. [Google Scholar] [CrossRef]
- Staaf, H.; Berg, B. A Structural and Chemical Description of Litter and Humus in a Mature Scots Pine Stand; Swedish Coniferous Forest Project, Internal Report 65; Swedish Agriculture University: Uppsala, Sweden, 1977; pp. 1–31. [Google Scholar]
- Berg, B. Decomposition of moss litter in a mature Scots pine forest. Pedobiologia 1984, 26, 301–308. [Google Scholar]
- Turetsky, M.R. The role of bryophytes in carbon and nitrogen cycling. Bryologist 2003, 106, 395–409. [Google Scholar] [CrossRef]
- Treseder, K.K.; Holden, S.R. Fungal carbon sequestration. Science 2013, 339, 1528–1529. [Google Scholar] [CrossRef] [PubMed]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment No. | Litter Type | Method | Habitat | Purpose | Duration | Period | Reference |
|---|---|---|---|---|---|---|---|
| 1 | Spruce needles | Cylindrical litter bags | Clearcut area | Decomposition rate Microarthropod succession | 10 years | 1977–1987 | [14] |
| 2 | Spruce needles | Needles fastened to nylon strips | No vegetation. | Decomposition rate Effect of mites inside needles Vertical transport | 4 years Strips 28 years | 1986–1990 Strips to 2014 | [15,16] |
| Moss vegetation | |||||||
| Bilberry and moss | |||||||
| 3 | Spruce needles | Marked needles dropped on the ground | No vegetation. | Vertical transport | 9 years | 1986–1995 | [17] |
| Moss vegetation | |||||||
| Bilberry and moss | |||||||
| 4 | Spruce cones | Marked cones placed on the ground | No vegetation. | Vertical transport Decomposition rate Chemistry | 13 years | 1995–2008 | [16] |
| Moss vegetation |
| Curve in Figure 2 | Habitat | Litter Type | Rate Constant | Half Time (Years) |
|---|---|---|---|---|
| A | Needle mat | Cones | 0.0703 | 9.9 |
| B | Moss | Cones | 0.0777 | 8.9 |
| C | Needle mat | Needles | 0.0119 | 4.9 |
| D | Moss | Needles | 0.0274 | 2.1 |
| Element | Needles | Cones | ||
|---|---|---|---|---|
| Concentration at Start (mg·g−1) | Concentration at Start (mg·g−1) | Change in Concentration | Change in Total Amount | |
| N | 8.5 | 4.80 ± 0.81 | Increase | Rather stable |
| P | 1.05 | 0.51 ± 0.17 | Stable | Decrease |
| K | 1.73 | 3.03 ± 0.95 | Decrease | Decrease |
| Ca | 7.00 | 0.16 ± 0.05 | Increase | Increase to stable |
| Mg | 0.60 | 0.62 ± 0.13 | Decrease or stable | Decrease |
| S | 0.73 | 0.43 ± 0.10 | Increase | Rather stable |
| Mn | 1.98 | 0.083 ± 0.017 | Increase | Increase to stable |
| Fe | 0.116 | 0.031 ± 0.009 | Increase | Increase to stable |
| Cu | 0.0034 | 0.0037 ± 0.0008 | Increase | Decrease |
| Zn | 0.050 | 0.025 ± 0.007 | Increase | Varying |
| Al | 0.322 | 0.06 ± 0.01 | Increase | Increase to stable |
| Age of Needle (Years) | 0 | 0 | 1–3 | 3–4 | 5 Years or More | Several Decades |
|---|---|---|---|---|---|---|
| Soil layer | Surface | Green moss | Litter (Oi) | Upper Oe | Oe | Oa |
| Alternatives for a single needle | Forest floor: No vegetation or moss cover? | If moss, how deep into the moss did the falling needle penetrate? | Is the needle inhabited by juvenile mites? | Is the needle fed upon by adult mites? | Is the needle close to a fine root? | Results in Oa |
| Slower decomposition | No vegetation | Needle landed in upper moss | Yes | Yes | No | Resistable faecal pellets. Recognizable, three-dimentional fragments of needles. |
| Faster decomposition | Moss cover | Needle penetrated through moss at landing | No | No | Yes | No faecal pellets. No recognizable fragments of needles. |
© 2016 by the author; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hågvar, S. From Litter to Humus in a Norwegian Spruce Forest: Long-Term Studies on the Decomposition of Needles and Cones. Forests 2016, 7, 186. https://doi.org/10.3390/f7090186
Hågvar S. From Litter to Humus in a Norwegian Spruce Forest: Long-Term Studies on the Decomposition of Needles and Cones. Forests. 2016; 7(9):186. https://doi.org/10.3390/f7090186
Chicago/Turabian StyleHågvar, Sigmund. 2016. "From Litter to Humus in a Norwegian Spruce Forest: Long-Term Studies on the Decomposition of Needles and Cones" Forests 7, no. 9: 186. https://doi.org/10.3390/f7090186
APA StyleHågvar, S. (2016). From Litter to Humus in a Norwegian Spruce Forest: Long-Term Studies on the Decomposition of Needles and Cones. Forests, 7(9), 186. https://doi.org/10.3390/f7090186